
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Глава одиннадцатая. Геологическая история растений
Наш обзор развития биосферы в фанерозое логично начать с растений. Растения не только прямо или опосредствованно служат пищей животным; летопись ископаемых остатков показывает, что растения раньше животных покинули море, эту колыбель всех форм жизни. Следовательно, мы сможем легче проследить историю развития животных, если сначала опишем растительный покров континентов и развитие растительности до наших дней. Летопись ископаемых растений, особенно в своей ранней части, изобилует довольно большими пробелами. Это не удивительно, потому что условия для сохранности ископаемых остатков на суше хуже, чем на дне моря, а большинство ископаемых растений росло именно на суше. Такие пробелы затрудняют установление эволюционных связей между группами растений. Несмотря на это, если не вдаваться в детали, можно наметить общую историю, основанную на известных к настоящему времени растительных остатках.
Структура и физиология растений
Сосудистая система. Краткий обзор особенностей современных растений - основных черт их строения и физиологии - показывает результат развития растительного мира. Большинство растений, особенно высших, отличается характерными чертами строения; все они, в частности, имеют: 1) длинный, обычно вертикальный и ветвящийся стебель, в большинстве случаев жесткий; 2) корни, хорошо закрепленные в почве; 3) в верхней части - зеленую листву. Самый важный процесс - фотосинтез - происходит в листьях, безразлично, широкие это листья, иглы или еще какие-нибудь. Клетки листьев синтезируют хлорофилл ("растительную зелень"), зеленый пигмент, который поглощает солнечный свет и, используя его энергию, может создавать питательные вещества из углекислого газа атмосферы и воды, которую растение извлекает из почвы. Через поры листьев поступает углекислый газ, происходит реакция фотосинтеза (химическое восстановление кислорода) и в результате образуются сахар, вода и свободный кислород.
Вода, которая нужна для фотосинтеза, поступает снизу, из почвы, по системе корней, через стебель и ветви. Теперь мы понимаем, что корни, стебель и ветви созданы не просто для того, чтобы нести на себе листья. Это также и водоподводящая система каналов, по которой вода в достаточных количествах подается даже на вершину самого высокого дерева. Очень высокие хвойные деревья на тихоокеанском побережье Северной Америки могут подтягивать воду на высоту до 120 метров над землей.
В действительности вода, которая подтягивается вверх, к листьям, это не просто вода; это сок деревьев, водный раствор. В одеревеневшей ткани живого дерева находятся вертикальные цепочки мертвых клеток. У некоторых видов деревьев верхние и нижние стенки этих клеток растворены - как если бы в высоком здании пришлось уничтожить перекрытия в целом ряде комнат, расположенных одна над другой, чтобы соорудить шахту для лифта. Цепочки этих мертвых клеток заполнены соком. Они образуют трубочки, по которым жидкость, поступающая из земли, подтягивается от корней к живым листьям. Из листьев вода постоянно испаряется в атмосферу через поры. Эта потеря влаги снижает давление внутри листьев и тонких веток и заставляет сок, заполняющий систему трубок, подтягиваться через стенки клеток к листьям, чтобы возместить недостаток жидкости. Таким образом всасывающее усилие передается по всей системе трубок, включая корни, которые в свою очередь начинают более активно поглощать влагу из почвы. Вся эта система "сосудов" представляет собой своего рода насос, действующий весьма эффективно.
Эта система, рассматриваемая в более широком плане, представляет собой часть круговорота воды (рис. 4). В таком круговороте часть осадков просачивается в землю, поглощается корнями растений, поднимается к листьям и с их поверхности испаряется в атмосферу, из которой выпадает затем в виде дождя. Но часть воды остается в растениях. В процессе фотосинтеза она соединяется с углекислым газом, который проникает в растение из атмосферы через поры листьев, и образует питательные вещества и ткани растения.
Для образования тканей растения требуется большое количество воды и углекислого газа. Например, на 0,4 га кукурузного поля произрастает около 10 000 растений, которые дают около 3,5 м3 зерна (семян кукурузы). Для образования этого количества семян (мы не считаем стебли, корни, листья и стержни початков) растениям требуется около 9 т углекислого газа, из которого они выделяют и используют 2,5 т углерода, а освободившийся кислород возвращают в атмосферу. Для этого растения должны переработать 21 000 т воздуха, то есть более 2 т на каждое растение. В то же время растения должны извлечь из почвы и перекачать в листья 1 892 500 литров воды, что составляет около 190 литров на каждое растение.
Этот процесс происходит на каждом участке кукурузного поля ежегодно. То же самое происходит на площадях, засеянных пшеницей и другими зерновыми. Такие же процессы, хотя и происходящие более медленно, наблюдаются на площадях, занятых деревьями, дающими фрукты или орехи или использующимися как топливо и строительный материал. Представив себе огромные пространства лесов, садов, пастбищ и обрабатываемых полей, безразлично, используемых человеком или нет, мы до некоторой степени можем понять, какую огромную роль растения, входящие в состав биосферы, играют в круговороте воды, а также в процессах выветривания (физического и химического), образующих часть круговорота горных пород.
Система размножения. Изучением способа размножения живых организмов, животных или растений, никогда не следует пренебрегать, потому что успешное воспроизводство организма - это один из важнейших факторов его успешной эволюции, залог способности выживать в меняющихся (к лучшему или худшему) условиях. Поэтому, рассмотрев способы размножения современных растений, мы можем узнать что-то новое и о геологической истории.
Небольшая часть ныне существующих растений размножается вегетативно. При вегетативном размножении у растения развиваются отростки, луковицы или побеги, которые укореняются и становятся самостоятельными растениями, точно воспроизводящими своего родителя. (Мы употребляем это слово в единственном числе, так как в этом случае родитель только один.) При таком размножении сочетания различных генов не происходит; потомок не приобретает никаких новых свойств, которые могли бы послужить ему для приспособления к новым условиям среды.
Половое размножение, напротив, допускает сочетание различных свойств, унаследованных от различных особей, и если условия среды благоприятствуют развитию возникающих признаков, то эволюция ускоряется. В своей основе цикл полового размножения бессеменных сосудистых растений включает две фазы. В первую фазу возникает маленькое безлиственное растение, продуцирующее сперматозоиды, или яйцеклетки, или то и другое одновременно. Сперматозоиды от мужского растения перемещаются в воде (присутствующей хотя бы в виде тонкой пленки) и оплодотворяют яйцо женского растения, находящегося поблизости. Из оплодотворенного яйца развивается новое растение, превосходящее по размерам растения первой фазы и имеющее листья. Во вторую фазу это большее растение образует споры - клетки, служащие для размножения и не требующие оплодотворения. Они уносятся и рассеиваются ветром, укореняются, и из них развиваются мелкие безлиственные растения первой фазы. Таким образом завершается цикл. Как мы видим, один момент данного цикла требует присутствия воды; на ранних этапах истории наземных растений это ограничивало их распространение, так как размножение могло происходить лишь в определенных природных условиях.
Появление семян устранило это затруднение и таким образом усовершенствовало процесс размножения. При наличии семян отпала необходимость движения сперматозоидов к яйцеклетке вводе. Семя - это не просто изолированная клетка. Оно включает множество клеток, образующих в совокупности корень, стебель и один-два листка, "упакованных" в питательное вещество и снабженных оболочкой, защищающей их от механического и химического воздействия. Кроме того, когда семя начинает свое существование, оно уже содержит оплодотворенную яйцеклетку. Оплодотворение происходит тогда, когда яйцеклетка еще не отделилась от родительского растения, которое и питает семя на начальных стадиях роста. К тому времени, когда семя отделяется, оно уже представляет собой миниатюрный организм, снабженный "приспособительными" устройствами. Этот организм может сохраняться живым до тех пор, пока не укоренится и не начнет свою жизнь как самостоятельное растение. Благодаря указанным свойствам он имеет гораздо большие шансы выжить по сравнению с бессеменными растениями, что в свою очередь означает большее разнообразие возможных сред обитания и лучшую конкурентную способность данных видов. Значение таких преимуществ показывает вся история развития растений.
Древнейшие морские растения
Эволюция растений начинается с морских водорослей (см. табл. 2). Самые древние из известных на Земле ископаемых остатков (возраст которых около трех миллиардов лет) представляют собой наиболее примитивные растения - сине-зеленые водоросли. Почти два миллиарда лет такие растения обитали в морях вместе с другими одноклеточными организмами. Но, как свидетельствуют находки ископаемых в Австралии, около одного миллиарда лет назад существовали и зеленые водоросли. Наличие зеленых водорослей показывает, что к этому времени растения приобрели два важнейших свойства в отличие от их предков - более сложное строение и способность к половому размножению. Последнее свойство мы уже рассматривали. Поскольку каждый организм-потомок происходил уже не от одного, а от двух родителей, он обладал новой комбинацией генетического материала, что обусловило большую эффективность эволюции.
Усложнение структуры хотя и не было столь заметным усовершенствованием, также имело существенное значение для будущего. Когда первичные одноклеточные растения приобрели вторую клетку, а затем еще большее число клеток, их форма изменилась от почти шарообразной до линейной или неправильной. Возможное объяснение этого факта заключается в том, что любая несферическая форма имеет большее отношение площади к объему по сравнению с шаром. Таким образом, приобретение растением линейной или неправильной формы означало увеличение площади соприкосновения с морской водой, то есть площади поверхности, поглощавшей углекислый газ и солнечные лучи, необходимые для питания растений. Когда создались неправильности формы, из них могли развиться органы, выполняющие различные функции. И это должно было случиться раньше, чем водоросли могли из воды выйти на сушу.
Заселение суши растениями
До сих пор известны только две находки очень древних ископаемых наземных растений. В верхнесилурийских слоях Центральной Европы были найдены остатки безлистных растений длиной до 10 сантиметров; это самые древние из известных нам наземных растений. Вторая находка была сделана в Австралии, ц слоях раннедевонского возраста. Она представляет собой окаменевшие стебли (до 25 сантиметров длиной) растения, очень близкого к современным плаунам Lycopodium (фото 23), стебель которых стелется по земле и образует вертикальные отростки. Lycopodium - это не только наземное растение, это также и сосудистое растение. Наличие ископаемых растений такого типа означает не только существенный прогресс эволюции, но также и крупный географический сдвиг - выход растительной жизни из моря на сушу.
Фото 23. А. Древнейшее из известных нам наземных растений. Ископаемый отпечаток растения рода Baragwanathia в нижнедевонских слоях юго-восточной Австралии.
Совершенно ясно, что к позднесилурийскому времени растения уже заселили сушу. Но сколько времени занял этот процесс? Между появлением древнейших известных нам морских зеленых водорослей (около одного миллиарда лет назад) и древнейших наземных растений, обнаруженных в позднем силуре Европы (приблизительно 430 миллионов лет назад), прошло около 570 миллионов лет. Это действительно большой отрезок времени, почти равный продолжительности всего фаьерозоя. Хотя мы и признаем, что заселение суши мягкими морскими водорослями и развитие у них сосудистой системы были их замечательным! "завоеваниями", мы не должны забывать, что эти растения имели в своем распоряжении очень много времени, достаточно много для миллионов "проб и ошибок". Если вспомнить, что за такое длительное время даже ничтожная на первый взгляд вероятность почти наверное реализуется, то мы вправе усомниться в том, что водорослям действительно потребовалось так много времени (570 миллионов лет) для завоевания суши. Может быть, где-нибудь существуют кембрийские или даже более древние слои, содержащие остатки наземных растений, и если это так, то когда-нибудь они, вероятно, будут найдены и определены. И тогда сократится большой пробел в известной нам истории развития растений.

Фото 23. Б. Близкородственное ему современное растение, плаун Lycopodium lucidulum с побегами длиной 10-12 см
Путь из моря на сушу. Пытаясь заполнить этот пробел логическими рассуждениями, мы можем представить, каким образом произошел этот великий переход растений из моря на сушу. Водоросли, жившие в море, выделяли свободный кислород, который поступал в атмосферу, приближая постепенно ее состав к современному. Кислород (См. примечания к главе седьмой, стр. 106) создал экран, защищающий организмы от смертоносного излучения Солнца, и таким образом позволил им переселиться из моря на сушу, не подвергаясь опасности немедленной гибели. Этими переселенцами были растения, очевидно, зеленые водоросли. Они сами подготовили условия для этого переселения, выделяя в ходе многих лет процесса фотосинтеза огромные массы свободного кислорода, который превратил атмосферу Земли в защитный экран.
Естественно предположить, что зеленые водоросли расселялись вдоль побережий и проникали в устья рек. Через миллионы лет они, вероятно, распространились вверх по рекам, перейдя из соленой морской воды через солоноватую воду устьев рек в пресную речную. Мы можем представить себе, что временами медленные поднятия на некоторых участках суши преграждали путь рекам и создавали озера. С течением времени озера могли неоднократно изменять уровень и даже высыхать, а потом снова возникать. Непрекращающееся взаимодействие экзогенных и эндогенных процессов могло принимать тысячи различных форм.
Конечно, мы не знаем, сколько видов водорослей прошло этот путь. Но по крайней мере один из них выжил в условиях пересыхающего водоема. Живя в воде, водоросли поглощали пищу всей своей поверхностью. Но когда они стали "выходить" на сушу, им пришлось отказаться от этого простейшего способа питания. Первый шаг в этом направлении мог быть вызван опытом жизни в условиях временного пересыхания, возможно, в устьевой части речной долины во время отлива. У растений развился подобный коже покров, защищавший их от высыхания; в то же время этот новый покров делал возможным поступление воды в растения и ее диффузию.
Одновременно неровности тела растения должны были преобразоваться в примитивные органы со специализированными функциями. Один такой выступ, возможно нитевидный, мог использоваться для прикрепления к грунту, а также для поглощения воды и растворенных питательных веществ из грунта. Должны были также развиваться дополнительные сосудистые ткани, служившие опорой для тянущегося вверх растения. Иначе говоря, постепенно растение стало уже существенно отличаться от прежней мягкой водоросли. Оно становилось примитивным наземным растением, обладающим подобием корня, стеблем и простейшей сосудистой системой. Однако голые зеленые стебли и тонкие ветви, лишенные листьев, все еще напоминали водоросли.
К позднесилурийскому времени (возможно и раньше) такая стадия водорослеобразных растений была достигнута и пройдена. Как только закончился сложный процесс выхода растений на сушу, его естественным результатом явилось расселение и приспособление их к различным местным условиям, что должно было протекать сравнительно легко. Основные предпосылки для этого - наличие почвы, воды, минеральных питательных веществ в почве и углекислого газа в воздухе - уже существовали. Поверхность Земли, напоминавшая поверхность Луны, начала одеваться зеленым покровом.
Древняя наземная растительность; бессеменные растения. Некоторое представление о древнейшей растительности дают остатки окаменевших растений, встречающиеся местами в пластах горных пород, например в речных отложениях первой половины девонского времени. Эти растения принадлежат к нескольким различным родам, но все они относятся к примитивным группам, впоследствии исчезнувшим. Большая часть их имела стелющиеся по земле стебли с отростками, поднимавшимися на несколько десятков сантиметров, без листьев или с зачатками листьев. Такие растения не имели семян. По найденным ископаемым остаткам были сделаны реконструкции, одну из которых вы видите на фото 24.

Фото 24. Реконструкция раннедевонского ландшафта, на котором показаны растения нескольких родов, найденные в ископаемом состоянии в различных районах земного шара. Для масштаба показана дорожная сумка
По мере того как растения приспосабливались к различным условиям, они постепенно дифференцировались, образуя все большее количество видов. К концу девона, то есть 360 миллионов лет назад, растительность стала иметь уже совсем другой облик. Выделялись папоротниковидные растения, предки настоящих папоротников. Они не давали семян; некоторые из них достигали 12 метров в высоту и около одного метра в диаметре ствола у основания. Представив себе эти размеры, мы можем сказать, что первые леса на Земле начали возникать в позднедевонское время. Кроме древовидных растений, в этот период времени существовали крупные хвощи и плауны, потомки более мелких видов, живших ранее в девоне. На фото 25 изображена другая реконструированная картина, предположительно показывающая облик позднедевонского ландшафта.

Фото 25. Реконструкция ландшафта позднедевонского времени, на котором показаны настоящие папоротники (П), примитивные лепидодендроны - 'чешуйчатые деревья' (Ч) и меньшие по размеру растения
Позднепалеозойская флора; семенные растения. Следующим важным событием в истории растений было появление семян, на что ясно указывают ископаемые из каменноугольных пластов. Семена эти были довольно примитивными: они не имели оболочки и развивались на плодолистиках. Яйцеклетка семени оплодотворялась сперматозоидами того же самого растения. Среди древнейших растений, размножавшихся семенами, были семенные папоротники. Современные потомки других древнейших растений, размножавшихся семенами, - хвойные (сосна, ель, тсуга) - до сих пор размножаются таким же образом. Как и семенные папоротники, хвойные и другие подобные растения принадлежат к группе так называемых голосеменных, и все они имеют семена самого примитивного типа.
Однажды появившись, голосеменные растения уже не обязательно должны были держаться вблизи увлажненных мест, потому что для их способа размножения не требовалось даже тонкой пленки воды. Они занимали возвышенности, горы и сухие участки, ранее лишенные растений, и в результате этого зеленый растительный покров распространился по Земле еще больше.
Очевидно, раз голосеменные существовали в течение 350 миллионов лет и до сих пор еще очень многочисленны, их система размножения действует нормально. Но для того, чтобы она действовала, каждое голосеменное растение всегда должно производить большое количество семян и еще больше - пыльцы (пылевидных мужских половых клеток) для опыления. Размножение голосеменных зависит от ветра, разносящего их семена, но при этом многие семена попадают в неподходящие условия, - в воду, в пустыни, в места, слишком холодные для прорастания. Голосеменные растения затрачивают много энергии на производство семян и пыльцы, большая часть которых погибает. Впоследствии растения преодолели эту трудность, найдя более экономичный метод, но это произошло уже почти на 200 миллионов лет позднее, в меловой период.
Представьте себе условия, которые существовали в течение данного большого промежутка времени. На континентах в поздне-каменноугольное время преобладали обширные низменности с мягким климатом. В обширных болотах обильно произрастала пышная растительность; болотная вода покрывала отмершие части растений, предохраняла их от разложения и способствовала их сохранности вплоть до спрессования и превращения в уголь. Таким образом, в нашем представлении каменноугольная растительность - это главным образом богатая растительность влажных низменностей. Об обстановке, существовавшей в это время на холмистых возвышенностях, мы знаем мало, потому что более крутые склоны возвышенностей и отсутствие, как правило, на них стоячих вод не способствовали сохранности отмерших растений.
Болотистые низменности могли выглядеть так, как это изображено на фото 26, показывающем влажный лес. В таком лесу водилось много животных, подобных саламандрам, и насекомых. Такой тип болот, конечно, встречался не только в позднекаменноугольное время, он существовал и во все последующие эпохи. В настоящее время он часто встречается по берегам Мексиканского залива и вдоль Атлантического побережья Соединенных Штатов. Таковы в общих - чертах современные болота, хотя, конечно, большая часть растений болот принадлежит теперь к совершенно другим видам, чем в каменноугольный период.

Фото 26. Реконструкция болота позднекаменноугольного времени с обычной для таких болот пышной растительностью. П - папоротники, X - хвощи, Ч - лепидодендроны - 'чешуйчатые деревья', СП - семенные папоротники; видна также большая стрекоза. Некоторые хвощи достигали в высоту 30 м
Мезозойская растительность. Следующая из наших реконструкций относится к триасовому периоду и характеризует район национального парка "Каменный лес" в восточной Аризоне В действительности это совсем не лес. Это остатки леса, росшего Гдругом месте; они представляют собой скопление стволов деревьев, заключенных в толщу аргиллитов, из которой они сейчас постепенно высвобождаются в результате процессов выветривания (фото 27). За время долгого пребывания в толще породы стволы окаменели, их клеточная структура была замещена кремнеземом и превращена в агат - разновидность кварца. Стволы принадлежали хвойным деревьям (конечно, голосеменным), родственным современным соснам. При жизни деревья достигали 30-60 метров высоты, а стволы их имели толщину около двух метров. Мы знаем, что они росли не там, где находятся сейчас, потому что их поверхность носит следы истирания, а все ветви и корни обломаны. Это - плавник, принесенный в половодье откуда-то с возвышенностей в верховьях реки и погребенный в тонких аллювиальных отложениях. Он свидетельствует о том, что уже 220 миллионов лет назад на западе Северной Америки росли величественные рощи хвойных деревьев, точно такие же, как и те, которые значительно позже увидели европейцы, впервые проникшие на континент.

Фото 27. Огромные стволы триасовых хвойных деревьев, окаменевшие и превратившиеся в кварц. Нацианальный парк 'Каменный лес', Аризона
Фото 28 представляет собой реконструкцию растений, характерных для мезозойской эры. Среди них преобладают две группы голосеменных - хвойные и цикадовые, а также родственные им виды. Хвойные включали виды, подобные соснам и кипарисам, а также гинкго. Цикадовые, вероятно, развились из семенных папоротников. Их внешний облик напоминал пальмы, хотя эволюционной связи между этими группами нет; изящная крона из папоротниковидных листьев увенчивала ствол _ короткий, шарообразный или бочонковидный, а иногда высокий. Некоторые цикадовые сохранились до наших дней. Бессеменные растения, в первую очередь папоротники и хвощи, в мезозойское время играли менее заметную роль по сравнению с этими голосеменными.

Фото 28. Реконструкция общего вида растительности, типичной для мезозойской эры (только голосеменные и бессеменные растения). 7. Растения, родственные цикадовым. 2 Растения, родственные хвощам. 3. Папоротники. 4. Хвойные. Другие мезозойские ландшафты показаны также на фото 40, 43, 44, 45, 47
Покрытосеменные растения
Цветы и плоды. Длительная история растений ознаменовалась еще одним эволюционным усовершенствованием - появлением цветковых растений, или покрытосеменных. Это название дано им потому, что в отличие от голосеменных, у которых ничем не защищенные семена росли прямо на листьях; у этих растений семена, поодиночке или несколько вместе, заключены в особую оболочку, образующую внешнюю часть плода (например, яблоко). Кроме того, покрытосеменные имеют не только плоды, но и цветки. И то и другое приспособление обеспечивают лучшее размножение. Яркие цветки привлекают опыляющих их животных, например пчел, которые переносят пыльцу - мужские половые клетки; у более примитивных растений перенос пыльцы зависел от ветра. Плоды также привлекают животных, которые поедают их и таким образом разносят содержащиеся в них семена.
Цветы, плоды и другие усовершенствования позволили покрытосеменным растениями заселить те участки суши, которые еще оставались безжизненными, - участки с наиболее суровыми условиями существования. Этот процесс можно сравнить с покорением человеком вершины Эвереста, которое стало возможным лишь после того, как был создан ряд специальных приспособлений, в том числе кислородные приборы. Сейчас покрытосеменные составляют более 95% существующих видов сосудистых растений.
Мы не знаем, когда впервые появились покрытосеменные. Судя по ископаемым остаткам, они определенно встречаются в меловых слоях и, возможно, в юрских и даже в триасовых (остатки, содержащиеся в последних слоях, напоминают покрытосеменные растения, но у них не сохранились части, которые точно указывали бы на это). Во всяком случае, к середине мелового периода, около 100 миллионов лет назад, 90% видов растений, определенных в ископаемом состоянии, относились к покрытосеменным. К этой же группе относится большая часть современных лиственных деревьев, плодовых кустарников, овощей и цветов, трав и злаков. Список этих растений довольно внушителен, так как появление покрытосеменных знаменует переход от "средневекового" к современному растительному миру. Уже позднемеловые флоры имели современный облик, а существовавшие в начале кайнозоя (фото 48) деревья и другие растения показались бы современному человеку, если бы он их увидел, знакомыми.
Покрытосеменные растения и климат. Наиболее заметное различие между покрытосеменными и более ранней растительностью заключается в том, что покрытосеменные деревья и кустарники повсюду, за исключением тропической зоны, являются листопадными. После созревания семян они теряют листья, фотосинтез прекращается, и они на время, иногда на несколько месяцев, "засыпают". На основании этого ученые предположили, что появление покрытосеменных, по крайней мере отчасти, явилось результатом изменения климата континентов, повлиявшего на процесс эволюции. Если в мезозое климат приобретал все более резко выраженный сезонный характер, причем сезоны становились все более холодными и сухими, то опадение листьев и прекращение жизненной активности было бы вполне целесообразным. Обдумывая это предположение, мы должны вернуться к рисунку 27 и посмотреть, как происходило перемещение плит земной коры и движущихся с ними континентов в течение всего мезозоя. Северная Америка и по крайней мере европейская часть Евразии двигались к северу в более высокие и более холодные широты. Вполне возможно, что это предположение может объяснить появление листопадных видов растений в юрский и меловой периоды. Это не более чем предположение. Наука движется вперед, пытаясь связать воедино самые разнообразные данные о прошлом Земли. Именно так были установлены связи между отдельными событиями долгой истории Земли.
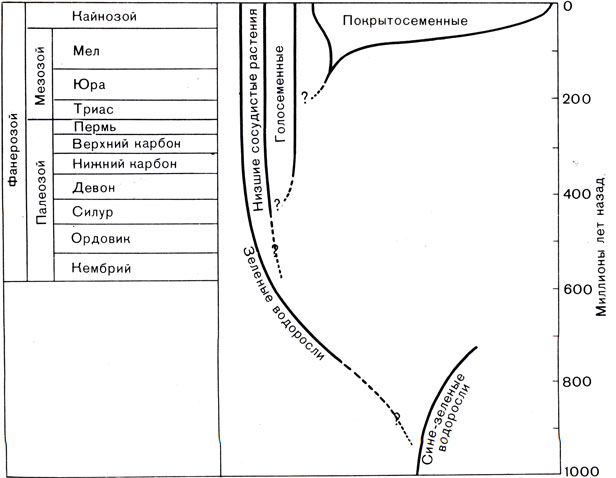
Рис. 39. Основные этапы геологической истории растений
Роль трав и злаков. Если судить по времени установления господства покрытосеменных, то растительность Земли приобрела современный облик еще до конца мезозойской эры, однако история развития растений на этом не кончается. Внутри самой группы покрытосеменных происходили существенные перемены, начавшиеся перед концом мелового периода и продолжавшиеся в течение большей части кайнозоя. Наибольший интерес для нас представляет развитие злаков.
Многие обитатели современных городов представляют себе травы только в виде газонов или площадок для гольфа. Но травы, в частности злаковые, - это самостоятельная группа покрытосеменных, которая приспособлена к жизни в засушливом или сезонно-засушливом климате. Родина злаковых трав - сухие равнины и степи с редким древостоем, называемые саваннами. Карта растительности (рис. 35) показывает, что почти вся западная часть центра Северной Америки представляет собой природную злаковую степь, не говоря уже об огромных площадях разреженной травянистой растительности в пустынных и полупустынных областях дальше к западу от штата Вашингтон до Мексики.
Возможно, благодаря распространению сухих климатов злаковые травы занимали все большую и большую площадь в течение кайнозоя. Имея съедобные стебли, листья и семена, злаковые травы являются идеальной высококалорийной пищей для животных, которые приспособлены к их поеданию. И как мы вправе были ожидать, быстро развивавшиеся в раннем кайнозое млекопитающие действительно поедали траву в большом количестве. Путем естественного отбора они все лучше и лучше приспосабливались к этому процессу. Форма их зубов и даже конечностей во многом определялась травяным рационом.
Хлебные злаки - это особый вид злаковых трав, в котором концентрация питательных веществ больше, чем в обычных злаках. Люди открыли высокую пищевую ценность хлебных злаков десять тысяч лет назад или еще раньше. Вскоре после этого люди начали сводить и расчищать леса, чтобы иметь возможность возделывать хлебные злаки. Еще позднее они начали повышать урожайность злаков путем отбора семян и искусственного выведения растений, а также путем применения удобрений и орошения. Сейчас большинство людей питается главным образом злаками: рисом, пшеницей, кукурузой и другими зерновыми. Десять тысяч лет - это короткий промежуток времени в истории Земли, чтобы питание повлияло на строение человеческого тела.
Прежде чем мы перейдем к рассмотрению эволюции животного мира, целесообразно восстановить основные события истории развития растений, помня о том, что без растений животный мир не смог бы успешно развиваться на суше. В результате изучения ископаемых остатков можно установить следующие основные моменты в развитии растений:
1. Выход на сушу морских водорослей; примитивные наземные растения.
2. Бессеменные растения. Папоротники, хвощи.
3. Появление семян. Голосеменные.
4. Появление покрытосеменных, характеризующихся наличием цветов и плодов.
5. Развитие злаковых трав и позднее хлебных злаков.
Эволюционные соотношения между основными группами растений и хронологическая шкала истории развития растений приведены на рисунке 39.
Литература
Andrews H. N.. Jr., 1964. Ancient plants and the world they lived in: Cornell University Press, Ithaca. N. Y.
Darrah W. C, 1960, Principles of paleobotany: The Ronald Press Co., New York.
Delevoryas Theodore, 1966, Plant diversification: Holt, Rinchart and Winston, New York. (Paperback.)
Scientific American Staff, eds.; 1957, Plant life: Simon and Schuster, Inc.. New York. (Paperback.)
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'