
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Ледниковые убежища и темпы расселения растений
Вопрос о возможности жизни высших растений и животных в горах во время оледенений давно привлекал внимание ученых разных специальностей. Особенно много материалов накоплено по Скандинавскому нагорью, где во время последнего оледенения в принципе могли существовать небольшие участки, свободные от льда. Впервые такое предположение было высказано в конце XIX в. норвежским ботаником А. Блиттом и шведским зоологом Р. Сернандером. В качестве наиболее подходящих мест для ледниковых убежищ, или рефугиумов, скандинавские исследователи отмечали высокогорные районы. Здесь были обнаружены изолированные местообитания некоторых видов растений и животных, удаленные от основного ареала либо представляющие собой разобщенные части ареала.
Среди растений таких видов насчитывается около 30. Многие из них концентрируются в двух районах нагорья: в массивах Ютунхеймен, Доврефьелль и Тролльхеймен на юге и от долины Сальтдаль до западного Финнмарка на севере. Виды, встречающиеся в обоих районах, например лапландский рододендрон, называют бицентричными, а встречающиеся только в одном районе - уницентричными.
Концепция ледниковых убежищ не избежала строгой критики. Ее острие было направлено против использования для палеогеографических реконструкций современных ареалов. В самом деле, разобщенные местонахождения растений и животных в горах можно рассматривать не только как ледниковые убежища, но и как результат образования островных ареалов в связи с похолоданием в позднем голоцене.
Помимо собственно горных убежищ, Р. Сернандер, Р. Нурхаген и другие исследователи выделяли прибрежные убежища, не покрывавшиеся льдом во время оледенений. К таким участкам были отнесены побережье Мере, некоторые из Лофотенских островов, берег Финнмарка и др. (рис. 18). Тем не менее высказывались и сомнения по поводу существования прибрежных убежищ. Шведский ботаник X. Шёрс отметил, например, отсутствие веских фактических аргументов палеонтологического характера и недостаточность использования современных ареалов.
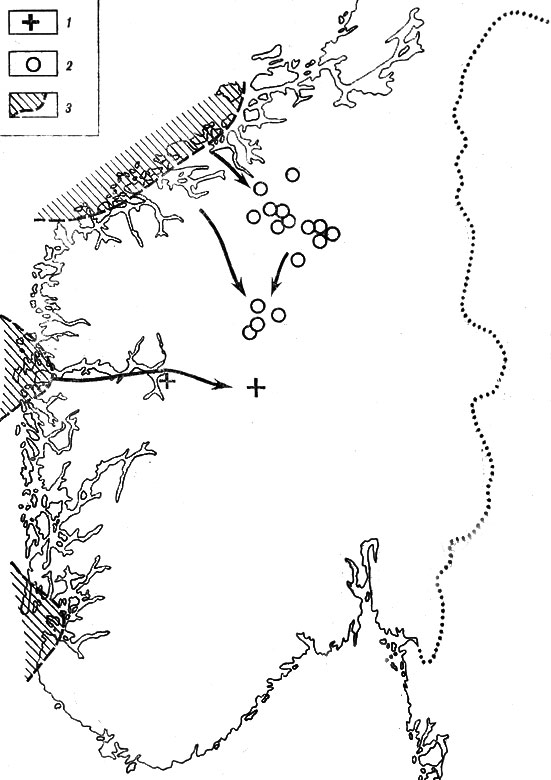
Рис. 18. Распространение горных маков в Норвегии (по Р. Нурхагену). 1 - Papaver relictum; 2 - P. Nordhagenianum subsp, ovatilobum; 3 - предполагаемые убежища ледникового времени
В ряде пунктов на западном побережье Норвегии с помощью спорово-пыльцевого анализа было установлено существование богатой и разнообразной арктоальпийской флоры в позднеледниковое время и в раннем голоцене, задолго до того как появилась возможность для миграции растений по суше. Наиболее вероятно, что в ту пору семена растений заносились с запада морскими течениями, ветром и птицами. Таким образом, предположение о существовании убежищ в ледниковую эпоху не подтверждается с биогеографических позиций.
Геологи и географы полагали, что во время оледенений мощность льдов уменьшалась по направлению к западному побережью Северной Скандинавии, к которому спускались только языки выводных ледников. Между этими языками якобы оставались свободные от льда участки. Непосредственно в пределах Скандинавского нагорья убежища могли быть приурочены к скалистым вершинам и гребням, поднятым над выровненной поверхностью основной части нагорья. Высота этой поверхности составляет 1000-1200 м над уровнем моря, тогда как сильно расчлененные скальные гребни и вершины возвышаются до 1500-2000 м и более.
Обширные валунные поля, встречающиеся на основной выровненной поверхности нагорья, особенно в его западной части, образовались в результате частой смены процессов таяния и замерзания. Для этого, однако, вовсе не требовалось много времени: судя по нынешним быстрым темпам морозного выветривания, каменные россыпи вполне могли сформироваться в голоцене. Во всяком случае, нет никаких поводов относить их именно к эпохе оледенения.
На востоке нагорья развит покров основной морены. Изучение состава встречающихся в ней валунов показало, что эти обломки были принесены не местными ледниками, а мощным ледниковым покровом, местами поднимавшимся до 1500 м над уровнем моря. Вершины и гребни, находящиеся выше этой отметки, и в настоящее время покрыты льдом и снегом. Следовательно, вряд ли они могли представлять собой обнаженные скалы в эпоху оледенения. Итак, возникает противоречие между фактами, указывающими на сплошное распространение последнего ледникового покрова, и биогеографической аргументацией в пользу существования убежищ для некоторых видов растений и животных. Правда, по мнению шведского географа Р. Даля, участки наибольшего развития оледенения могли перемещаться во времени и пространстве. Эта концепция представляется довольно логичной. Во всяком случае, она помогает объяснить, каким образом на территории нагорья на начальных и заключительных стадиях оледенения могли оставаться свободные ото льда участки.
Помимо убежищ в горных странах, несомненно, существовали убежища на сопредельных равнинах и в предгорьях, расположенные поодаль от Европейского ледникового покрова. Во время деградации оледенения и в послеледниковое время из этих очагов происходило длительное расселение растений и животных. Чтобы составить представление о ходе данного процесса, ученые обратились к анализу остатков организмов, сохранившихся в ископаемом состоянии, чаще всего в болотных и озерных отложениях. Для опознавания этих остатков применялись палеонтологические методы. Особенно эффективную помощь оказал спорово-пыльцевой анализ, обеспечивающий наибольшую полноту информации. Возраст устанавливался радиоуглеродным методом.
Для примера рассмотрим последовательность распространения ели обыкновенной - характерной древесной породы европейской тайги (рис. 19). Во время последнего оледенения она произрастала в более южных районах Русской равнины, а в центральных районах уступала место холодовыносливой сибирской ели, которая растет даже там, где существует многолетняя мерзлота.
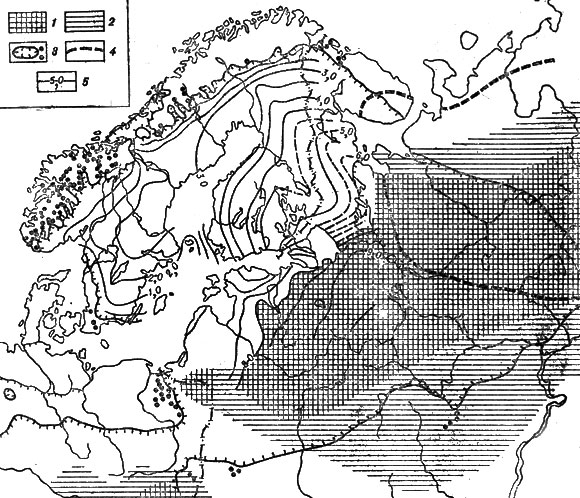
Рис. 19. Распространение ели на востоке и севере Европы. 1 - основной ареал ели около 8 тыс. лет назад; 2 - разреженные островные местообитания ели около 8 тыс. лет назад; 3 - граница современного ареала и островные местообитания ели европейской; 4 - граница современного ареала ели сибирской; 5 - изохроны появления ели (возраст по 14С указан в тыс. лет)
Улучшение климата в самом конце ледниковой эпохи вызвало миграцию ели обыкновенной к северу, и около 10-8 тыс. лет назад она постепенно заселила центральные районы Русской равнины, хотя тогда она еще не играла видную роль в составе лесов. Одновременно ель сибирская отступила в северо-восточные районы, К концу указанного интервала времени ель обыкновенная проникла в восточную часть Прибалтики и на юг Онего-Ладожского перешейка.
За следующие полторы тысячи лет (8-6,5 тыс. лет назад) темпы расселения ели на севере уменьшились, и только на западе Прибалтики и на севере Польши распространение ели еще продолжалось. Именно в это время, по-видимому, произошло слияние двух миграционных потоков ели с юга, со стороны Карпат, и с востока, со стороны Русской равнины.
Во время послеледникового климатического оптимума (6,5-5,8 тыс. лет назад) появились наиболее благоприятные условия для увеличения ели в лесах и расширения ее ареала. Эта порода проникла в северные районы Эстонии, и тем самым завершился процесс ее распространения на северо-западной окраине Русской равнины.
На территории Финляндии и Швеции ель мигрировала очень быстрыми темпами. Примерно 3,6-3,3 тыс. лет назад эта порода появилась в Центральной Лапландии, а еще спустя несколько столетий достигла подножий Скандинавского нагорья. Расселение ели в центральные и южные районы Швеции произошло в самые последние тысячелетия и продолжается до сих пор.
Приведенные факты показывают, что ель в послеледниковое время мигрировала из центральных районов Русской равнины на северо-запад. Для других пород в пределах той же территории были выявлены несколько иные пути расселения (рис. 20). Так, например, липа в послеледниковое время продвигалась двумя разными путями - на север вдоль восточного побережья Балтийского моря и на северо-запад из Подмосковья по возвышенностям в бассейнах рек Тверцы и Мсты к побережью Финского залива. Спустя 2,5 тыс. лет оба миграционных потока соединились на севере Эстонии, тогда как на обширной промежуточной территории в бассейне Чудского и Псковского озер расселение липы происходило с большим запозданием.
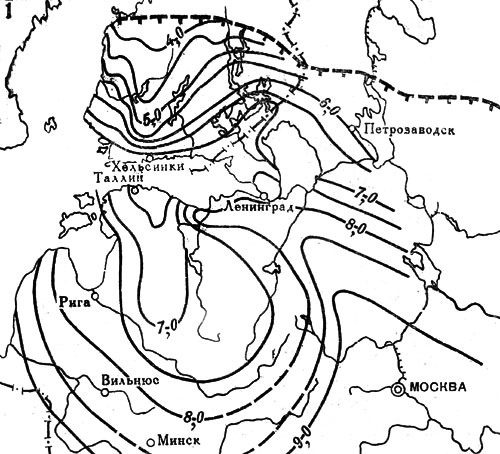
Рис. 20. Распространение широколиственных пород на северо-западе СССР и юге Финляндии в послеледниковое время. Изохроны появления: 1 - липы; 2 - вяза; 3 - дуба. Изохроны начала широкого распространения в лесах: 4 - липы; 5 - вяза; 6 - дуба. Цифрами обозначены значения возраста (в тыс. лет) от 1950 г. Штриховыми линиями с подсечками выделены северные границы современных ареалов соответствующих пород
Несмотря на очень быстрые темпы распространения в послеледниковое время теплолюбивых широколиственных пород - липы, дуба, вяза, выяснилось, что даже во время климатического оптимума они не проникали далеко на север Европы, за пределы своих нынешних ареалов. Северных границ эти породы деревьев достигли около 4,5-4,0 тыс. лет назад, и с тех пор имели место только небольшие осцилляции. Такой вывод расходится с представлениями о продвижении теплолюбивых древесных пород на территории Сибири к берегам морей Северного Ледовитого океана во время послеледникового климатического оптимума и о последующем возвращении их в пределы современных ареалов.
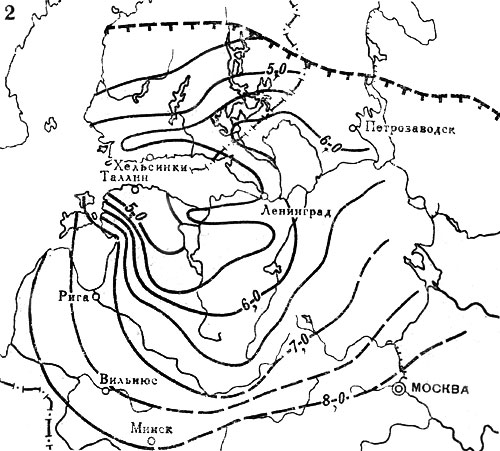
Рис. 20. Распространение широколиственных пород на северо-западе СССР и юге Финляндии в послеледниковое время. Изохроны появления: 2 - вяза. Цифрами обозначены значения возраста (в тыс. лет) от 1950 г. Штриховыми линиями с подсечками выделены северные границы современных ареалов соответствующих пород
Чтобы представить себе общую направленность миграций растительности на территории Русской равнины в голоцене, обратимся к исследованиям, выполненным в центрально-черноземных областях с применением спорово-пыльцевого и радиоуглеродного анализов.
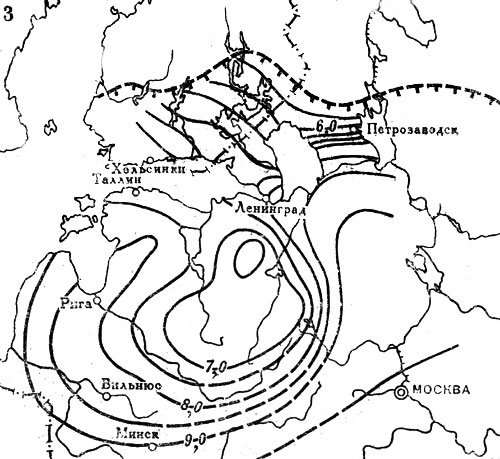
Рис. 20. Распространение широколиственных пород на северо-западе СССР и юге Финляндии в послеледниковое время. Изохроны появления: 3 - дуба. Цифрами обозначены значения возраста (в тыс. лет) от 1950 г. Штриховыми линиями с подсечками выделены северные границы современных ареалов соответствующих пород
Во время последнего плейстоценового оледенения на возвышенных участках этой территории распространялись перигляциальные травянистые сообщества в сочетании с зарослями ерника (кустарниковых берез) и ольховника. Структура растительного покрова отличалась мозаичностью и во многом зависела от пространственного распределения разных типов многолетнемерзлых грунтов. Однако по крупным долинам даже в это суровое время произрастали редкостойные таежные леса из сосны, ели, лиственницы и березы. Подобную ярусность растительности с лесами по долинным таликам и кустарниково-травянистыми сообществами на плакорах в настоящее время можно наблюдать на предгорных равнинах в Восточной Якутии.
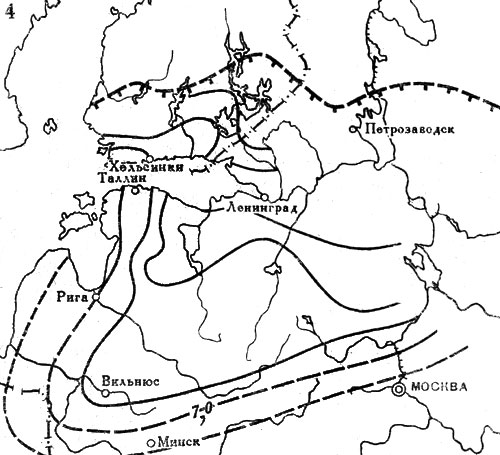
Рис. 20. Распространение широколиственных пород на северо-западе СССР и юге Финляндии в послеледниковое время. Изохроны начала широкого распространения в лесах: 4 - липы. Цифрами обозначены значения возраста (в тыс. лет) от 1950 г. Штриховыми линиями с подсечками выделены северные границы современных ареалов соответствующих пород
Улучшение климата в конце плейстоцена вызвало таяние многолетней мерзлоты и распространение переувлажненных грунтов в перигляциальной зоне Русской равнины. В составе травянистых сообществ того времени было много осок. В более сухих местообитаниях в изобилии росли полыни и маревые. Значительно улучшились условия произрастания древесных пород, однако редколесья все еще концентрировались преимущественно по речным долинам. Основной фон растительного покрова составляли перигляциальные кустарниково-травянистые сообщества. Пространственная структура растительности претерпевала изменения под влиянием климатических пульсаций. Похолодания способствовали большему распространению открытых перигляциальных ландшафтов, потепления стимулировали расширение площадей редкостойных лесов.
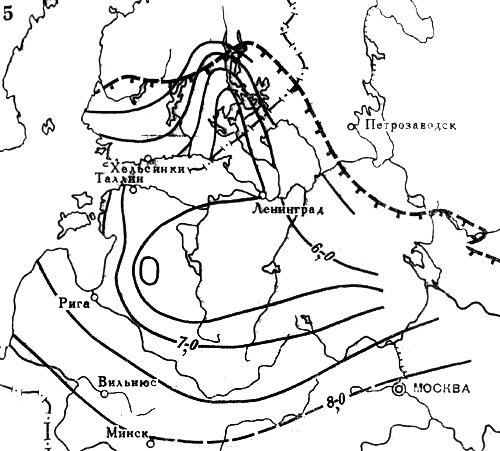
Рис. 20. Распространение широколиственных пород на северо-западе СССР и юге Финляндии в послеледниковое время. Изохроны начала широкого распространения в лесах: 5 - вяза. Цифрами обозначены значения возраста (в тыс. лет) от 1950 г. Штриховыми линиями с подсечками выделены северные границы современных ареалов соответствующих пород
Процесс перестройки перигляциальной растительности закончился в раннем голоцене, когда установилась широтная дифференциация растительных зон, близкая к современной. В бореальный период (9-8 тыс. лет назад) на северо-западе Среднерусской возвышенности произрастали сосновые и березовые леса. По направлению к югу и юго-востоку в лесах наряду с сосной и березой становилось больше широколиственных пород. На севере возвышенности среди этих пород в первую очередь появлялись липа и вяз, на юго-западе и западе главную роль играл дуб. Проникновение широколиственных пород происходило с запада и юго-запада, из Подольского рефугиума термофильной флоры. Южная граница лесной зоны в бореальное время пересекала центральную часть Среднерусской возвышенности по линии Железногорск - Кромы. К юго-востоку от этого рубежа возрастала роль лугово-степных сообществ, которые в сочетании с массивами лесов формировали зону лесостепи, сменявшуюся на юге возвышенности степной зоной.
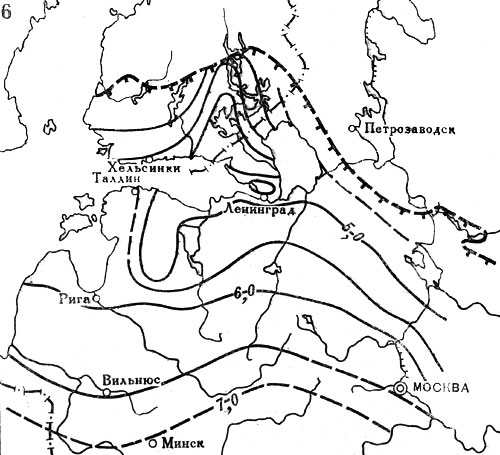
Рис. 20. Распространение широколиственных пород на северо-западе СССР и юге Финляндии в послеледниковое время. Изохроны начала широкого распространения в лесах: 6 - дуба. Цифрами обозначены значения возраста (в тыс. лет) от 1950 г. Штриховыми линиями с подсечками выделены северные границы современных ареалов соответствующих пород
В атлантический период (7-5 тыс. лет назад) вся лесная зона на Русской равнине значительно продвигалась к северу, соответственно в том же направлении смещалась и степная зона. При этом лесостепная зона значительно сужалась, поскольку положение южной границы лесной зоны сравнительно мало менялось. Сходный вывод о стабильности южной границы лесной зоны в атлантический период сделан и для юга Западной Сибири.
Палеоботанические данные свидетельствуют, что в это время северные и западные районы Среднерусской возвышенности были заняты лесами с большой примесью широколиственных пород: на севере липы, на западе дуба. Вся южная половина возвышенности была занята степями, и северная граница степной зоны доходила до верховьев Оки.
Что же лежит в основе миграции растительных зон в эпоху климатического оптимума голоцена? Ведущую роль здесь, по-видимому, играл температурный фактор. Повышение температуры в разных условиях произвело неоднозначный эффект: в гумидных районах оно не сопровождалось критическим изменением относительной влажности, а в аридных привело к резкому увеличению дефицита увлажнения. Реакция растительности на эти изменения и определила отмеченное выше перемещение зон в северном направлении. Стимулом для движения степей к северу могло быть даже небольшое повышение температуры.
Суббореальный период (5-3 тыс. лет назад) ознаменовался новыми тенденциями в развитии растительности, что диктовалось планетарными климатическими изменениями. Признаки наступания леса на степь приобрели отчетливое выражение со второй половины суббореального периода. Граница лесной зоны в бассейне Верхней Оки тогда смещалась к югу примерно на 70 км, соответственно в том же направлении мигрировала и зона лесостепи, причем даже в южных районах Среднерусской возвышенности по речным долинам распространялись значительные массивы ольшаников и широколиственных лесов. Таким образом, палеоботанические данные однозначно указывают на нарастание увлажненности во второй половине суббореального периода. Мнение о сухом и теплом климате этого интервала, разделявшееся некоторыми исследователями, не подтверждается.
В субатлантический период в обстановке еще более влажного и прохладного климата продолжалось продвижение лесной растительности к югу. Широкое распространение ели в лесных районах центра Русской равнины затронуло северную и северо-западную окраины Среднерусской возвышенности. На ее крайнем западе по долинам росли сосновые боры, а на севере - липовые и дубово-липовые леса. В центральных и южных районах встречались дубравы, сочетавшиеся с разнотравно-злаковыми и злаково-разнотравными сообществами, создавая характерную мозаичность лесостепной растительности. Типичные степи были оттеснены на юго-восток возвышенности.
В субатлантическое время большое влияние на развитие растительности оказала деятельность человека. Можно выделить несколько этапов рубки лесов в связи с развитием подсечно-огневого земледелия, особенно в начале и конце I тысячелетия н. э. Однако наиболее значительные преобразования ландшафта произошли за последние 200- 300 лет, в эпоху сплошной распашки земель.
Выявленные по палеоботаническим и радиоуглеродным данным смены растительности на территории Среднерусской возвышенности в голоцене вполне согласуются и с материалами зарубежных исследователей. Например, существование степного периода в середине голоцена и последующее наступание леса на степь установлено на территории Венгрии, Чехословакии, США. В этих исследованиях можно усмотреть модель растительных смен во время межледниковий плейстоцена. Тем самым данные о расширении лесной зоны к северу и югу за пределы занимаемого ею ныне ареала во время климатических оптимумов межледниковий вызывают сомнение.
Проблема ледниковых убежищ флоры и фауны детально изучалась во многих горных странах, в частности на Большом Кавказе, где размеры последнего плейстоценового оледенения были довольно велики. Морфологические следы его четко выражены в характере продольных и поперечных профилей крупных долин, в выположенности верхнего яруса гор, в системе краевых ледниковых образований. Помимо облекающего покрова льда и фирна на высотах более 2300-2400 м многие кавказские долины были заняты ледниками, и оледенение местами приобретало сетчатый характер.
Во время максимального развития оледенения лесной пояс на Кавказе был вытеснен в предгорные и равнинные районы. Перигляциальные растительные сообщества могли тогда спускаться до подножий гор, а лесная растительность соответственно мигрировала на подгорные равнины. Изучение местонахождений реликтовых и эндемичных видов растений указывает на существование убежищ в окрестностях Эльбруса, аридных котловинах Балкарии, Лескенских лесах, на Бечасынском плато и в некоторых других районах. Некоторым из этих убежищ приписывается очень молодой, голоценовый возраст, другим - более древний, ледниковый и даже плиоценовый (Лескенские леса).
С помощью спорово-пыльцевого анализа, проведенного географом Е. С. Малясовой для озерно-болотных отложений урочища Криют в Юрской продольной депрессии, удалось доказать, что здесь на высотах 2100-2200 м в раннем голоцене произрастали субальпийские редколесья с большой примесью бука, граба, дуба, хотя основной фон составляли береза и ольха. Судя по составу спорово-пыльцевых спектров, поблизости, но, конечно, на меньших высотах располагался пояс буковых лесов с примесью других широколиственных пород.
В конце раннего голоцена, на рубеже бореального и атлантического периода (около 8 тыс. лет назад), Безенгийский ледник спускался примерно на 12 км ниже, чем в настоящее время. Между тем буковые леса ныне вообще не заходят в Безенгийскую долину. Приведенные факты свидетельствуют, что в самом конце ледниковой эпохи в данном районе существовало убежище лесной флоры. По-видимому, на фоне происходившего тогда улучшения климата лесная растительность расселялась вверх по долинам вслед за отступавшими ледниками.
Во время климатического оптимума голоцена (атлантический период) растительные пояса быстро перемещались вверх, и в итоге на северном макросклоне Центрального Кавказа буковые леса с участием граба и дуба поднялись до 2150-2200 м над уровнем моря. Верхняя граница субальпийского березового редколесья находилась на уровне 2350-2400 м. Далее над поясом редколесий расстилались высокогорные луга. Амплитуда вертикальных миграций отдельных растительных поясов в атлантический период оценивается нами примерно в 300 м.
С середины суббореального периода (около 4 тыс. лет назад) на фоне понижения температуры проявилась нисходящая миграция растительных поясов, усугубившаяся благодаря возросшей увлажненности в субатлантический период. Преобразованию высокогорных ландшафтов способствовало их хозяйственное освоение. Этот процесс, вероятно, начался в конце I тысячелетия н. э. и усилился за последние 150-200 лет. Масштабы антропогенных процессов и их направленность, конечно, отличались в разных районах высокогорного Кавказа, однако там, где проводился неумеренный выпас скота, естественный растительный покров претерпел существенные изменения.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'