
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Вымирание животных в позднем плейстоцене
Вопрос о вымирании организмов во время материковых оледенений заслуживает особого внимания. Эту, казалось бы, совершенно очевидную причинную зависимость не просто подтвердить на фактическом материале, поскольку необходимо располагать точными датировками поэтапного распространения ледников и связанного с ним резкого похолодания и исчезновения соответствующих видов. Более того, не всегда можно провести обоснованные и четкие сопоставления между вымиранием организмов и изменениями природного окружения.
В принципе оледенение должно было сопровождаться ухудшением среды обитания животных на перигляциальной суше. Оледенение не только непосредственно вело к гибели растений и животных, но и создавало неблагоприятные условия для их существования: перенаселенность отдельных местообитаний, обезлесение, новые отношения к разным врагам и конкурентам, нарушения в ритме репродукции и т. д. B поисках лучших условий многие животные покидали прежние местообитания, и в процессе этих миграций значительная часть популяций погибала. Тем не менее размеры области отрицательного влияния оледенения на органический мир многими исследователями, вероятно, сильно преувеличиваются.
Существуют разные точки зрения на причины гибели крупных животных в ледниковую эпоху. Одни ученые полагают, что такой причиной было само оледенение. Известный геолог В. И. Громов, например, связывал наиболее критический момент в развитии органического мира с максимальным - рисским оледенением. Другие исследователи, в частности авторитетный зоолог И. Г. Пидопличко, допускали, что современные степные животные Юга СССР непосредственно связаны с плиоценовой фауной в пределах своих нынешних ареалов, причем многие из них встречаются в отложениях всего четвертичного периода. Вместе с тем ученый признавал влияние изменений климата в четвертичном периоде на развитие фауны, но основную роль в ее преобразовании отводил деятельности человека. Такую же позицию занимали и другие исследователи.
Истоки этой дискуссии восходят еще ко второй половине прошлого века, когда впервые были получены сведения о составе фауны в конце последнего материкового оледенения. Естественно, эти данные ставились в зависимость от изменений климата и ледников. Такое объяснение, однако, не получило всеобщего признания, особенно после того, как Ч. Дарвин во время кругосветного путешествия на "Бигле" обратил внимание, что в некоторых местностях остатки вымерших животных концентрируются в слоях, имеющих послеледниковый возраст. На этом основании английский исследователь А. Уоллес в качестве основной причины массовой гибели животных назвал деятельность первобытного человека.
Как известно, на протяжении плейстоцена состав фауны менялся неоднократно, но каждый раз освобождавшиеся экологические ниши занимали новые представители того же рода, так что общий баланс биосферы не нарушался. В конце плейстоцена на земном шаре произошло колоссальное по своим масштабам исчезновение крупных млекопитающих. По подсчетам американского ученого П. Мартина, в это время погибло не менее 200 родов, причем главным образом крупных травоядных и трофически связанных с ними хищников. Благодаря применению радиоуглеродного метода остатки этих животных в ряде мест были датированы, и оказалось, что вымирание фауны совпало со временем распространения культур позднепалеолитических охотников, специализировавшихся на добыче крупных зверей.
Впоследствии сокращение состава фауны крупных млекопитающих частично восполнилось за счет увеличения биомассы мелких млекопитающих и насекомых, однако общий баланс экосистем был существенно нарушен и, несмотря на возросшее поголовье домашнего скота, не восстановился.
Любопытно заметить, что в фауне морских беспозвоночных, являющейся признанным индикатором изменений биосферы, около 11 тыс. лет назад произошли резкие перемены: холодовыносливые комплексы уступили место более теплолюбивым. Однако вымирание, тем более на уровне родов, не было зафиксировано.
Несмотря на крупные масштабы миграций растительных зон и поясов, позднеплейстоценовое вымирание фактически не затронуло флору. Благодаря применению спорово-пыльцевого и диатомового анализов были довольно детально изучены сложные смены растительных формаций. Свидетельствами вымирания родов диатомей или высших растений в позднем плейстоцене мы не располагаем, хотя в более ранние этапы плейстоцена, например, в средней полосе Европы установлено исчезновение многих родов растений (магнолия, секвойя, лапина и др.), которые ныне сохранились в отдельных районах Восточной Азии и Северной Америки.
Масштабы позднеплейстоценового вымирания фауны крупных млекопитающих в Европе были меньше, чем в других частях света. Для сравнения напомним об исчезновении гигантских сумчатых в Австралии, гигантских лемуров на Мадагаскаре и, наконец, огромной группы млекопитающих в Северной Америке (не менее 28 видов, входивших в позднеплейстоценовый ранчолабрейский комплекс: мастодонт, мамонт, колумбов слон, верблюд, лошадь, гигантский броненосец, гигантский мегатерий, тапир и др.).
Тем не менее и для Европы видовой состав, а также обилие позднеплейстоценовых фаун млекопитающих наряду с более крупными размерами особей убедительно свидетельствуют о состоянии естественного климакса в развитии животного мира. Положение весьма быстро изменилось во время интенсивного таяния последнего ледникового покрова.
В более ранние этапы плейстоцена фауна млекопитающих Европы неоднократно испытывала изменения с исчезновением теплолюбивых форм. Причем помимо общих филогенетических факторов, несомненно, активно проявлялись смены биотопов, обусловленные динамикой оледенения. Следы этого влияния, в частности, видны в разобщенности ареалов некоторых мелких млекопитающих. В холодные ледниковые интервалы эти животные укрывались в убежищах на юге вноледниковой территории. Нередко подобная изоляция сопровождалась образованием подвидов и даже видов. Так, из выхухолей (Desmanidae), широко распространенных в Европе в плиоцене и раннем плейстоцене, до настоящего времени сохранились два территориально разобщенных вида - пиренейский выхухоль (Desmana pyrenaica) на севере Пиренейского полуострова и европейский выхухоль (D. moschata) в южных и юго-восточных районах Русской равнины. Ископаемые остатки европейского выхухоля с позднего вюрма известны из пещерных отложений Центральной Европы.
Период наибольшего распространения последнего материкового оледенения характеризовался усилением роли холодостойких млекопитающих. Они имели благоприятные предпосылки для широкого расселения на равнинах Европы в условиях сокращения лесных биотопов и развития многолетней мерзлоты. Одновременно более теплолюбивые виды мигрировали далеко на юг, в средиземноморские районы. Именно к данному периоду относится расселение вплоть до Северного Причерноморья холодовыносливых млекопитающих, включая мамонта, северного оленя, песца и копытного лемминга.
Деградация последнего оледенения в средней полосе Европы сопровождалась повышением роли лесных биотопов и их последовательным продвижением к северу. К этому же времени относится расцвет охотничьих культур поздних стадий палеолита, специализировавшихся на добыче крупных травоядных млекопитающих. Люди вступили в единоборство с неблагоприятными природными условиями, обнаружив удивительную адаптацию к суровой обстановке; фактически началось активное преобразующее влияние человека на животный мир.
Надо заметить, что крупные звери составляли один из наиболее существенных компонентов природного окружения древнего человека, что и определило раннее развитие охотничьего промысла, но только в позднем палеолите охота заняла важную позицию в хозяйственной жизни. Об этом свидетельствуют произведения палеолитического искусства, посвященные в основном изображению оленей, лошадей, бизонов, зубров, носорогов, мамонтов, которые играли важную роль в пищевом рационе. Наряду с ними изображались и хищные животные - соперники по охоте и враги первобытных людей.
Ко времени деградации последнего оледенения относят вымирание большой группы крупных млекопитающих Европы, в том числе мамонта, волосатого носорога, овцебыка, большерогого оленя, пещерного медведя, пещерного льва и пещерной гиены. Видимо, тогда на большей части Европы исчезла дикая лошадь. Вместе с антилопой-сайгой она была постепенно оттеснена во внутренние районы Азии. В конце позднеледниковьи - начале послеледниковья северный олень мигрировал на север Европы. Сильно сократились популяции многих видов крупных млекопитающих, например первобытного зубра. Рассмотрим особенности позднеплейстоценовой истории некоторых типичных видов млекопитающих.
Мамонт настоящий (Mammuthus primigenius Blum.), произошедший от степного слона (М. trogontherii Pohl.), сложился во время рисского оледенения. В вюрмское время мамонты широко распространились во всей Северной Евразии. Первые сведения об ископаемых мамонтах появились еще в середине I тысячелетия до н. э. Долгое время бивни этих животных были ходовым меновым товаром в Сибири. По сохранившейся статистике можно составить некоторое представление о количестве самих находок. Например, в 1903 г. в Якутске было продано 57,6 тыс. фунтов бивней мамонтов, что, вероятно, соответствует 50 тыс. особей, В многолетнемерзлых толщах Сибири, помимо костных остатков, сохранились целые трупы мамонтов. В Старуни, в северных предгорьях Карпат, найден труп мамонта, захороненный в алевритах и пропитанный нефтью и солями.
Об адаптации мамонта к условиям холодного климата свидетельствуют его длинный волосяной покров, небольшие уши и бивни, с помощью которых он разгребал снег в поисках корма (рис. 22), Как отмечалось выше, он мог существовать и в условиях умеренной зоны. Судя по геологическому положению ископаемых остатков мамонта я характеру флоры и фауны, это крупное травоядное животное предпочитало открытые безлесные биотопы, Мамонт расселялся в вюрме в пределах всей Европы, от Крайнего Севера до Закавказья. Миграции мамонтов, вероятно, подчинялись определенным климатическим ритмам.

Рис 22. Мамонты в перигляциальной обстановке (рисунок А. Ван-Олдена)
Данные радиоуглеродного метода позволили установить, что в длительный предледниковый период (40-25 тыс. лет назад) мамонт был наиболее широко распространен, в конце последнего оледенения (13-9 тыс. лет назад) животные встречались реже. Несомненно, они вымерли в самом начале послеледникового времени, когда произошло резкое изменение климата. Мамонт, найденный в районе Хельсинки, имеет возраст около 9 тыс. лет.
Наиболее оптимальные условия для обитания мамонтов в Евразии в предледниковый период доказываются результатами сравнительно-морфологических исследований А. В, Гейнца и В. Е. Гарутта. Именно в этот период мамонты были наиболее крупными, что связывается о обилием кормовых ресурсов.
Мамонты, жившие в эпоху наступания последнего ледникового покрова, отличались гораздо меньшими размерами. Довольно большая группа радиоуглеродных датировок порядка 24-18 тыс. лет назад установлена для ископаемых остатков мамонтов, обнаруженных на юге Скандинавского нагорья, а также на севере Белоруссии и в восточных предгорьях Альп.
Во время максимального развития оледенения (16-14 тыс. лет назад) сильное похолодание могло привести к изменению экологических условий и вымиранию значительной части поголовья мамонтов на севере Евразии. Уцелевшие группы животных мигрировали в более южные районы, откуда вернулись на север только в самом конце ледниковой эпохи. Именно с этим интервалом, по всей вероятности, были связаны последние страницы истории мамонтов.
Роль первобытных охотников в истреблении гигантских травоядных животных была достаточно велика. Об этом свидетельствуют хотя бы произведения первобытного искусства конца палеолита, запечатлевшие загонную охоту и применение западней. Колоссальные скопления костей мамонтов использовались даже для укрепления каркасов жилищ. Тем не менее считать главной причиной гибели животных деятельность человека нет достаточных оснований. Дело в том, что данные о характере заселения севера Евразии в позднем палеолите весьма неопределенны. Антропогенный фактор все же мог существенно дополнять влияние климата, которое по окончании ледниковой эпохи привело к широкому распространению лесов, что, естественно, и сузило сферу обитания мамонтов.
Сходные причины вызвали гибель волосатого носорога (Coelodonta antiquitatis Blum.), трупы которого сохранились в многолетней мерзлоте Сибири и алевритах Старуни. Это крупное травоядное животное с длинным густым волосяным покровом было хорошо приспособлено к условиям холодного климата (рис. 23). В вюрмскую эпоху оно заселяло открытые биотопы в перигляциальной лесостепи и степи, однако, судя по находкам в Добрудже и Северном Причерноморье, могло проникать и далее к югу. Волосатый носорог часто встречается в риссе, но особенно широко в вюрме. Самые древние радиоуглеродные датировки остатков этого животного - с р. Индигирки (СССР) около 38 тыс. и из окрестностей Глазго (Шотландия) 27,5 тыс. лет назад. По-видимому, волосатый носорог вымер немного раньше мамонта, поскольку его остатков нет на стоянках позднего мадлена.

Рис. 23. Волосатый носорог (рисунок Б. Куртена)
Большерогий олень (Megaloceros giganteus Blum.) развился из формы с небольшими рогами (М. giganteus antecedens), приуроченной к лесным и отчасти степным биотопам, обитавшей в рисс-вюрме. Более поздние популяции (М. giganteus giganteus), судя по огромным рогам, исключавшим возможность существования в лесах (рис. 24), водились в условиях открытого ландшафта. В. И. Громова отмечает обилие этих популяций в эпохи межстадиальных потеплений вюрма и обращает внимание на сравнительно редкие находки костей большеротого оленя не только на мустьерских, но и на позднепалеолитических стоянках. Распределение подвидов этого животного во времени не ясно, поскольку мало точных датировок. На основании имеющихся разрозненных определений возраста можно предполагать, что во время последнего оледенения олени подвида giganteus обитали на равнинах Европы - на юге перигляциальной лесостепи и в перигляциальной степи вплоть до Большого Кавказа. Самые молодые находки остатков большеротого оленя на Среднеевропейской равнине и в Сконе (Южная Швеция) были отнесены к аллерёду. По-видимому, окончательное исчезновение этого животного произошло в позднем дриасе (11-10 тыс. лет назад).
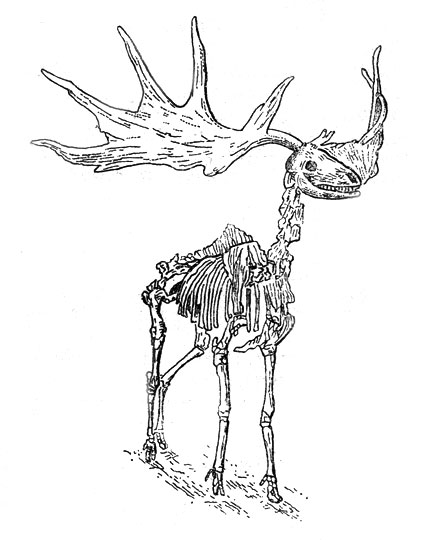
Рис. 24. Скелет большерогого оленя, найденного в межледниковом торфянике в Ирландии (реконструкция Древерманна)
Лось (Alces alces) встречался в рисс-вюрме (есть, например, в травертинах Тюрингии). Формы, близкие к современным, появляются в вюрме, когда эти обитатели открытых биотопов с широко раскинутыми рогами постепенно приспособились к жизни в лесах. Подобно современным, вюрмские лоси распространялись от приледниковой зоны до степей, но всюду в небольшом количестве. Данные по фауне пещер Добруджи показывают, что лоси обитали как во время теплых, так и холодных интервалов вюрма. Однако оптимальные условия для их распространения появились только во второй половине голоцена, когда наметилась тенденция к укрупнению размеров особей.
Северный олень (Rangifer tarandus), в настоящее время населяющий тундру, лесотундру и тайгу, в I тысячелетии до н. э. заходил в более южные районы лесной зоны (до Оки). Во время климатического оптимума эмского микулинского межледниковья (рисс-вюрм), когда распространились широколиственные леса, эти животные, по-видимому, мигрировали из средней полосы Европы на север.
Северный блень в вюрмское время был одним из самых распространенных животных Европы (рис. 25). Его остатки обнаружены на стоянках позднего мустье, ориньяка, солютре и особенно мадлена. Множество рогов найдено в позднеледниковых отложениях Средней и Восточной Европы вплоть до ее горного обрамления. Единичные местонахождения есть в Венгрии и даже на севере Испании и Италии.
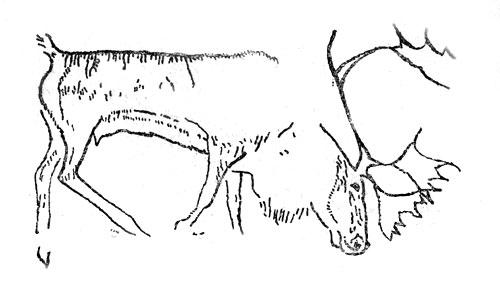
Рис. 25. Северный олень. Рисунок на стене пещеры Кесслерлох в Швей-царии (по Б. Куртену)
Овцебык, или мускусный бык (Ovibos moschatus Zimm.), развился в позднем плейстоцене из близкого вида, приспособленного к более теплым климатическим условиям. В конце последнего оледенения он встречался в перигляциальной лесостепи и на севере перигляциальной степи (Витебская и Воронежская области, Средний Урал). Причину вымирания этого вида во время деградации последнего оледенения нередко связывают с деятельностью позднепалеолитических охотников.
В Северной Америке, на севере Канадского Арктического архипелага и в Гренландии овцебык выжил в свободных ото льда убежищах, обнаружив удивительную адаптацию к суровым приледниковым условиям. Например, в Северо-Восточной Гренландии, где сохранилась довольно крупная популяция овцебыков, вегетационный период длится менее двух месяцев. Скудный и редкий растительный покров тундры приурочен главным образом к узкой прибрежной полосе, окаймленной высокими горами.
Зубр (Bison priscus) в рисс-вюрме населял леса Средней Европы наряду с лосем, бобром, туром, древним слоном и носорогом Мерка. Во время оледенений вюрма он приспособился к безлесным биотопам и наряду с мамонтом, волосатым носорогом, северным оленем и овцебыком встречался на севере перигляциальной лесостепи. Возможно, в условиях лесного ландшафта и теплого климата формировались более мелкие особи зубров, а в условиях открытых перигляциальпых местообитаний - более крупные. В позднем вюрме появляются очень крупные зубры, нашедшие отражение в произведениях позднепалеолитического искусства (рис. 26).

Рис. 26. Зубр, раненный копьями. Роспись охрой в пещере Нио во Франции (по Я. Корнуоллу)
После деградации оледенения зубр мельчает, постепенно превращаясь в форму Bison bonasus, сохранившуюся до настоящего времени в заповедниках. Датский исследователь М. Дегербёль поставил под сомнение прямую унаследованность Bison bonasus от В. priscus и выдвинул предположение, что в позднем вюрме существовали два самостоятельных вида, из которых один исчез, а другой выжил.
Первобытный бык, или тур (Bos prinaigenius), так же, как и лось, встречался в рисс-вюрме и вюрме (в теплые и холодные интервалы). В голоцене он широко распространился с эпохи неолита (рис. 27).
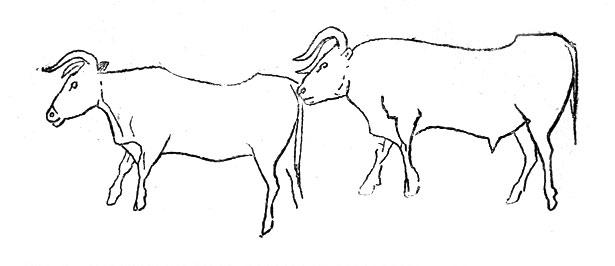
Рис. 27. Первобытные быки. Рисунок на стене грота де ла Мейри во Франции (по О. Абелю)
До сих пор неясна картина вымирания дикой лошади (Equus caballus L.). Существовало множество морфологически различных популяций, которые отражали разнообразие природных условий. На протяжении плейстоцена происходило измельчание этих животных. Самые маленькие лошади встречались во второй половине вюрма в период развития культур позднего палеолита. Одновременно существовали крупные и мелкие формы. По мнению В. И. Громовой, первые были приурочены к лесным биотопам (в качестве пастбищ использовались сырые луга и болота), вторые - преимущественно к перигляциальным биотопам.
Лошадь была важным, если не самым основным, объектом промысла позднепалеолитических охотников (рис. 28). На стоянке Солютре в Центральной Франции обнаружен слой с остатками костей не менее 100 тыс. диких лошадей. Вероятно, в этой местности стада лошадей были загнаны к крутому обрыву, с которого животные падали и разбивались насмерть. Здесь же встречаются орудия ориньякской культуры позднего палеолита.

Рис. 28. Дикая лошадь. Рисунок на стене пещеры Ла-Пасиега в Испании (по Г. Обермайеру)
Под влиянием интенсивной охоты и сведения лесов ареал дикой лошади сильно сократился. В настоящее время только в Джунгарии сохранилась небольшая популяция лошади Пржевальского (Equus przewalskii), находящаяся на грани вымирания. Исторические документы свидетельствуют о существовании в средние века в Западной Европе дикой лошади - тарпана (Equus gmelini). В Польше ее встречали еще в XVIII в., а на Украине - в первой половине XIX в. До сих пор окончательно не установлено, был ли тарпан результатом естественного развития Equus caballus или просто результатом одичания группы домашних лошадей.
Осел (Asinus hydruntinus Reg.) появился только в позднем плейстоцене, начиная с рисс-вюрма. Остатки его часто находят на стоянках позднего палеолита на территории всей Средней Европы, в том числе и вместе с холодостойкой фауной. В неолите, а на востоке Европы даже в мезолите увеличивается число местонахождений остатков осла, который, по-видимому, был одним из первых домашних животных.
Остатки пещерного медведя (Ursus spelaeus) очень часто встречаются в пещерах (рис. 29), однако преимущественно в виде отдельных костей. В 1979 г. польским спелеологам удалось обнаружить в одной из пещер Татрских гор прекрасно сохранившийся скелет этого животного, что позволит точно установить его размеры и вес. Пещерный медведь появился в Европе еще в миндельское время, пережил несколько оледенений и межледниковий и вымер в конце вюрма. Установлено, что одновременно существовал и бурый медведь (Ursus arctos), однако оба вида редко встречались вместе. Пещерный медведь отличался строением скелета и зубов; по всей вероятности, он в основном был фитофагом.

Рис. 29. Пещерный медведь (рисунок В. Куртена)
Предположение о ведущей роли первобытного человека в истреблении пещерного медведя окончательно не доказано, хотя остатки этого животного встречаются даже на мустьерских стоянках в горах Центральной Европы. Вымирание животного происходило постепенно, путем оттеснения небольших популяций в изолированные горные районы. Это ухудшало условия репродукции и приводило к уменьшению размеров особей. В конечном итоге пещерный медведь полностью исчез.
Польский ученый В. Ковальский обратил внимание на тот факт, что находки костей животного в пещерах можно объяснить лишь их лучшей сохранностью. В настоящее время сходная ситуация наблюдается в пещерах центральной части Татр, содержащих много костей бурого медведя, хотя эти животные устраивают свои берлоги не в пещерах, а под выкорчеванными деревьями. Однако вне пещер костные остатки плохо сохраняются вследствие резкого преобладания различных эрозионных процессов.
Из-за крайне неравномерного размещения пещер трудно представить, что какой-либо вид крупных млекопитающих был тесно с ними связан. В позднем палеолите заселение пещер человеком происходило весьма нерегулярно, за исключением, может быть, отдельных районов Франции и Испании, тем более в условиях высокогорий. Таким образом, конкурентная борьба с человеком за кров явно не могла быть причиной вымирания пещерного медведя. Скорее всего, здесь сыграло роль сокращение подходящих биотопов.
Пещерный лев (Felis leo spelaea), пещерная гиена (Crocuta crocuta spelaea) и саблезубый кот (Homotherium latidens), по-видимому, не были важными объектами промысла позднепалеолитических охотников (рис. 30). Все три вида вымерли в конце вюрма.
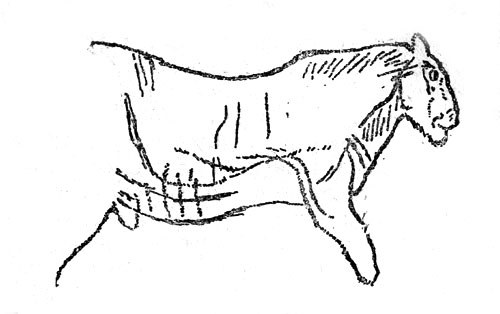
Рис. 30. Пещерный лев. Рисунок на стене пещеры Ле Комбарель во Франции (по Брёйлю)
Подводя итоги, надо отметить, что в Европе большое число видов крупных млекопитающих смогло пережить позднеплейстоценовые оледенения и сохранилось в составе современной фауны. К этой группе относятся лось, благородный олень, косуля, кабан, волк, бурый медведь и др. Зубр находился на грани истребления в историческое время и уцелел только в заповедниках, тогда как первобытный бык и лошадь полностью исчезли в естественном состоянии, но сохранились в составе поголовья домашнего скота.
Влияние изменений климата и структуры биотопов активно отразилось на вымирании большеротого оленя, пещерного медведя, пещерного льва и пещерной гиены. С теми же факторами иногда связывают гибель таких видов животных, как мамонт, волосатый носорог, хотя в данном случае невозможно исключить активное воздействие антропогенного фактора: эти крупные звери, несомненно, играли немалую роль в обеспечении ресурсами мяса возросшего позднепалеолитического населения Европы. Причиной гибели животных могло быть и отсутствие адаптации к глубокому снежному покрову, появившемуся в эпоху последнего оледенения. Однако установлено, что мамонт и волосатый носорог были широко распространены именно во время этого оледенения и вымерли лишь на последних стадиях его деградации. Следовательно, вряд ли можно говорить о повышенной снежности в то время.
Сложность решения проблемы усугубляется из-за недостаточной точности реконструкций природных обстановок прошлого. Как известно, оценки климата в основном сводятся к установлению средних показателей температур и осадков, тогда как на гибель животных могли повлиять в первую очередь сдвиги в экстремальных, а не средних показателях. Позднеплейстоценовое вымирание фауны производит особо сильное впечатление, поскольку речь идет о наиболее молодом этапе истории животного мира (о предыдущих этапах мы знаем гораздо меньше). Окончательному вымиранию должно было предшествовать прогрессировавшее уменьшение размеров особей и численности популяций. Для того чтобы проследить ход этого процесса, необходимо тщательно изучить костные остатки, определить их возраст.
Наконец, следует заметить, что анализ остатков животных из ныне известных стоянок не всегда полно отражает состав фауны в окружающих районах - помимо разной степени сохранности могли сказаться сезонность охотничьего промысла и специализация, характерная для первобытного населения данного района. Отдельные группы первобытных людей выбирали объектом своей охоты определенные виды животных: мамонтов, северных оленей, диких ослов, зубров. Эта специализация, достаточно ясно прослеживаемая и на многослойных стоянках, сильно влияет на интерпретацию современными исследователями картины развития животного мира.
Б. Куртен приводит интересные материалы по пещере Фогельхерд в Вюртемберге (ФРГ), где выявлена четкая последовательность культурных горизонтов палеолита. В раннем вюрме неандертальцы, представители мустьерской культуры, специализировались главным образом на добыче дикой лошади и в меньшей степени - мамонта. В конце среднего вюрма появился кроманьонский человек, носитель ориньякской культуры. Охота на лошадей тогда продолжалась, однако возросло значение мамонта и в меньшей степени - северного оленя. В позднем вюрме, с которым совпало развитие мадленской культуры, северный олень и дикая лошадь выдвинулись в промысле на первое место, а значение мамонта резко сократилось.
Эти весьма показательные данные вряд ли можно использовать для целей точной характеристики динамики животного мира в вюрме, поскольку трудно исключить фактор специализации местных охотников. Гораздо более надежные результаты дают привлечение массовых данных по группам стоянок и их сравнительная оценка. Кроме того, необходимо учитывать сведения о находках остатков животных, не связанных с культурными горизонтами. Только таким путем можно составить наиболее объективное представление о временной дифференциации териофаун.
Как известно, эволюция видов, их выживание и исчезновение определяются долговременными непрекращающимися изменениями в окружающей среде. За время существования жизни на Земле непрерывно происходила гибель организмов, которые не смогли приспособиться к изменившимся условиям. Перестройка природной среды в конце плейстоцена характеризовала один из этапов этой сложной истории развития и, вполне естественно, могла привести к вымиранию каких-то групп животных. Следует, однако, заметить, что благодаря высокой мобильности животные отличаются повышенной способностью к выживанию. Во всяком случае, именно таким путем можно объяснить отсутствие массового исчезновения родов животных в периоды деградации более древних плейстоценовых оледенений.
В конце последнего оледенения под влиянием ряда факторов естественного отбора многие животные оказались в стадии предельного равновесия со средой. Но тогда возросло, значение кратковременных внезапных изменений, способных вызвать коренную перестройку цепи взаимосвязанных экосистем. Такие трансформации были внесены в связи с быстрым развитием охотничьих культур позднего палеолита, и совместное действие природных и антропогенных факторов оказало глубокое влияние на животный мир.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'