
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Отико-окципитальный отдел
Передне-, заднеушные и боковые затылочные кости полностью срастаются, лишь у пинакозавра (экз. N 4046) из Бага-Тариача виден вертикальный шов проотики и парокципитального отростка приблизительно на уровне лагены (рис. 8). Передняя часть проотики срастается с латеросфеноидами, идущими к крыше черепа, и с вентральным выступом теменных костей - медиальным продолжением посторбитального шельфа.
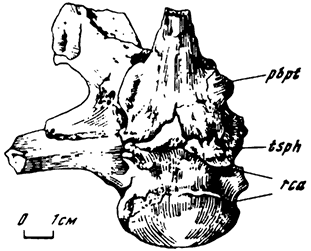
Рис. 8. Мозговая коробка анкилозаврида Pinacosaurus grangery Gilmore (снизу). Экз. ПИН. N 4043: Бага-Тараич; верхний мел, джадохтская свита. Обозначения см. на рис. 7
Внутреннее ухо (см. рис. 7) открыто со стороны мозговой полости, так как его медиальная стенка не окостеневала. Лагены крупные. Над ними в боковой стенке мозговой коробки расположено преддверие. Ампулы полукружных каналов четко обозначены: задняя в виде сильного расширения нижней части заднего и конца горизонтального полукружных каналов, передняя и горизонтальная - сближены; последняя расположена несколько ниже и медиальнее передней.
Передний полукружный канал проходит в толще передней стенки капсулы внутреннего уха и выходит верхней частью на ее внутреннюю поверхность для соединения с задним каналом. Здесь в стенке эндокрания имеется углубление, направленное назад и вниз и заканчивающееся слепо. Возможно, эта структура вмещала утолщенную верхнюю часть crus communis, который шел в медиальной, не окостеневающей стенке. Но не исключено, что оно не имело связи с перепончатым лабиринтом (Курзанов, Туманова, 1978), в этом случае оно может быть сравнимо с recessus interacusticus хищного динозавра итемируса (Курзанов, 1976). Над полостью внутреннего уха, в верхнем крае стенки эндокрания, проходит борозда, по-видимому, эндолимфатического протока, разделяющаяся на две веточки: задняя далеко не прослеживается, а передняя входила в полость внутреннего уха на уровне верхнего края горизонтальной капсулы. За пределами полости эндокрания борозда прослеживается до верхнего края парокципитального отростка, где эндолимфатический проток мог выходить наружу.
Затылок анкилозаврид широкий (см. рис. 3, г; 4, г). Кости прочно срастались между собой. Посттемпоральное окно в виде щели имеется у пинакозавра, но полностью зарастает у остальных анкилозаврид. Большое затылочное отверстие крупное, составляющее свыше 1/3 высоты затылка, округло-ромбовидное, несколько удлиненное в горизонтальном направлении у пинакозавра, сайхании и амтозавра (рис. 9, а) и вытянутое вертикально у таларуруса и тархии (см. рис. 3, г, 5, г). Парокципитальные отростки, по высоте в два раза уступающие затылку, тянутся перпендикулярно сагиттальной оси черепа (см. рис. 3, б), у нодозаврид - под некоторым углом (см. рис. 1, а), у сайхании они очень низкие, с загибающимся вниз нижним краем. Парокципитальные отростки у Tarchia (см. рис. 3, г), приблизительно одинаковой высоты по всей длине, но у Talarurus (см. рис. 5, г) и особенно у Shamosaurus (см. рис. 4, г) в дистальной части они шире, а в проксимальной вдаются вперед, в результате чего остеодермальный край крыши черепа несколько нависает над медиальной частью затылка. У таларуруса (см. рис. 5, г) концы парокципитальных отростков крючкообразно загнуты вниз, у большинства анкилозавров - лишь незначительно опущены.
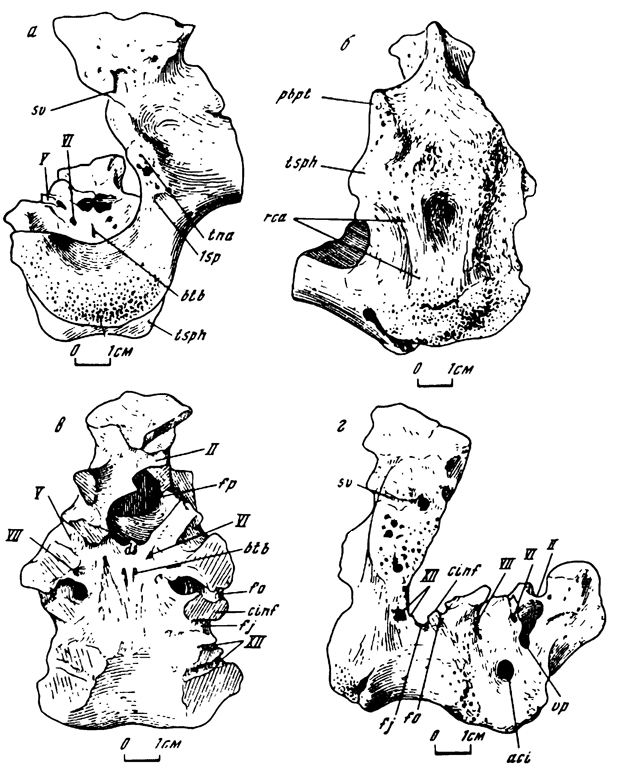
Рис. 9. Мозговая коробка Анкилозавра Amtosaurus magnus Kursanov et Tumanova. Голотип - ПИН № 3780/2; Амтгай; верхний мел; баинширэинская свита. а - сзади, б - снизу, в - сверху, г - сбоку. Обозначения см. на рис. 7
У таларуруса (экз. 3780/1) благодаря неполному срастанию затылка с крышей черепа на нижней поверхности крыши черепа, позади ямки для верхнего отростка квадратных костей, видна поперечная бороздка, вмещающая верхний край парокципитальных отростков. "Tabulare", обозначенное на черепе Pinacosaurus grangeri (Maryanska, 1971, 1977), является элементом нависающего заднего края крыши черепа, составленного остеодермами.
Верхняя затылочная кость подходит к foramen magnum лишь в самой его вершине. У всех анкилозавров, кроме молодого экземпляра пинакозавра, supraoccipitale прочно срастается с крышей черепа (см. рис. 3, г; 4, г).
Боковые затылочные кости полностью срастаются с заднеушными костями. На границе с большим затылочным отверстием на Exoccipitalia наблюдаются парные бугорки, по-видимому, для невральных дуг первого шейного позвонка, а в основании бугорков - бороздки, в которых могла проходить затылочная вена.
Основная затылочная кость выстилает заднюю часть дна мозговой полости и ограничивает затылочное отверстие снизу, образуя затылочный мыщелок. Затылочный мыщелок - от узкоовального у таларуруса и амтозавра (см. рис. 7, в) до широкоовального у сайхании, тархии (см. рис. 3, б) и округлого у пинакозавра, малеевуса и шамозавра (рис. 10, а; 4, б). Суставная поверхность направлена более или менее постеровентрально. На дорсальной стороне основной затылочной кости неглубокая ямка.
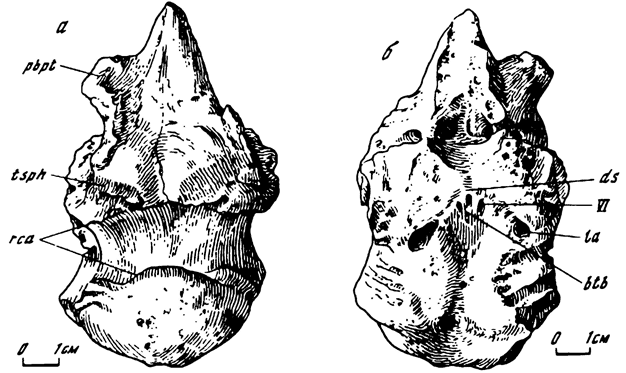
Рис. 10. Мозговая коробка анкилозаврида Maleevus disparoserratus (Maleev). Экз. ПИН, N 554/2-1; Ширэгин-Гашун; верхний мел, баинширэинская свита. а - снизу; б - сверху. Обозначения см. на рис. 7
На вентральной поверхности основной затылочной кости, между мыщелком и сфенокципитальными буграми, находится поверхность прикрепления m. rectus capiti anteriores. Рельеф этой области сильно разнится: у шамозавра здесь сравнительно узкая, выпуклая вентрально, округлая поверхность; у таларуруса срединное положение занимает пологий гребень, латерально от которого тянутся депрессии; у амтозавра рельеф обратный: в центре - углубление, по бокам - пологие гребни, направленные спереди назад. У пинакозавра поверхность здесь сильно сглажена, вогнута, с резким опусканием к медиальной части сфенокципитальных бугров; у малеевуса вентральная поверхность основной затылочной кости также сглаженная, антеролатерально расширяющаяся, медиально еще более резко опускающаяся к сфенокципитальным буграм; у сайхании и тархии этот регион короткий и выровненный. На границе основной затылочной кости с базисфеноидом, между сфенокципитальными буграми, неизменное углубление, достигающее у сайхании и тархии значительных размеров (см. рис. 3, б).
Эндокраниальная полость. Самое конечное положение в боковой стенке мозговой коробки занимали отверстия для двух веточек XII нерва, на границе exoccipitale и opisthotica - крупное яремное отверстие, через которое выходили IX, X и XI черепные нервы. Немного вперед, отделяясь crista interfenestralis, - овальное окно. У амтозавра (см. рис. 9, в, г) crista interfenestralis - толстая, сплошная, у таларуруса (рис. 10, а, е) - неполная, у тархии - яремное отверстие и овальное окно полностью сливаются.
Эндокраниальная полость небольшого объема. Ее высота увеличивается вперед. Дно ровное, прямое, лишь слегка поднимающееся у амтозавра в передней половине полости, перед гипофизарной ямкой. Угол, составленный плоскостями парабазисфеноидного комплекса и основной затылочной кости колеблется между 120° у амтозавра (см. рис. 9, г) и 150° у таларуруса (см. рис. 7, а).
Вдоль передней половины дна мозговой полости тянется гребень, сравнимый с basis tuberculi basalis черепах (Gaffney, 1972), заканчивающийся у амтозавра на вершине спинки турецкого седла (см. рис.9, в) треугольным выростом. У основания задней стенки спинки, по бокам от basis tuberculi basalis, - углубления (у амтозавра значительные) для прикрепления тяжей, поддерживавших продолговатый мозг. Спинка турецкого седла хорошо выражена. Ее передняя стенка наклонена вперед, составляя острый угол с задней поверхностью sella turcica, затем, примерно на уровне половины высоты (у пинакозавра и малеевуса несколько ниже) она становится почти вертикальной. Через спинку турецкого седла вперед и в сторону проходит канал VI нерва. Над ним - небольшое выходное отверстие глазной ветви тройничного нерва, несколько сзади - отверстие для двух челюстных ветвей n. trigeminus (V2,3). Несколько назад и выше - более мелкий канал VII нерва. По форме наружного отверстия можно судить о направлении ветвей лицевого нерва: гиомандибулярной, шедшей наружу и назад, и нёбной - вперед и вниз по желобку к нёбной ветви внутренней сонной артерии. Соединение происходило на дорсальной поверхности базиптеригоидного отростка - парабазальный канал отсутствует. От входного отверстия в канал VII нерва по внутренней поверхности эндокрания тянется желобок VIII нерва, направленный в костный лабиринт внутреннего уха. Над следом от соединенных нёбных ветвей сонной артерии и лицевого нерва находится вертикально-овальное крупное входное отверстие церебральной ветви внутренней сонной артерии. Вперед и выше относительно него расположено отверстие примерно равного размера в форме неправильного овала с наклоненной назад осью. В этом месте из мозговой полости выходила, по всей вероятности, питуитарная вена. Антеролатеральную стенку гипофизарной ямки пронизывают каналы меньших размеров, направленные наружу вверх, предназначенные, по-видимому, для a. ophtalmica, имеющей в этом случае отдельный несколько заниженный выход. Впереди гипофизарной ямки поперек проходит крупная борозда зрительного нерва.
Указания о большой морфологической отдаленности анкилозавров и других орнитисхий и предположение о раннем отделении их от предкового ствола псевдозухий уже имелись (Coombs, 1978а). Основанием для этого служила морфология черепа, а также детали строения посткраниального скелета. Изучение строения эндокрания подтверждает это предположение. При сравнении морфологических схем строения мозговых коробок динозавров (Osborn, 1912; Brown, and Schlaikjer, 1940; Ostrom, 1961; Russel, 1969; Курзанов, 1976) и приведенного выше плана, характерного для монгольских анкилозавров, становится очевидно, что по ряду признаков мозговая коробка анкилозавров отлична от таковой других динозавров. Отчасти специфика строения мозговой коробки анкилозавров связана с полностью "выпрямленным" ее основанием.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'