
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Семейство Rhagionidae latreille, 1802
Богато представлено в поздней юре Каратау, где является крупнейшим семейством азиломорф как по общей численности, так и по систематическому разнообразию и одним из субдоминантов среди семейств наземных двукрылых. Подавляющая часть материалов из Каратау остается необработанной. Семейство известно также из местонахождения Бахар в Монголии (предположительно - средняя юра). Можно предположить, что систематическое разнообразие рагионид в юре было выше, чем в настоящее время (Ковалев, 1984).
В сибирской юре рагиониды распространены шире, чем какое-либо другое семейство бибиономорф или азиломорф. Они отмечены в ранней - средней юре Забайкалья, Иркутского и Кузнецкого бассейнов, в средней юре Чулымо-Енисейской впадины и в средней - поздней юре Забайкалья, однако везде представлены немногими остатками, и можно предположить, что общая численность их была намного ниже, чем в Каратау. В целом в материалах из Сибири они составляют 10% от общего числа наземных двукрылых, но в ряде местонахождений оказываются единственными наземными (Могзон-Вишневый, Борже - скв. 35, глубина 154 - 156 м) или вообще единственными (Лягушье, Ия) представителями отряда.
Меньшим, чем в Каратау, очевидно, было и систематическое разнообразие. Почти весь материал был описан ранее (Ковалев, 1981б, 1982). В общей сложности он насчитывает 10 видов 6 родов из Забайкалья, Иркутского и Кузнецкого бассейнов. Единственный представитель из Чулымо-Енисейской впадины описывается ниже.
Все рагиониды рассматриваемых фаун принадлежат к родам, известным только из сибирской юры, но из этого преждевременно делать вывод о высоком уровне эндемизма юрской фауны Сибирской области, поскольку изучение индоевропейских рагионид, прежде всего каратауских, только началось.
Каждый вид рагионид юры Сибири известен из единственного местонахождения по одному-двум экземплярам.
После недавней ревизии системы семейства (Nagatomi, 1982) появилась возможность уточнить положение юрских родов. Роды Palaeobrachyceron V. Kovalev и Jurabrachyceron V. Kovalev, скорее всего, принадлежат к подсемейству Glutopinae. Менее вероятна их принадлежность к Rhagioninae. Основной признак, используемый для различения этих подсемейств, - строение яйцеклада - не удалось изучить ни на одном из остатков. Известный только отпечатку крыла род Kubekovia gen. nov. по жилкованию ближе всего к Glutopinae, но сколько-нибудь надежное указание подсемейственной принадлежности рагионид по жилкованию невозможно. Род Palaeoptiolina V. Kovalev относится к Spaniinae, a Palaeobolbomyia V. Kovalev - к Rhagioninae; систематическая принадлежность этих родов достаточно надежно устанавливается по строению антенн и (для Palaeobolbomyia) яйцеклада. Род Ussatchovia V. Kovalev не принадлежит ни к одному из современных подсемейств и близок к роду Rhagionempis Rohdendorf из. поздней юры Каратау. Этот последний род первоначально был описан в составе рагионид как представитель особого подсемейства (Родендорф, 1938), а позднее был выделен в особое семейство Rhagionempi- didae (Родендорф, 1962), синонимизированное нами с Rhagionidae (Ковалев, 1982). В настоящее время представляется целесообразным восстановить его как подсемейство Rhagionempidinae в составе рагионид; его уточненный формальный диагноз дан ниже. Положение рода Ija V. Kovalev, известного по фрагменту крыла, остается неопределенным; очевидно, этот наиболее крупный представитель юрских двукрылых Сибири (размером с небольшого слепня) не близок ни к одному из остальных юрских родов. Подробнее данные об эволюции рагионид в юре и гипотеза о ключевом положении этого семейства в филогении азиломорф изложены ранее (Ковалев, 1982).
Личинки юрских рагионид были, вероятно, хищными и обитали в почве, скоплениях разлагающихся растительных остатков или в слоевищах печеночников. Низкая численность Rhagionidae в ориктоценозах может объясняться либо их реальной низкой численностью, либо обитанием вдали от воды.
Подсемейство Rhagionempidinae rohdendorf, 1938, stat. restaur.
Rhagionempidinae: Родендорф, 1938, с. 33; Rhagionempididae: Родендорф, 1962, с. 336.
Диагноз. Мелкие рагиониды с крылом до 4 мм. Голова полусферическая, сзади плотно прижата к груди. Глаза самки на лбу широко разделены. Антенны короче головы, с цельным коническим симметричным 3-м члеником, завершенным нечленистой стилевидной толстой аристой, которая короче членика. R2+3 впадаете костальный край далеко от R1; отрезок костального края между R1 и R2+3 гораздо длиннее, чем между Sc и R1. Вилка R4 и R5 явно короче ствола R2+3; жилки R4 и R5 явственно расходящиеся. Три или четыре ветви М. Яйцеклад образован только VIII и последующими сегментами брюшка; VIII сегмент отделен от VII более длинной мембраной, чем между предыдущими сегментами.
Состав: роды Rhagionempis Rohdendorf, Probolbomyia Ussatchov и Ussatchovia V. Kovalev.
Сравнение. От подсемейства Spaniinae, с которым сближается по строению антенн, отличается укороченной вилкой R4 и R5 с сильно расходящимися ветвями и удлиненной мембраной между VII и VIII сегментами.
Распространение. Средняя - верхняя юра Забайкалья, верхняя юра Казахстана.
Подсемейство ?
Род Kubekovia V. Kovalev, gen. nov.
Название рода от села Кубеково.
Типовой вид - К. accessoria sp. nov.; средняя юра Чулымо-Енисейской впадины.
Диагноз. Мухи небольшого размера. Крыло широкое. Жилкование четкое. С сильная по переднему краю крыла, тонкая по заднему. Sc оканчивается у середины крыла. R1 слабо и равномерно изогнута. Отрезок костального края между Sc и R1 короче, чем между R1 и R2+3. RS длинный, примерно равен стволу R4+5, начинается намного проксимальнее основания d. R2+3 и R4 не изгибаются вперед. Вершинные отрезки R4 и R5 длинные, длиннее ствола R4+5; R4 достигает края крыла перед его вершиной, R5 - на вершине или чуть позади нее. R5 выглядит непосредственным продолжением R4+5, а R4 - ее отростком, отходящим под значительным углом, но сразу дуговидно изгибающимся и далее параллельным R5. bа гораздо длиннее и уже, чем bp, проксимальнее первого развилка RS разделена косой поперечной сверх комплексной жилкой, составляющей как бы продолжение основного края d. rm на уровне окончания Sc, перед серединой d; последняя находится в центре крыла, с почти параллельными передним и задним краями, сильно удлиненная. Четыре полные ветви М. M1, М2 и М3 отходят от вершины d; основания M1 и М3 равно удалены от М2. Вершинный отрезок М3 не длиннее заднего края d между точками отхождения от нее М3 и М4, параллелен М4. mcu отделена от d короткой tb. Вершинный отрезок СuА ДЛИННЫЙ, длиннее половины основного, а замкнутая на крае крыла.
Состав. Типовой вид.
Сравнение. Отличается от всех прочих рагионид наличием внутри bа сверх комплексной поперечной жилки. Схема жилкования наиболее близка к Palaeobrachyceron V. Kovalev и Glutops Burgess, но у первого вершинный отрезок М3 длиннее заднего края dt у второго а открытая.
Название вида от accessorius (лат.) - добавочный.
Голотип - ПИН, N 1255/269, позитивный отпечаток крыла, анальная лопасть утрачена; Кубеково, итатская свита.
Описание (рис. 120). Крыло в 2,3 раза длиннее своей ширины, затемненное, с большим еще более темным стигмальным пятном, занимающим большую часть r1 и вершину sc; мембрана в пределах sc до стигмального пятна и в основании r1 до развилка RS светлая. Жилки темные, ствол М между bа и bp не окрашен. R2+3 образует слабый волнообразный изгиб. Отрезок костального края между R1 и R2+3 в 1,7 раза длиннее отрезка между Sc и R1. M1 параллельна RS. Отрезки края крыла между М2 и М3 и между М3 и М4 равны, расстояние между вершинами M1 и М2 в 1,3 раза короче. A1 прямая.
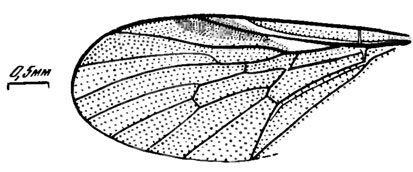
Рис. 120. Сем. Rhagionidac. Kubekovia accessoria sp. nov., голотип, крыло
Размеры, мм: длина крыла - 4,2.
Материал. Голотип.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'