
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Подкласс Foraminifera. Фораминиферы
Общая характеристика. К подклассу фораминифер (лат. foramen, род. foraminis - отверстие, дыра, fero - носить) относится большая группа саркодовых, насчитывающих до 20 000 современных и ископаемых видов, цитоплазма которых заключена в органическую, агглютинированную или известковую раковину. Псевдоподии фораминифер состоят из тонких, разветвленных, корневидных, соединяющихся между собой (анастомозирующих) нитей, выходящих из раковины либо только через устье, либо через устье и каналы, пронизывающие стенку раковины. Фораминиферы в большинстве своем морские бентосные или планктонные, свободноживущие или прикрепленные формы. Небольшая часть фораминифер приспособилась к жизни в солоноватоводных бассейнах и лишь немногие известны в пресных водоемах. В ископаемом состоянии известны начиная с кембрия.
Строение тела. Цитоплазма фораминифер обычно бесцветная, иногда окрашена в розовый, оранжевый или желтый цвета. Эктоплазма, довольно однородная по структуре, осуществляет обмен веществ с внешней средой и служит местом образования псевдоподий. Под электронным микроскопом псевдоподии представляют собой пучок волоконец разного диаметра; каждое волоконце окружено оболочкой. Способность псевдоподий вытягиваться и втягиваться основана на свойстве цитоплазмы изменять свое агрегатное состояние, переходя из жидкого состояния (золь) в вязкое (гель). Псевдоподии, не связанные с субстратом, разветвляются, соединяются перемычками и образуют своеобразную ловчую сеть, в которую попадают личинки, различные микроорганизмы и органический детрит (рис. 26). Переваривание пищи нередко происходит вне раковины.
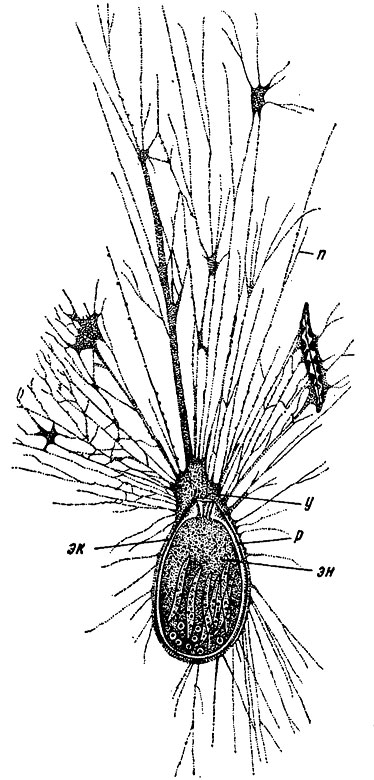
Рис. 26. Отряд Allogromiida. Схема строения Allogromia ovoidea Rhumb, (совр.): п - псевдоподии, р - раковина, у - устье, эк - эктоплазма, эн - эндоплазма
Эндоплазма заполняет внутреннюю полость раковины и отличается от эктоплазмы более темной окраской и менее однородной структурой. Эндоплазма содержит ядро, многочисленные ультрамикроскопические структуры и различные включения, к числу которых относятся поступившие извне: пища, минеральные частицы, паразитические микроорганизмы, а также продукты жизнедеятельности клетки. В эндоплазме многих фораминифер в симбиозе с ними живут микроскопические водоросли - зооксантеллы, которые в процессе фотосинтеза поглощают углекислый газ, вырабатываемый фораминиферами при дыхании, и выделяют кислород, необходимый последним для газообмена. Перед размножением эндоплазма претерпевает глубокие изменения, она очищается от посторонних частиц и просветляется. При этом у многих фораминифер образуются временные оболочки - цисты, состоящие из тектиновой основы, посторонних частиц и продуктов выделения, эндоплазмы.
Строение раковины. Подавляющее число фораминифер имеет раковину, и лишь у небольшой части цитоплазма окружена утолщенной эластичной органической оболочкой - мембраной. Раковина может быть относительно простой или достигать большой сложности (рис. 27). Ее размеры колеблются от 0,02 до 110-120 мм. Стенка раковины может быть органической, агглютинированной и известковой. Наиболее низкоорганизованные фораминиферы (аллогромииды) имеют стенку, состоящую из тектина, представляющего комбинацию протеинов и углеводов. У многих фораминифер в тектиновую стенку включены посторонние частицы разного минерального и химического состава: зерна кварца, различных тяжелых минералов, карбонатов, пластинки слюды, спикулы губок, органический детрит (обломки спикул губок, раковинки иных фораминифер, скелеты радиолярий, обломки раковин моллюсков) и другой "строительный материал".

Рис. 27. Типы строения раковин фораминифер и предполагаемые пути их изменения (Покорный, 1958)
При этом фораминиферы обычно так же, как и раковинные амебы, "заглатывают" во внутрь этот "строительный материал". Через некоторое время протоплазма разбухает и "строительный материал" выталкивается на поверхность, где цементируется тектином, карбонатом кальция, окислами или карбонатом железа.
Таким образом, возникают агглютинированные раковины.
Раньше предполагалось, что в редких случаях цементом у некоторых фораминифер мог быть кремнезем. Однако у современных фораминифер наличие кремневого цемента пока не установлено. Многие исследователи считают, что наблюдаемый у ряда ископаемых фораминифер кремневый скелет является вторичным и развился в процессе фоссилизации по карбонату кальция. Неясным остается также вопрос, откуда берется железистый цемент, обладают ли фораминиферы способностью выделять железо из цитоплазмы или оно привносится извне в виде обломков железистых минералов. Цитоплазма некоторых фораминифер обладает своеобразной избирательной способностью - для постройки скелета она "выбирает" материал только определенного размера, цвета и даже состава, например только зерна кварца или спикулы кремневых губок, или листочки слюды. Но чаще всего используется всякий подходящий обломочный материал, рассеянный на дне водоема. Цемент и агглютинированные частицы входят в состав раковины в разной пропорции: у одних форм частицы плотно прилегают друг к другу, у других они разделены участками цемента, иногда цемент полностью преобладает. Микроструктура стенки агглютинирующих фораминифер изучена недостаточно. У многих имеется внутренняя органическая выстилка.
Большинство фораминифер имеет секреционную известковую раковину, стенка которой состоит из тектиновой основы, пропитанной минеральными солями; важную роль здесь играет углекислый кальций (кальцит или арагонит) с различным количеством примеси углекислого магния (до 18%) и фосфата кальция и магния. Стенка известковых раковин по своему строению довольно разнообразна. Существует три основных типа микроструктур стенки: микрогранулярный, фарфоровидный и гиалиновый (стекловидный). В последнее время выделен еще криптокристаллический. Употребляемые названия "фарфоровидный" и "стекловидный" не очень подходящие, так как они отражают не специфику самой микроструктуры, а общий облик стенки, но эти названия общеприняты и пока существуют в литературе.
Микрогранулярный тип стенки наблюдается у палеозойских эндотирид, фузулинид и у некоторых мезокайнозойских отрядов; он характеризуется наличием зерен микрозернистого кальцита размером от 1 до 5 мк, отсутствием цемента и непостоянной примесью агглютинированных частиц. Раковина с таким типом микроструктуры стенки не имеет скульптуры, дополнительных скелетных образований; внутренний скелет представлен в виде выростов стенки. Поверхность раковины тусклая, светлого или серовато-желтого цвета.
Фарфоровидный тип стенки отличается беспорядочным расположением кристаллов и их кристаллографических осей; кристаллы имеют различную форму, их размеры от 0,5 до 5 мк. В отраженном свете стенка белая, фарфоровидная, иногда блестящая. Стенка раковины содержит органическую основу. Этот тип стенки характерен для отряда милиолид.
Стекловидный, или гиалиновый, тип разделяется на два подтипа: стекловато-зернистый и стекловато-радиальный. У первого подтипа кристаллы кальцита или арагонита однообразной округлой или угловатой формы, плотно прилегают друг к другу; размеры кристаллов 0,5-10 мк; оптические оси ориентированы беспорядочно или с преобладанием определенной ориентировки с осью С под углом к поверхности стенки. У стекловато-радиального подтипа кристаллы кальцита или арагонита сильно удлиненные, расположены в основном перпендикулярно к поверхности стенки; так же расположена оптическая ось С.
Криптокристаллический тип микроструктуры стенки характерен для палеозойских фораминифер; стенка состоит из кристаллов кальцита с нечеткими границами.
Нередко в процессе фоссилизации секреционных известковых раковин возникают вторичные микроструктуры, связанные с процессами перекристаллизации. В одних случаях происходит укрупнение кристаллов, в других - распадение удлиненных кристаллов на мелкие субизометрические зерна.
Макроструктуру стенки раковины образуют морфологически обособленные слои, внутрикамерные выстилки, вторичные слои на внешней поверхности раковины и на поверхности септ.
Первичная стенка раковины может быть однослойной либо состоящей из двух или нескольких слоев. Первично-однослойные стенки развиты преимущественно у представителей с фарфоровидной микроструктурой, а также у многих агглютинированных и тектиновых раковин. У фораминифер со стекловидной и с микрогранулярной структурой развиты как однослойные, так и многослойные стенки; у многослойной стенки отдельные слои разделены тонкими прослоями органического вещества; слои, слагающие стенки, обычно отличаются друг от друга особенностями строения. Для некоторых групп (фузулиниды) эти слои имеют специальные названия: первичная стенка называется протекой; она состоит из наружного тонкого слоя - тектума и основного внутреннего слоя, носящего, различные названия. У швагерин он имеет ячеистое строение и назван кериотекой (см. рис. 39). У стекловатых многослойных раковин трехслойную первичную стенку предложено называть биламеллярной, поскольку первично в ней различали внутренний и наружный (или основной) слои.
Стенка раковины изнутри бывает выстлана тонкой органической пленкой. На внешней поверхности раковины и на внутренних оборотах развиты вторичные слои стенок раковины; они образуются после формирования новой камеры в виде последующих наслоений на наружной или внутренней стороне ранее образовавшейся стенки (их называют иногда слоями нарастания, или утолщения, или вторично-многослойными).
В простейшем случае при образовании новой камеры вся открытая часть раковины перекрывается новым раковинным веществом и ее старая часть значительно утолщается (рис. 28), в то время как вновь образованная септа и все предшествующие септы остаются однослойными (рис. 28, 1); такой тип строения наблюдается у нодозариид, булиминид и простейших семейств роталиид. Во втором случае при образовании новой камеры раковинное вещество перекрывает всю открытую часть раковины и налегает на предшествующую септу таким образом, что она становится двойной, а вновь образованная апертурная септа остается однослойной (рис. 28, 3). У подобных двойных септ в полостях, остающихся между двумя слоями, развивается система септальных каналов. Такой тип двойных септ с системой внутрисептальных каналов характерен для отряда роталиид и получил название роталоидных септ. В третьем случае вновь образованная камера с конечной апертурной септой является первично-двойной и по способу образования напоминает первый случай (рис. 28, 2). Подобные двойные септы, также снабженные системой каналов, характерны для раковин некоторых групп отрядов булиминид и нуммулитид (орбитоиды).
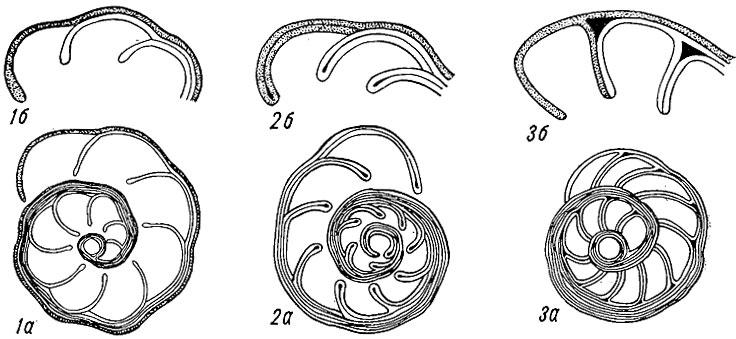
Рис. 28. Типы строения стенки: 1 - моноламеллярный, стенка первично-однослойная; 2 - биламеллярный, стенка первично-двуслойная; 3 - роталоидный, стенка первично-однослойная (зачернено внутрисептальное пространство); а - общий вид, б - три последние камеры
Пористость стенки. У многих фораминифер наблюдается пористая стенка. Поры могут быть простыми и сложными. Простые поры представлены цилиндрическими канальцами диаметром 0,2-0,5 мк; сложные поры характеризуются объединением мелких поровых канальцев в более крупные (кериотекальная пористость у фузулинид).
У некоторых мезокайнозойских фораминифер наблюдается альвеолярное строение стенки, образованной различными выростами, составляющими дополнительные внутрикамерные скелетные образования. Все поровые каналы обычно покрыты органической выстилкой. Форма и частота пор на раковине в последние годы интенсивно изучаются с применением электронного сканирующего микроскопа.
Форма раковины. Раковина фораминифер может быть одно-, двух- и многокамерной (рис. 29). При непрерывном росте образуется раковина, не разделенная на камеры; такая раковина называется однокамерной. В простейшем случае однокамерная раковина имеет форму шара или колбы, с одним устьем (Saccammina, Lagena) или с несколькими отверстиями (Astrorhiza). Она может быть агглютинированной или известковой. При усиленном нарастании вдоль устьевого края возникает раковина в форме трубки, открытой с одной стороны или с обеих.
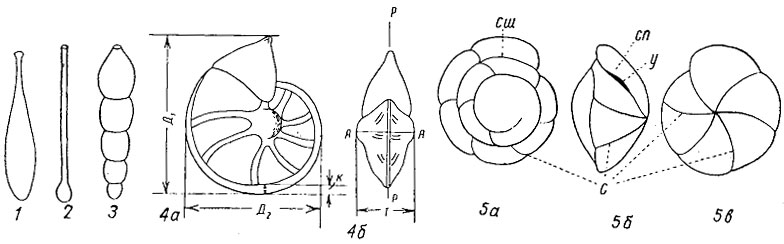
Рис. 29. Схема строения раковин фораминифер: 1 - однокамерная; 2 - двухкамерная; 3-5 - многокамерные: 3 - однорядная, 4 - спирально-плоскостная: 4а - сбоку, 4б - со стороны устья, 5 - спирально-коническая: 5а - с дорзальной стороны, 5б - со стороны устья, 5в - с вентральной стороны; АА - ось навивания, Д1 - большой диаметр, Д2 - малый диаметр, к - кнль, рр - плоскость симметрии, с - септальные швы, сп - септальная поверхность, сш - спиральный шов, Т - толщина раковины, у - устье
Двухкамерные раковины состоят из шаровидной начальной камеры и второй, длинной, неразделенной, трубчатой, отделенной от первой одной перегородкой. Вторая камера может быть прямой или разветвленной, или завитой в неправильную клубковидную, плоскую или коническую спираль.
Раковина, у которой внутренняя полость разделена перегородками, или септами, на камеры, называется многокамерной (рис. 29, 3-5). Возникновение многокамерности связано с изменением характера роста цитоплазмы и раковины. Рост из постоянного становится периодическим, причем периоды усиленного роста отделяются Друг от друга периодами покоя. Каждому периоду роста отвечает образование новой камеры, которая, как правило, бывает больше предыдущей; форма и расположение новой камеры и апертурной септы, отделяющей вновь образованную камеру от внешней среды, зависят от физико-химических свойств цитоплазмы, от величины краевых углов, образуемых расходящимися псевдоподиями со стенками предшествующей камеры, и от характера поверхности последней. Возникновение периодичности роста имело большое значение в развитии фораминифер, так как освобождало, их от необходимости непрерывно строить раковину. Следы такой периодичности можно уже наблюдать на некоторых одно- и двухкамерных трубчатых раковинах, несущих легкие пережимы.
Наиболее простой формой многокамерной раковины можно считать одноосную или однорядную, когда каждая последующая камера, имея форму шара как наиболее выгодную, обладающую наибольшим объемом при наименьшей поверхности, наращивается над предыдущей. Но у таких однорядных форм довольно велика опасность излома, особенно в местах пережимов, поэтому совершенствование формы приводит к тому, что новая камера охватывает своей основной частью часть предыдущей камеры, как бы надвигаясь на нее.
Другим способом укрепления раковины служит ее закручивание в спираль. Наиболее примитивным типом будет неправильно-клубковидный, при котором обороты навиваются беспорядочно в нескольких направлениях. При упорядочивании такого навивания возникают плектогирные раковины или раковины милиолинового типа. В первом случае ось навивания последующего оборота отклоняется, на некоторый угол от положения оси предыдущего оборота. Во втором случае камеры образуют спирально-свернутый клубок, располагаясь в нескольких взаимно пересекающихся плоскостях. Это объясняется тем, что направление оси навивания изменяется с ростом раковины на определенный угол. Длина каждой камеры составляет обычно половину оборота. У одних форм камеры отстоят друг от друга на 144° и располагаются в пяти плоскостях (Quinqueloculina), пересекающихся под углом 72° (см. рис. 42), у других камеры расположены в трех плоскостях (Triloculina), взаимно пересекающихся под углом 120°, и, наконец, у третьих, каждая камера расположена от предшествующей на 180° (Pyrgo, или Biloculina).
Спирально-плоскостной тип рассматривается как видоизмененный одноосный, у которого главная ось спирально завивается в одной плоскости. Линии соприкосновения смежных оборотов спирали раковины называются спиральными швами. Воображаемая прямая линия, вокруг которой происходит навивание оборотов раковины, носит название оси навивания. По оси навивания у спирально-плоскостных измеряется толщина раковины. Перпендикулярно к оси навивания через начальную камеру проводится диаметр, раковины. Сечение раковины перпендикулярное диаметру является экваториальным. С экваториальным сечением совпадает плоскость симметрии. Форма спирально-плоскостных раковин разнообразна и зависит от диаметра и толщины (см. рис. 41, 3). При диаметре, значительно превышающем толщину, раковина имеет дисковидную или чечевицеобразную форму. При диаметре, почти равном толщине, раковина приобретает шаровидную форму. При толщине, значительно превышающей диаметр, возникает веретеновидная форма. Если при рассмотрении спиральной раковины сбоку видны все обороты, она называется эволютной (см. рис. 35, 1), Если последний оборот охватывает все предыдущие обороты, то раковина называется инволютной (см. рис. 48, 5). Между этими двумя крайними типами строения имеется большое число форм, занимающих промежуточное положение (полуэволютные и полуинволютные).
Степень возрастания оборотов бывает различна. У большинства спирально-плоскостных раковин возрастание оборотов происходит постепенно, но у некоторых форм обороты возрастают очень быстро и раковина приобретает вид "рога изобилия" или даже становится веерообразной. Иногда быстрое возрастание оборотов может приводить к смыканию противоположных концов веера и к возникновению циклического типа раковины. У циклических раковин камеры расположены по концентрическим окружностям в одной плоскости (см. рис. 49).
При спирально-коническом типе (роталиевый) камеры располагаются по улитковидной, или трохоидной, спирали (рис. 29, 5). Сторону, отвечающую основанию конуса, где виден обычно только последний оборот, принято называть вентральной, или брюшной. Сторона, отвечающая вершине конуса, где видны все обороты, называется дорсальной, или спинной. Спиральный шов отделяет друг от друга спиральные обороты.
Спирально-винтовой тип раковин отличается тем, что высота нарастания камер происходит по высокой спирали, которая значительно превосходит диаметр основания (см. рис. 37). Обычно у таких раковин спиральное расположение камер выглядит как двух-, трех- или многорядное расположение камер и поэтому для них чаще употребляются названия двухрядные, трехрядные или многорядные раковины. У прикрепленных фораминифер раковина приобретает древовидную или неправильно разветвленную форму (см. рис. 34, 4).
Форма камер отличается большим разнообразием. Различают камеры: шаровидные, овальные, трубчатые, циклические, радиально удлиненные, угловатые (конические, ромбовидные, усеченно-конические), валикообразные.
Однако рассмотренными выше основными типами строения раковин не исчерпывается все многообразие их форм.
Гетероморфизм. Нередко в процессе индивидуального развития (онтогенеза) происходит изменение типа строения раковины, что приводит ее к гетероморфному строению. Например, начальная раковина может быть спирально-плоскостной, следующий отдел может состоять из двух редко расположенных камер и конечный отдел может быть однорядным. Такая раковина называется триморфной. Если раковина сочетает всего два типа строения, то она биморфна (см. рис. 37, 2б, в), и, наконец, если она однотипна по своему строению, то ее называют мономорфной. Наиболее резко гетероморфное строение раковины бывает выражено у микросферических особей (шизонтов).
Апертура, или устье. Отверстие, при помощи которого цитоплазма сообщается с внешней средой, расположенное в конце однокамерной или в последней септе многокамерной раковины, носит название устья, или апертуры. Последняя септа образует септальную, или устьевую, поверхность. При образовании новой камеры устье предыдущей камеры становится отверстием, соединяющим соседние камеры. Это отверстие называется фораменом (отверстие, дыра); отсюда весь подкласс получил название фораминиферы. Устье (рис. 30) расположено в центре, эксцентрично или в основании апертурной перегородки; оно может быть простым, т. е. состоять из одного отверстия разнообразной формы: округлой, овальной, щелевидной, крестообразной, ветвистой, радиально-лучистой. Сложное устье состоит из нескольких отверстий. Наиболее распространенным типом сложного устья является ситовидное устье, состоящее из многочисленных мелких отверстий. У многих фораминифер строение устья осложняется дополнительными образованиями, к числу которых относятся особые выросты, называемые зубными пластинками, или зубами. Они имеют важное таксономическое значение и служат, по-видимому, для укрепления края раковины и прикрепления пучка выходящих псевдоподий.

Рис. 30. Типы строения устья: 1 - простые; 2 - устья с зубом; 3 - удлиненное; 4 - лучистое, с устьевой камерной; 5 - дендровидное; 6 - добавочные устья; 7 - сложные
Кроме основного устья для выхода эктоплазмы наружу служат различные отверстия, имеющиеся в раковине. К их числу относятся тонкие каналы, пронизывающие стенку некоторых агглютинированных и известковистых микрозернистых и радиально-лучистых раковин; дополнительные устья расположены в разных местах: вдоль периферического края, вдоль шва и т. д.
Система каналов. У наиболее высоко организованных фораминифер (роталииды, нуммулитиды) имеется система каналов внутри раковины (рис. 31). Основными элементами этой системы являются спиральный и межсептальный каналы. Спиральный канал связан с брюшной лопастью каждой из камер; от него отходят межсептальные каналы, расположенные в полостях двойных септ и открывающиеся тонкими порами в швах. У некоторых роталиид система каналов очень сложна: наблюдается не один, а два спиральных канала, от которых отходят пупочные и межсептальные каналы.
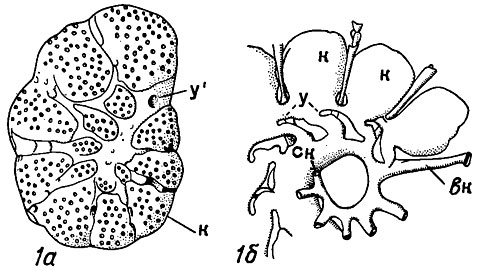
Рис. 31. Система каналов у роталиид: 1а - вид с вентральной стороны; 1б - внутренний слепок по продольному сечению; вк - внутрисептальный канал, к - камеры, ск - спиральный канал, у - устье, у' - устье спирального канала
Дополнительный скелет. К дополнительному скелету относятся те образования, которые усложняют строение раковины и септ. Они могут быть внутренними и наружными. К внутренним образованиям относятся известковые отложения, расположенные у эндотирид и фузулинид по краям экваториального устья (хоматы) или по бокам дополнительных устьев (парахоматы), или прерывисто только около септ (псевдохоматы). К ним относятся также столбики конической формы нуммулитид, пронизывающие раковину. На поверхности оборотов они имеют вид бугорков - гранул и служат для укрепления раковины.
К числу наружных дополнительных скелетных образований относятся различные скульптурные элементы в виде ребрышек, ячеек, килей, бугорков, игл, шипов и различных выростов на раковине.
У некоторых фораминифер, имеющих спиральную раковину, пупочная область бывает закрыта своеобразной втулкой или диском, состоящим из стекловатого кальцита; нередко этот диск бывает пронизан канальцами, связанными с внутренней системой каналов. У многих раковин планктонных фораминифер имеются тонкие длинные иглы, значительно увеличивающие их общую поверхность и облегчающие парение в толще воды.
Размножение и развитие. У фораминифер наблюдается сложный жизненный цикл развития (рис. 32), сопровождаемый чередованием бесполого и полового поколений. При половом размножении на каком-то этапе развития у особи, достигшей взрослого состояния, ядро делится на огромное число (тысячи) частиц, вокруг которых обособляется небольшая частица цитоплазмы. Таким путем возникают одноядерные клетки, снабженные двумя жгутами. Это половые клетки, или гаметы. По своему строению они совершенно одинаковы и благодаря жгутикам обладают подвижностью. После слияния двух гамет (оплодотворение), обычно происходящих от разных особей, возникает оплодотворенная клетка - зигота, имеющая диплоидный набор хромосом. Вокруг зиготы выделяется первая (эмбриональная) известковая камера. От нее, у многокамерных фораминифер, образуется вторая, третья и т. д. камеры. Зигота дает начало микросферическому поколению, или шизонту. Шизонт (форма В) сравнительно долгое время остается одноядерным, но с диплоидным набором хромосом. Затем на каком-то этапе роста происходит редукционное деление и ядро становится гаплоидным (с одинарным набором хромосом). При достижении шизонтом взрослого состояния ядро делится последовательно несколько раз и шизонт временно становится многоядерным; образуются десятки, а иногда свыше сотни маленьких ядер, вокруг которых обособляется цитоплазма. В этом случае возникают так называемые "эмбрионы", или амебовидные зародыши. Вокруг каждого "эмбриона" образуется довольно крупная эмбриональная камера. "Эмбрионы" покидают материнскую раковину и переходят к самостоятельному существованию. Этот процесс представляет собой бесполое размножение. Возникшие особи постепенно растут, строят новые камеры и дают макросферическое поколение, получившее название гамонтов (форма А).
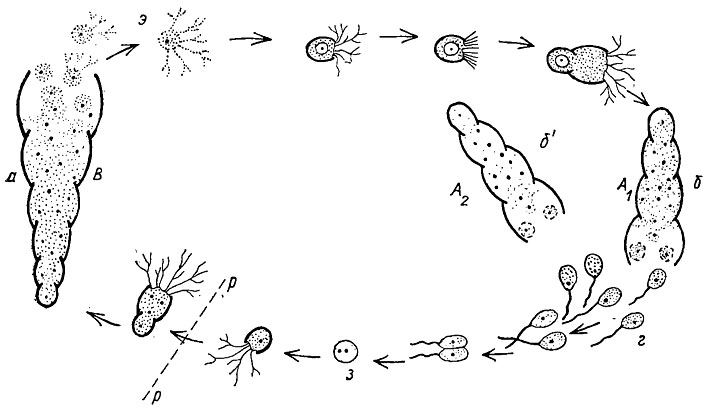
Рис. 32. Схема чередования поколений у фораминифер: а - микросферическая форма (шизонт В) с дочерними 'эмбрионами'; б, б' - мегасферические формы (гамонты А1, А2); г - гамета с гаплоидным (п) набором хромосом, з - зигота с диплоидным (2п) набором хромосом, рр - редукционное деление, э - дочерние 'эмбрионы'
Изучение онтогенеза фораминифер показало, что обычно наблюдается закономерное чередование гамонтов и шизонтов. Но иногда это закономерное чередование нарушается, за одним шизонтом (форма В) следует два поколения гамонтов (формы А1, А2). В одних случаях гамонты почти не отличимы или несколько отличаются по размерам, - в других - гамонты крупнее шизонтов и обладают большим числом камер, в третьих - гамонты и шизонты отличаются по размерам начальных камер. У макросферических особей начальная камера обычно крупных размеров, раковина сравнительно небольшая и число камер меньше, чем у микросферических особей. Последние отличаются маленькими размерами начальных камер, сравнительно большой раковиной и общим большим числом камер. Явление, связанное с образованием у фораминифер двух типов строения раковины, получило название диморфизма. Изучение диморфизма (или триморфизма) важно не только с точки зрения систематики, но и для изучения происхождения и родственных связей между фораминиферами. При этом более важное значение имеют особи, возникшие в результате полового процесса и более полно отражающие онтогенетическое развитие.
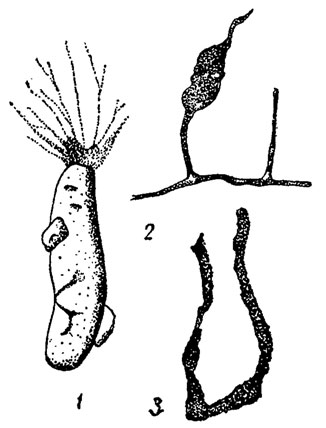
Рис. 33. Отряд Allogromiida: 1 - Chitinosaccus (совр.); 2 - Chitinodendron (поздний кембрий - силур); 3 - Chitinolagena (поздний ордовик)
Основы систематики и классификации. Важное значение для систематики фораминифер имеют строение и состав стенки раковины, строение цитоплазмы и ядра, особенности чередования поколений и другие признаки. На этом основании Д. М. Раузер-Черноусова и А. В. Фурсенко (1959) выделили 13 отрядов. Американские исследователи А. Леблик и Е. Таппан (1964) предложили разделять отряд фораминифер на пять подотрядов. В соответствии с принятым в учебнике рангом фораминифер в качестве подкласса эти подотряды подняты до уровня надотрядов. Подкласс фораминифер на основании строения стенки раковины разделен на пять надотрядов: Allogromioidea, Textularioidea, Fusulinoidea, Miliolidoidea, Rotalioidea.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'