
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Metazoa. Многоклеточные
Происхождение и развитие многоклеточных. Существует достаточно много гипотез о происхождении многоклеточных, недавно подробно и критически рассмотренных А. В. Ивановым в работе "Происхождение многоклеточных организмов" (1968). Большинством исследователей принимаются гипотезы происхождения многоклеточных от клеточных предков, и лишь немногие придерживаются мнения о независимом происхождении клеточных структур простейших и многоклеточных от доклеточных предков. Первая группа гипотез исходит из предположения о происхождении метазой от многоядерных или колониальных простейших (протозойные гипотезы). Наибольшее распространение получили теория гастреи Геккеля (1872) и теория фагоцителлы Мечникова (1877), предполагающие, что первичные многоклеточные были одиночными свободноподвижными животными.
Бионт простейшего представляет собой одну клетку, или колонию клеток, или полиэнергидный комплекс. Бионт многоклеточных в отличие от простейших состоит из большого числа клеток, дифференцированных на соматические (тканевые) и половые; соматические клетки образуют различного типа ткани и органы, которые специализируются на выполнении какой-либо одной или нескольких функций и подчиняются организму как единому целому. В то же время, если бионт простейшего состоит из колоний клеток, то составляющие его отдельные клетки дифференцированы слабо. Если бионт простейшего представлен одной клеткой, подобной тканевой клетке, то эта клетка в отличие от последней физиологически и экологически является самостоятельным организмом. Простейшие и многоклеточные отличаются строением ядерного аппарата и характером жизненного цикла. Если строение ядра и жизненные циклы простейших очень разнообразны, то для многоклеточных характерен специализированный способ деления ядра (типичный кариокинез, см. рис. 17) и определенный тип жизненного цикла, который для всех многоклеточных - от губок до человека, за небольшим исключением, сохраняется единым. Жизненный цикл состоит в следующем: мужская и женская гаплоидные гаметы сливаются вместе и образуют зиготу; диплоидная зигота начинает дробиться, распадаясь на ряд клеток, получивших название бластомер. В результате последовательного дробления размеры бластомеров уменьшаются и возникает шаровидной формы зародыш - морула без полости или бластула с первичной полостью внутри (рис. 58, 1, 2).
В процессе дальнейшего развития, обусловленного морфогенетическими корреляциями, образуется двухслойный зародыш, или гаструла. Образование гаструлы может происходить тремя путями: иммиграцией, инвагинацией и эпиболией (рис. 58, 3а-в).
При иммиграции часть наружных клеток бластулы, размножаясь, внедряется внутрь зародыша, где образует плотную внутреннюю клеточную массу, дающую у кишечнополостных начало энтодерме. Подобный тип зародыша Мечников (1877) назвал фагоцителлой; наружный слой ее клеток он назвал кинобластом, а внутреннюю клеточную массу - фагоцитобластом.
При инвагинации. Стенки бластулы на одном полюсе впячиваются внутрь и образуют энтодерму, выстилающую первичную пищеварительную полость, которая открывается наружу первичным ртом, или бластопором. Инвагинация наблюдается у сцифоидных медуз и актиний. Геккель (1872) назвал описанный гаструлоподобный организм гастреей и разработал теорию, в соответствии с которой от гастреи произошли все многоклеточные животные. Геккель, на основании работ Вольфа, Пандера, Бэра, А. О. Ковалевского (см. раздел "Основные этапы развития палеонтологии"), обосновал свой биогенетический закон и идею о гомологии эктодермы и энтодермы у всех многоклеточных. Геккель приписывал абсолютное рекапитуляционное значение начальным стадиям онтогенеза, считая, что каждой стадии соответствует определенный предок. Особенно важное значение он придавал гастрее; от нее все многоклеточные унаследовали первичный кишечник и два первичных зародышевых листка, из которых развились все остальные ткани. Мезодерма в процессе эволюции возникла позднее.
При эпиболии мелкие клетки (микромеры) обрастают более крупные клетки (макромеры). Этот способ образования зародышевых листков наблюдается у моллюсков.
Развитие мезодермы характерно для так называемых трехслойных многоклеточных. Мезодерма образуется двумя способами: телобластическим и энтероцельным (рис. 58, 4а, б).
При телобластическом способе в области бластопора гаструлы обособляются две крупные клетки - телобласты. Они делятся и образуют симметрично расположенные полоски клеток, которые в дальнейшем заполняют всю первичную полость гаструлы. Бластопор превращается в ротовое отверстие взрослого животного. Этот способ образования мезодермы характерен для большой группы трехслойных животных, объединяемых в подраздел первичноротых (Protostomia).
При энтероцельном способе образование мезодермы происходит иначе: от первичного кишечника отшнуровываются три пары мешковидных впячиваний, которые разрастаются и заполняют первичную полость гаструлы (рис. 58, 4б). Бластопор замыкается, рот возникает в другой части зародыша, а на месте бластопора образуется анальное отверстие. Этот способ формирования мезодермы характерен для второй группы трехслойных животных, объединяемых в подраздел вторичноротых (Deuterostomia). У первичноротых и вторичноротых внутри мезодермы в дальнейшем образуется вторичная полость тела, или целом, выстланная слоем - эпителиеподобных клеток.
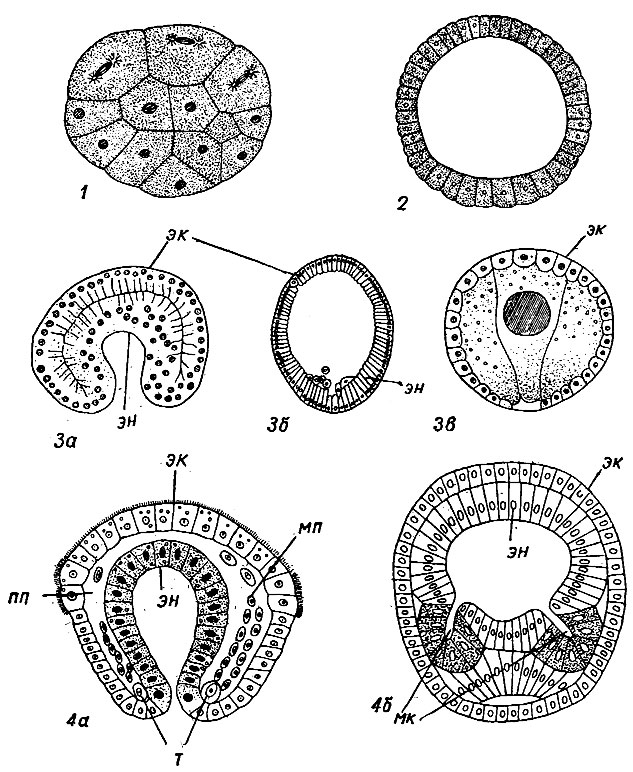
Рис. 58. Эмбриональное развитие многоклеточных: 1 - морула гидроидного полипа; 2 - бластула ланцетника; 3 - различные способы образования гаструлы: 3а - инвагинация, 3б - иммиграция, 3в - эпиболия; 4 - различные типы развития мезодермы: 4а - телобластический - у первичноротых, 4б - энтероцельный - у вторичноротых; мк - мезодермальные карманы, мп - мезодермальные полоски, пп - первичная полость, т - телобласт, эк - эктодерма, эн - энтодерма
Тело многоклеточных, таким образом, представляет собой в разной степени интегрированную и индивидуализированную систему, ограничивающую самостоятельность клеток, входящих в состав тела и подчиняющую себе их жизнедеятельность. Одной из важнейших черт прогрессивной эволюции многоклеточных является усиление интеграции их организма; чем выше степень его интеграции, тем выше выражена его собственная индивидуальность, тем более она подчиняет себе отдельные клетки, тем более стирается их индивидуальность. Каждый тип многоклеточных обладает определенным планом строения, определенным уровнем организации, свойственным только этому типу, и соответствующими ступенями симметрии.
В развитии многоклеточных, по мнению А. В. Иванова, можно выделить следующие стадии: 1) свободноплавающая шаровидная колония с периферическим расположением жгутиконосцев; все индивиды колонии однородны; размножение бесполое; 2) колония с клетками, дифференцированными на соматические и половые, с радиальной симметрией; появление полового процесса; 3) возникновение свободноплавающего организма типа мечниковской фагоцителлы и разделение клеток на наружный кинобласт и внутренний фагоцитобласт; разделение всего онтогенеза на эмбриональный период, когда развивалась бластулообразная личинка, и постэмбриональный, в течение которого осуществлялся рост, сопровождаемый делением клеток.
Дальнейшее развитие предположительно шло следующим путем (рис. 59). Губки отделились от основного ствола многоклеточных, когда у фагоцителлы еще не было ни рта, ни кишечника. Кинобласт у губок сменил локомоторную функцию на гидрокинетическую, погрузившись внутрь фагоцитобласта. В то же время, возможно, отделились археоциаты, крибрициаты и ряд групп неясного систематического положения. Позднее, когда у фагоцителлы возникло ротовое отверстие, от основного ствола возникли две новые группы многоклеточных. Одна дала начало гастрееобразным первичным кишечнополостным, вторая - бескишечным сколецидам, у которых еще не обособился кишечник и пищеварение происходило в фагоцитах - блуждающих клетках или в полостях паренхимы.
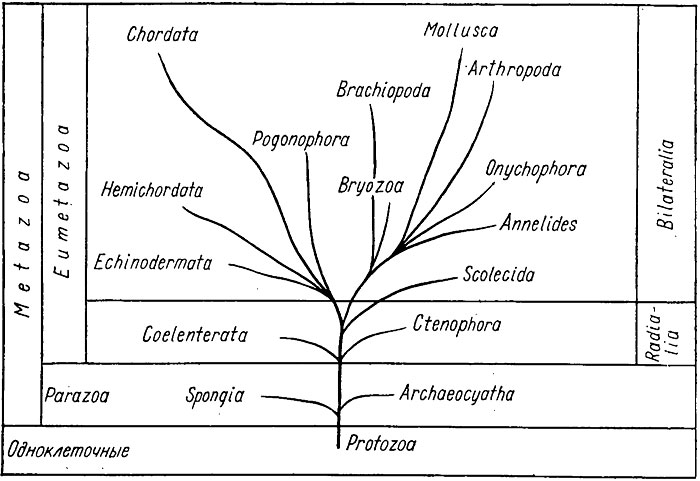
Рис. 59. Схема эволюционного развития животного мира
От первичных кишечнополостных возникли настоящие кишечнополостные, ведущие сидячий образ жизни (гидроидные и коралловые полипы) или перешедшие к подвижному образу жизни (сцифоидные медузы), и гребневики, приспособленные к жизни в пелагиали. Фагоцитобласт этих групп превратился в первичный пищеварительный орган - энтодерму, достигшую сложного строения. За счет кинобласта развилась эпителиальная ткань, покрывающая наружную поверхность тела. Кишечнополостные и гребневики, достигшие в своем развитии только стадии гаструлы (двухслойного мешка), объединяются в подраздел двухслойных многоклеточных. Как отмечалось выше, у всех остальных многоклеточных закладывается третий зародышевый листок - мезодерма и они образуют группу трехслойных животных. Среди первичноротых выделяют бесполостных и первичнополостных, к которым относятся низшие черви. У них появилась кровеносная система, выполняющая функцию газообмена и переноса питательных веществ, обособились мышечная и нервная ткани, у части низших червей возник сквозной пищеварительный тракт, появились органы выделения. Вторая группа первичноротых, наиболее разнообразная и многочисленная, представлена вторичнополостными, или целомическими, организмами. Это - кольчатые черви, онихофоры, членистоногие, моллюски, мшанки, форониды. Развитие каждой группы шло своим путем и будет рассмотрено ниже.
Развитие вторичноротых, к которым относятся иглокожие, гемихордовые и хордовые, шло по пути повышения организации, совершенствования всех систем тканей и органов и привело в конце концов к становлению человека. Иглокожие в своем развитии уклонились от основного ствола вторичноротых и в связи с прикрепленным или малоподвижным образом жизни приобрели радиальную симметрию тела и особую амбулакральную систему. Промежуточное положение между первичноротыми и вторичноротыми занимают брахиоподы. До сих пор неясным остается систематическое положение погонофор и хетогнат.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'