
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Подкласс ostracodoidea. Остракоды, или ракушковые рачки
Общая характеристика.Остракоды (гр. ostracon - раковина, черепок) - мелкие ракообразные, живущие в морских, пресноводных и солоноватоводных бассейнах. Маленькое тело заключено в карапакс, состоящий из двух створок. Большинство видов имеют микроскопические размеры (от 0,4 до 1,5 мм), некоторые пресноводные формы бывают до 5 мм, но среди свободноплавающих морских форм встречаются особи, достигающие 30 мм. У большинства остракод хитиновый карапакс пропитан солями кальция, гладкий или скульптированный, без следов нарастания. Во время линьки (8-9 личиночных стадий) происходит рост и изменение тела и скелета. Тело без следов сегментации. Голова несет простой глазок и пару сложных фасеточных глаз. Туловище снабжена от одной до трех пар ходильных ног; тело заканчивается обычно фуркой. Кровеносная система и органы дыхания отсутствуют. Нервная система состоит из двух окологлоточных ганглиев и брюшной цепочки. Раздельнополые; размножаются половым путем. В ископаемом состоянии известны с кембрия. Встречаются в морских, а начиная с карбона и в пресноводных отложениях.
Строение мягкого тела. Тело короткое, сжатое с боков, без следов сегментации, заключено в двустворчатую раковину, представляющую обызвествленный карапакс (рис. 135). Голова несет четыре пары видоизмененных конечностей - антеннулы, антенны, мандибулы и максиллы. Антеннулы, или первые антенны, одноветвистые, в типичном случае состоят из 8 члеников; выполняют локомоторную функцию (служат для плавания, ползания или зарывания) или функцию органов чувств, либо служат в качестве органов равновесия. Антенны также выполняют функцию локомоторных органов и служат для плавания, хождения или ползания. В отличие от антеннул они двухветвистые; прикрепляются по бокам головы около верхней губы. Мандибулы расположены по обе стороны рта; они состоят из протоподита, экзоподита и эндоподита. Протоподит снабжен зубом, предназначенным для жевания, эндоподит несет щетинки, которые служат для разрезания пищи и ее удержания. Остракоды используют мандибулы при зарывании и ползании. Макриллы развиты в разных группах по-разному и взаимосвязаны с мандибулами. Они служат для доставки пищи в рот, удаления несъедобных частиц и создания тока, воды для газообмена около головного отдела.
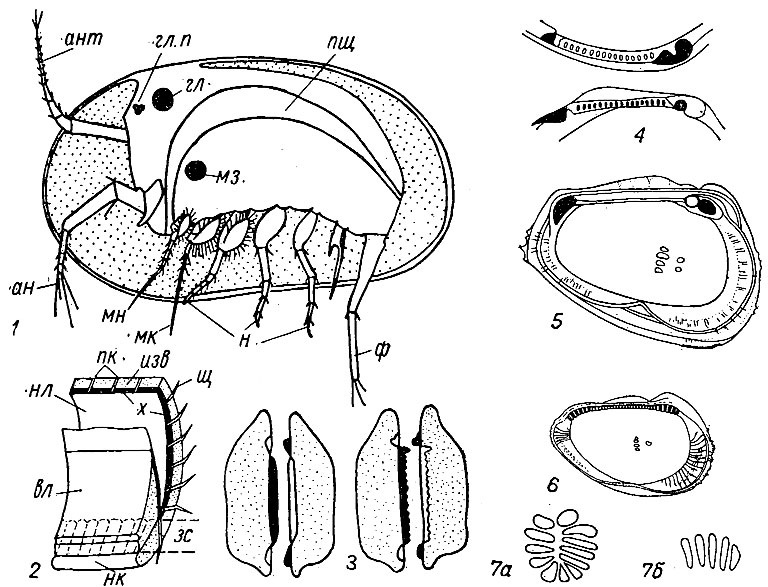
Рис. 135. Подкласс Ostracoda: 1 - схема строения современного представителя; 2 - схема строения края раковины; 3 - схема сочленения створок (зачернены зубы); 4 замок у Loxoconcha (мел - ныне); 5 - внутреннее строение створки Cythereis (мел - ныне); 6 - то же Protocythere (юра - мел); 7 - форма отпечатков мускулов; ан - антенна, ант - антеннула, вл - внутренний листок, гл - глаз фасеточный, глп - глаз простой, зс - зона сращения наружного и внутреннего листков, изв - обызвествленный слой, мз - мускул-замыкатель, мк - максилла, мн - мандибула, н - ножки, нк - наружный край раковины, нл - наружный листок, пк - поровые каналы, пщ - пищеварительный тракт, ф - фурка, х - хитиновый слой, щ - щетинки
Туловище несет от одной до трех пар конечностей. Первая пара туловищных ног прикреплена к телу около места соединения головы и туловища. Первая пара ходильных ног у одних остракод связана с челюстным аппаратом и рассматривается в качестве вторых максилл, у других выполняет функцию ходильных ног, причем часто ноги самцов отличаются от ног самок и наблюдается асимметрия - правая нога отличается от левой. Вторая пара туловищных ног, если присутствует, напоминает первую пару; она выполняет, правда не всегда, локомоторную функцию. Третья пара, развитая у части остракод, служит для очистки внутренней полости раковины или выполняет локомоторную функцию. Тело заканчивается фуркой разнообразной формы и строения, напоминающей по своему строению конечности.
Пищеварительный тракт состоит из рта, пищевода, желудка с пищеварительной железой и задней кишки, которая заканчивается анальным отверстием. Рот образован шлемовидной верхней губой и примыкающей к ней нижней, по бокам которой расположены мандибулы. Имеются слюнные железы. В желудок вдается хитиновый аппарат, фильтрующий пищу. Органы выделения представлены тремя парами выделительных желез. Органы кровообращения отсутствуют; только у морских миодокопид имеется пульсирующий пузырек - сердце. Газообмен осуществляется через покровы тела и внутренние листки створок, снабженных жаберными придатками. Нервная система состоит из надглоточного и подглоточного узлов (ганглиев) и брюшной нервной цепочки. Глаза представлены одним простым и 1-2 личиночными глазками или двумя фасеточными, расположенными по бокам головы, иногда отсутствуют. Створки скреплены вдоль спинного края лигаментом и соединены мускулом-замыкателем, который прикрепляется к внутренней поверхности створок, оставляя след прикрепления.
Все конечности снабжены поперечнополосатыми мышцами, приводящими их в движение. Следы прикрепления мышц к карапаксу также иногда сохраняются на внутренней стороне. Тело снабжено многочисленными щетинками, проходящими через поровую зону раковины. Кроме того, многие конечности несут щетинки, выполняющие функцию осязания.
Размножение и развитие. Остракоды раздельнополы, снабжены хорошо развитыми половыми органами, размножаются часто партеногенетически, при этом развитие идет вне тела матери или в особой выводковой камере. Из яйцевых оболочек выходит личинка науплиус (у морских форм), снабженная двустворчатой раковиной, одним глазком и тремя парами конечностей: антеннулами, антеннами и мандибулами. В процессе линьки сбрасывается не только карапакс, но и хитиновые покровы ряда частей тела, в том числе задней кишки; во время линек происходит последовательное развитие всех частей и органов тела. Размеры карапакса и его пропорции изменяются аллометрически. На второй стадии появляются максиллы, фурка и остальные конечности; на третьей - порово-канальная зона с узким пояском поровых каналов; на пятой стадии обычно появляется скульптура и начинает формироваться замок. После седьмой линьки появляются признаки полового диморфизма. Пресноводные остракоды достигают половозрелости обычно раньше морских. У остракод это время колеблется от 30 дней до трех лет.
Строение скелета. Карапакс остракод состоит из двух створок разнообразной формы и размеров (см. рис. 135, 2-6). Створки образованы двойной складкой кожи, которая прикрепляется к телу на границе головы и туловища. Она состоит из двух листков - наружного и внутреннего. Наружный листок образован двумя хитиновыми и находящимся между ними известковым слоями. Иногда хитиновые слои также обызвествляются. Внутренний листок представляет собой тонкую кутикулярную пластинцу, переходящую непосредственно в хитиновый покров животного. Вдоль внешнего края пластинка обызвествляется, образуя краевую пластинку. Внутренняя и наружная пластинки срастаются вдоль брюшного края, образуя так называемую зону сращения. В наружной пластинке и в зоне сращения имеются поровые каналы (так называемая порово-канальная зона), через которые выходят щетинки, выполняющие функции органов чувств.
В карапаксе различают дорзальную и вентральную части, передний и задний концы. Вдоль спинной части расположен замок, вдоль брюшного, переднего и заднего краев створки соприкасаются без сочленения. Створки могут быть равными и неравными, симметричными и асимметричными; у неравностворчатых раковин одна створка охватывает другую. Створки могут быть гладкими или скульптированными. К скульптуре относятся лопасти, бугры, ребра, шипы, борозды, ямки, ячейки. Лопасть представляет собой выпуклую часть створки, ограниченную бороздами; створка может иметь от двух до четырех лопастей, развитых преимущественно у палеозойских остракод (особенно бейрихиид). Бугор - высокий конический выступ на внешней части створки, расположенный чаще всего посредине створки. У многих форм выделяется также глазной бугорок, расположенный в переднеспинной части створки против глазка остракод. Ребра представляют собой выступы створок, расположенные в любой их части. Они могут быть продольными, поперечными и косыми. Иногда вдоль свободного края створки развивается краевое ребро. Шипы представляют собой скульптурные образования; они могут быть сплошными, полыми, крупными, мелкими. К ямкам относятся небольшие углубления на поверхности створок; к ячейкам - небольшие углубленные участки створок, окаймленные гранями. Сочетание ямок, ячеек, ребер, бугорков и шипов создает различные типы скульптуры, характерные для различных групп остракод.
На внутренней поверхности створок сохраняются (не всегда) следы прикрепления различных мускулов: замыкающего, мандибулярных, антеннальных и спинных, управляющих как конечностями, так и органами задней части туловища. Чаще всего сохраняются следы прикрепления мускула-замыкателя, расположенного ближе к переднему краю створок, примерно посередине. Эти следы имеют форму круглого пятна, состоящего из отдельных бугорков (иногда до 200) микроскопических размеров (см. рис. 135, 5-7). На внешней поверхности бугоркам обычно соответствуют ямки- или бороздки. У мезокайнозойских остракод сохраняются следы прикрепления мандибулярных мышц в виде двух отдельных бугорков; в спинной части створок над мускулом-замыкателем наблюдаются иногда следы прикрепления мускулов спинной группы, а впереди них следы прикрепления антеннальных мускулов.
Вдоль спинной стороны створки соединяются эластичной связкой - лигаментом и замком. Замок состоит из выступов - валиков или зубов и углублений - желобков, или ямок (см. рис. 135, 3-4). Строение замочного края различно. Он может иметь: а) гладкую замочную площадку, б) прямой или изогнутый, гладкий или зазубренный валик в одной створке и бороздку или желобок в другой створке, в) валик и бороздку, сопровождаемые кардинальным зубом и соответствующей ему ямкой, д) замок, состоящий из зубов в одной створке и ямок в другой либо чередования зубов и ямок в каждой из створок. Зубы могут быть гладкими или мелко зазубренными - кренулированными. Строение замочного края имеет важное значение для диагностики остракод. В последние годы выделено большое число типов замков, получивших соответствующие названия.
Половой диморфизм. Самцы обычно отличаются от самок не только по строению половой системы, но и по строению раковин, хотя половые отличия на раковинах не всегда выражены достаточно четко. Обычно раковины самцов более длинные и низкие, несут несколько отличную скульптуру. У древних остракод раковины самок выделяются по наличию выпуклостей у заднего конца створок, наличием выводковых камер (рис. 136, 4а). Интересно, что половые различия более четко выражены на раковинах палеозойских форм, чем на скелетах современных.

Рис. 136. Подкласс Ostracoda: 1 - отряд Archaeocopida, Bradoria (ранний - средний кембрий); 2 - отряд Leperditicopida, Leperditia (силур - девон): 2а - левая створка, 26 - правая; 3-4 - отряд Palaeocopida: 3 - Quadrilobella (ранний ордовик), 4 - Beyrichia (силур - средний девон): 4а - женская особь с выводковой камерой (вк), 46 - мужская особь; 5-7 - отряд Podocopida: 5 - Cythere (совр.), 6 - Bardia (ордовик - ныне), 7 - Cypris (? юра, плейстоцен - ныне); 8-9 - отряд Platycopida: 8 - Kloedenella (силур - карбон), поперечный разрез через переднюю (а) и заднюю (6) части (левая створка зачернена), 9 - Cytherella (юра - ныне); 10-12 - отряд Myodocopida: 10 - Cypridina (совр.), 11 - Cypridella (карбон), 12 - Cyprella (карбон), вид спереди; ин - инцизура; направление стрелки указывает передний край
Основы систематики и классификация. В основу выделения отрядов среди вымерших остракод приняты особенности строения карапакса, форма и строение отпечатков мускула-замыкателя, строение замка. При выделении отрядов среди ныне живущих представителей использованы особенности строения мягкого тела. В настоящее время остракоды разделены на 6 отрядов: Archaeocopida, Leperditicopida, Palaeocopida, Podocopida, Platycopida, Myodocopida.
Отряд Archaeocopida. Археокопиды (гр. archaios - древний, коре - весло). Карапакс обызвествлен, содержит большое количество хитина, делающего его эластичным и гибким. Замочный край прямой и длинный. Глазной бугорок обычно выступает. Рядом с глазным бугорком (немного позади и ниже) расположен отпечаток мускула-замыкателя. Отряд включает 12 родов (рис. 136, 1), распространенных в раннем и среднем кембрии. Предположительно к этому отряду отнесен род Eremos из тремадока Швеции.
Отряд Leperditicopida. Лепердитикопиды. Карапакс известковый, толстый, крупных размеров; замочный край прямой. Отпечаток мускула-замыкателя крупный, составляет до 30% высоты створки и состоит из многочисленных бугорков (до 200). Поверхность карапакса гладкая или слабо скульптированная. Появились в конце кембрия, были широко распространены в ордовике и силуре и дожили до конца девона. Известно 15 родов (рис. 136, 2).
Отряд Palaeocopida. Палеокопиды (гр. palaios - древний, коре - весло). Карапакс с прямым смычным краем, крупных размеров. Поверхность гладкая, но чаще скульптированная; имеются лопасти, бугры, ребра. Обызвествленная внутренняя пластинка отсутствует. Часто наблюдается половой диморфизм. Наиболее распространенный отряд палеозойских остракод, насчитывающий до 260 родов (рис. 136, 3-4). Первые представители появились в ордовике, максимального развития достигли в силуре-девоне, но несколько родов существовало в карбоне- перми; одно семейство существует в современных морях.
Отряд Podocopida. Подокопиды. Раковина от овальной до закругленно-прямоугольной, неравностворчатая, иногда с оттянутым задним концом; на конце могут быть шипы; поверхность раковины гладкая или скульптированная. У современных представителей антеннулы несут когтевидные щетинки, мандибулы и максиллы имеют жаберные придатки. Отпечаток мускула-замыкателя округленный, состоящий из многочисленных бугорков у ранних представителей и небольшого числа у более поздних. Широко распространены во всем мире и встречаются как в морских, так и в пресноводных водоемах. Насчитывается около 450 родов (рис. 136, 5-7).
Отряд Platycopida. Платикопиды (гр. platys = platos - плоский). Отряд объединяет остракод с овальной неравностворчатой раковиной, с необособленной порово-канальной зоной; у самок задняя часть раковины вздутая; туловищный отдел несет две пары конечностей; отпечаток мускула-замыкателя состоит из двух рядов бугорков, расположенных в передней трети раковины. Каждый ряд состоит из 5-9 бугорков. Спинной край округленный. Ордовик - ныне. Несколько родов (рис. 136, 8-9).
Отряд Myodocopida. Миодокопиды (гр. myodos - мускул, kope - весло). Карапакс состоит из двух сильно выпуклых створок, несущих на переднем крае выступ - ростр и инцизуру - отверстие для выхода антенн; поверхность створок раковин гладкая, редко ребристая; у палеозойских форм со срединной бороздой; раковина обычно слабо обызвествлена. Размеры колеблются от 20 до 30 мм. На последней стадии жизненного цикла ведут планктонный образ жизни. Первые представители появились в ордовике. В палеозое существовало 30 родов; из мезозоя и кайнозоя известно только 2 рода, в современных морях насчитывается около 25 родов (рис. 136, 10-12).
История развития и экология остракод. К наиболее древним остракодам относятся представители отряда археокопид, появившиеся в начале кембрия и имевшие слабо обызвествленный карапакс с прямым смычным краем, крупным отпечатком мускула-замыкателя, расположенным непосредственно под глазным бугорком. Они имеют некоторое сходство с конхостраками, к которым были вначале отнесены. Слабое обызвествление карапакса отличает их от остальных остракод. По-видимому предки остракод, возможно существовавшие уже в конце протерозоя, имели хитиновый покров. От археокопид в конце кембрия появились лепердитикопиды. Они обладали крупным карапаксом, сильно обызвествленным, сложным мускулом-замыкателем, имеющим форму шеврона. Лепердитикопиды были широко распространены в ордовикских и силурийских морях Европы, Азии и Северной Америки.
В начале ордовика возникают палеокопиды - широко распространенные в палеозое остракоды, среди которых наибольшее значение имеют бейрихииды, образующие самостоятельный подотряд среди палеокопид. Бейрихииды отличались крупной равностворчатой раковиной, обычно сильно расчлененной, снабженной шаровидными образованиями; они были наиболее широко распространены в силуре и девоне. Основная группа палеокопид вымирает в середине перми; в современных морях обнаружен один род.
В начале ордовика ответвилась новая ветвь остракод, выделенная в отряд подокопид, разделившаяся с самого начала на две ветви - собственно подокопин и метакопин. Собственно подокопины представлены, большой группой бердиид (Bairdiacea), получивших наибольшее развитие в карбоне (но доживших до наших дней), цитераций (Cytheracea) развитых в мелу и кайнозое. Все они несут 7 пар конечностей, простые глазки (иногда глаза отсутствуют). Тело заключено в округленную овальную раковину с хорошо дифференцированным замком и развитой порово-канальной зоной. Цитерации кроме морских заселили также пресноводные бассейны. Их представители известны из континентальных, отложений верхней перми Кузбасса и Русской платформы и из солоноватоводных бассейнов раннего триаса Поволжья и Эмбы. Вторая ветвь подокопид - метакопины - отличается бобовидной или яйцевидной раковиной с крупным отпечатком мускула-замыкателя, состоящего из немногочисленных относительно круглых бугорков. Их надсемейство ципридации - обитатели пресноводных бассейнов начиная с триаса. В ордовике возникли платикопиды, у которых бугорки для мускула-замыкателя (13-14) расположены двурядно в передней трети раковины. Створки обычно неравные, с изогнутым замочным краем.
Особое место среди остракод занимают представители отряда миодокопид, появившиеся в начале ордовика. Они имели крупную раковину до 30 мм, снабженную на переднем конце ростральным выступом а особым отверстием - инцизурой, служащей для постоянного выхода антенн. Современные миодокопиды ведут планктонный образ жизни. Раковина часто слабо обызвествлена.
До настоящего времени, таким образом, доживают представители четырех отрядов: подокопид, платикопид, миодокопид и палеокопид. Современные остракоды обитают в самых разнообразных бассейнах - от морских до пресноводных. В морях планктонный образ жизни ведут миодокопиды. Подокопиды встречены в морских и континентальных бассейнах. Морские подокопиды ведут бентосный образ жизни, обитая обычно на глубинах до 200 м, но чаще в более мелководных зонах и преимущественно на водорослях. Многие остракоды живут в илистых осадках, ползая или зарываясь. Подокопиды континентальных водоемов приспособились к жизни в разных условиях. Они живут в пресных и соленых озерах, реках, источниках, прудах, в торфяниках и даже в подземных водах. Впервые подокопиды приспособились к жизни в континентальных бассейнах в конце карбона (дарвинулиды). Они встречены вместе с остатками пресноводных моллюсков, червей и рыб.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'