
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Класс Bivalvia. Бивальвии, или двустворчатые моллюски
Общая характеристика. Бивальвии (гр. bi - два, valva - створка) - морские, солоноватоводные или пресноводные, малоподвижные или неподвижные моллюски бентоса; тело заключено в двустворчатую известковую раковину, состоящую из двух равных или неравных створок, соединенных на спинной стороне лигаментом, служащим для их открывания, и одним или двумя мускулами-аддукторами - закрывания створок (рис. 165). Для прочного соединения двух створок вдоль смычного края может быть развит замок. Лопасти мантии, сросшейся на спинной стороне, соединяются с краем раковины на внутренней стороне паллиальными мускулами, оставляющими след на нем в виде мантийной, или паллиальной, линии. Нога служит для ползания, зарывания и прикрепления, имеет биссусную железу (у Autobranchia и Septibranchia). В мантийной полости, сзади, расположены парные жабры, выполняющие функцию газообмена.
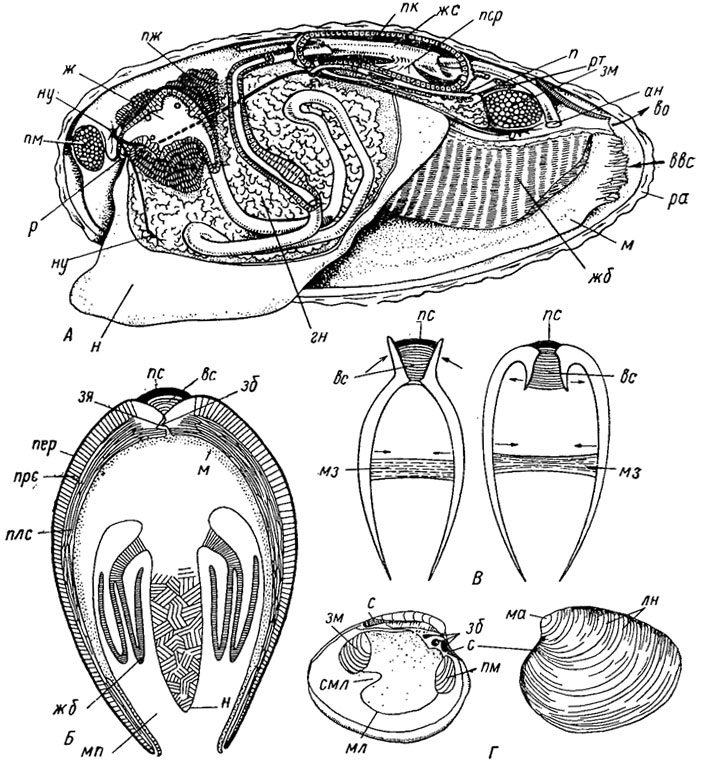
Рис. 165. Класс Bivalvia: А - продольный разрез через тело пресноводной Anodonta; Б - схематический поперечный разрез; В - схема взаимодействия мускула-замыкателя (мз) и связки (лигамента); Г - строение раковины Venus; ан - анальное отверстие, вес - вводной сифон, во - выводной сифон, вс - волокнистая связка, гн - гонады, ж - желудок, жб - жабры, жс - желудочек сердца, зб - зубы, зм - задний мускул, зя - зубная ямка, ли - линии нарастания, м - мантия, ма - макушка, мз - мускул-замыкатель, мл - мантийная линия, мп - мантийная полость, н - нога, ну - нервные узлы, п - почка, пер - периострак, пж - пищеварительная железа, пк - перикардий, плс - перламутровый слой, пм - передний мускул, прс - призматический слой, пс - пластинчатая связка, пер - предсердие, р - рот, ра - раковина, рт - ретрактор ноги, с - связка, смл - синус мантийной линии
Голова не обособлена от туловища; отсутствуют челюсти, радула, слюнные железы. Пищеварительный тракт состоит из короткого пищевода, объемистого желудка, печени из трех долей, открывающихся в желудок с вентральной стороны, и длинной задней кишки. Кровеносная система незамкнутая; сердце состоит из желудочка и двух предсердий. У протобранхий имеются нервные стволы, у остальных ганглии расположены ниже рта, в ноге и позади заднего мускула. Имеется одна пара деломодуктов (почки связаны с гонодуктами). Двустворчатые моллюски гермафродиты и раздельнополые. В верхней части ноги расположены парные гонады. Половой диморфизм выражен слабо. Оплодотворение внешнее, в воде. Индивидуальное развитие сопровождается личиночными стадиями - трохофорой и велигером; у пресноводных наблюдается живорождение или развитие особого типа личинок - глохидий, лазидий или гаусториальных личинок, временно паразитирующих на рыбах. В ископаемом состоянии известны со среднего кембрия, но существенную роль в фауне бентоса начинают играть с конца триаса занимая разнообразные экологические ниши, занятые в палеозое брахиоподами.
Строение мягкого тела. Тело двусторонне-симметричное, несегментированное, покрыто с боков двумя лопастями мантии, сросшейся на спинной стороне. Голова необособлена от туловища, сжатого с боков, поэтому бивальвий иногда называют Acephala - безголовые (а - отрицание, cephalon - голова); на брюшной стороне тела развита мускулистая нога, имеющая удлиненную, червеобразную или клиновидную форму (отсюда еще одно название Pelecypoda - секироногие, от гр. peles - секира), у примитивных представителей с уплощенной подошвой. В ноге находится биссусная железа, выделяющая клейкие биссусные нити, служащие для прикрепления моллюска к подводным предметам. Нога выходит между створок и служит для ползания, зарывания в рыхлый осадок, помогает при сверлении твердых известковых пород у прикрепленных форм бывает недоразвита или редуцирована полностью.
Все внутренние органы заключены в раковину, выделяемую мантией. Мантия двумя лопастями охватывает тело с боков. В задней части обособлена мантийная полость. Лопасти мантии соединены вдоль спинного края и у одних остаются с брюшной стороны свободными (рис. 166); у других срастаются сзади, оставляя отверстие для ввода и вывода воды и продуктов обмена из мантийной полости; у третьих срастаются спереди и снизу, оставляя отверстие для ноги. У многих форм задние части мантии вытягиваются в трубки разной длины, образуя вводной и выводной сифоны. Сифоны могут быть раздельными либо срастаются вместе по всей длине или только частично. У одних двустворок при закрывании раковины сифоны с помощью мускулов втягиваются внутрь раковины, у других остаются в течение всей жизни выставленными наружу. Сифоны развиты у двустворок, ведущих зарывающийся образ жизни.
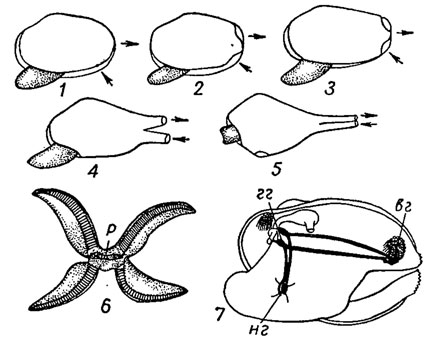
Рис. 166. Класс Bivalvia: 1-5 - схема строения мантии и сифона (см. текст); 6 - рот и губы Mitilus edulis, вид спереди; 7 - схема строения нервной системы; ганглии: вг - висцеральный, гг - головной, нг - ножной; р - рот; стрелки указывают ввод и вывод воды
Край мантии имеет три складки, выполняющие различные функции (рис. 167). Наружная складка выделяет периострак и внешний известковый слой; средняя складка несет небольшие щупальца и другие органы чувств; внутренняя складка снабжена многочисленными мелкими мускулами (паллиальными), при помощи которых мантия прикрепляется изнутри к раковине, образуя мантийную, или паллиальную, линию, проходящую почти параллельно внешнему краю раковины. У двустворок, имеющих сифоны, в задней части тела обособлено место для втягиваемых внутрь раковины сифонов, ограниченное мантийным, или паллиальным синусом. По глубине мантийного синуса судят о размерах сифонов. У форм, не втягивающих сифоны, раковина остается сзади зияющей. Двустворки, живущие на поверхности дна, сифонов не имеют.
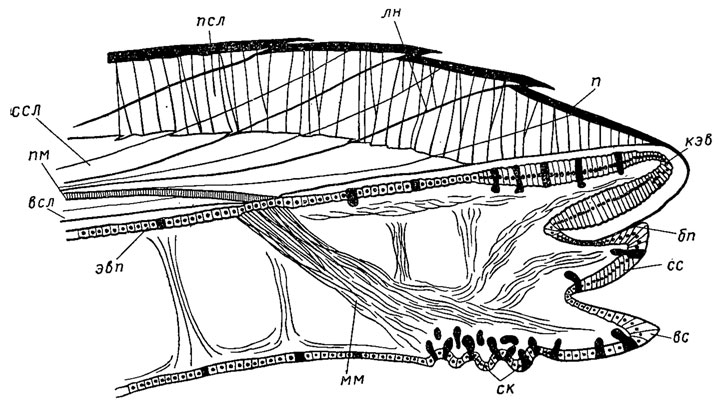
Рис. 167. Схема строения переднего края мантии Anadonta cygnea (Тейлор, Кеннеди, Холл, 1969): бп - борозда периострака, вс - внутренняя складка, всл - внутренний слой, кэв - клетки эпителия внешней складки мантии, лн - линии нарастания, мм - мантийный мускул, п - периострак, пм - палиальный миострак, псл - призматический слой, ск - слизистые клетки, сс - срединная складка, ссл - средний слой, эвп - эпителий внешней поверхности мантии
Раковина состоит из двух створок, соединенных связкой, или лигаментом, и замыкающими мускулами, или аддукторами. Мускулы, один или два, являются производными мантийной мускулатуры. Они служат для замыкания створок, в то время как связка - для открывания. Парные мускулы обычно расположены ближе к спинной стороне; непарные - ближе к центру. Можно проследить все стадии редукции переднего мускула и усиление заднего. Поэтому среди двустворчатых различают: равномускульных, неравномускульных и одномускульных.
Основными органами дыхания служат жабры, расположенные симметрично с боков в мантийной полости в задней части тела (рис. 168). У примитивных двустворок жабры имеют вид лепестков или пластинок, расположенных в два ряда с каждой стороны под углом друг к другу (гребенчатые жабры). Пластинки соединяются между собой соединительной мембраной, вдоль которой в нижней и верхней частях проходят кровеносные сосуды. Нижняя и боковые поверхности пластинок покрыты мерцательным эпителием, создающим ток воды к ротовому отверстию. У большинства двустворок имеются нитчатые жабры, состоящие из коленообразно изогнутых нитей, свисающих в мантийную полость. Нити наружного ряда изгибаются наружу, внутреннего - внутрь. Дальнейшее усложнение жабр связано с образованием перемычек между нитями и превращением их в пластинчатые решетки (отсюда название класса Lamellibranchiata, или пластинчатожаберные). Оба типа жабр служат не только для газообмена, но и для сортировки и транспортировки питательных частиц к ротовому отверстию, являющемуся своеобразным фильтром-ситом.
Особым типом строения жабр отличается небольшая группа преимущественно глубоководных двустворок, у которых мантийная полость разделена горизонтальной или изогнутой перегородкой, или септой, на верхний и нижний отделы (септальные жабры); в септе для прохождения воды имеются ситовидные пластинки с мелкими отверстиями или несколько более крупных отверстий (рис. 168, 4). С помощью мышц септа приводится в движение, образуя своеобразный мембранный насос для создания тока воды. У этих бивальвий жабры служат только для добывания пищи.
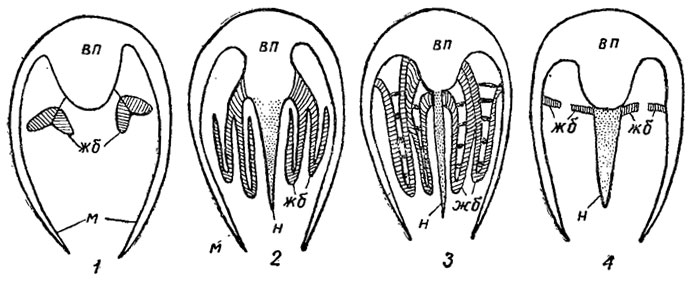
Рис. 168. Схема строения жабр: 1 - гребенчатые, 2 - нитевидные, 3 - пластинчатые, 4 - септальные; вп - висцеральная полость, жб - жабры, м - мантия, н - нога
Ротовое отверстие имеет форму щели. Вокруг рта расположены две пары ротовых лопастей, покрытых, мерцательным эпителием. У нукулид ротовые лопасти имеют длинные хоботовидные придатки с желобком посредине; они проходят в отверстие между створками позади ноги и служат для сбора пищи (см. рис. 175). По желобкам лопастей пища поступает в рот.
Пищеварительная система состоит из короткого пищевода, удлиненного желудка, средней и задней кишки, печени и кристаллического стебелька. Челюсти, радула и слюнные железы отсутствуют. Пища передвигается мерцательным ресничным эпителием. Кристаллический стебелек, как и у гастропод, перемешивает пищу, растворяясь, выделяет ферменты, способствующие внеклеточному перевариванию. Стенка желудка напротив стебелька превращена в своеобразный желудочный щит. Мелкие пищевые частицы поступают в печень, состоящую из слепых выростов - дивертикул, в которых происходит внутриклеточное переваривание и всасывание; в этом процессе важную роль играют блуждающие клетки - амебоциты. Основной пищей двустворок служат фитопланктон, зоопланктон, бактерии и органический детрит, исключение составляют септибранхии - хищники, и тередины, питающиеся древесиной.
По особенностям внутреннего строения различают несколько типов желудков - митилоидный, пектиноидный, астартоидный и др. Из желудка остатки пищи попадают в среднюю кишку, проходящую через стенку перикардия, желудочек сердца (создается впечатление протыкания сердца кишечником), далее в заднюю кишку, и фекалии выводятся через анальное отверстие наружу. Фекалии, продукты обмена и непереваренные частицы облекаются слизью и превращаются в плотные массы шаровидной, овальной или лентовидной формы, часто снабженные бороздами, отражающими соответствующие выступы тифлозоля. Известен один случай находки в ископаемом состоянии остатков пищеварительного тракта, сохранившегося в раковинах у Nuculana, из ранней юры Англии вместе с ожелезненными остатками фекалий. По-видимому захоронение и ожелезнение произошли очень быстро.
Кровеносная система незамкнута; сердце состоит из желудочка и двух предсердий, кровь сокращением сердца гонится к органам тела по артериям и венам, по системе лакун и синусов, расположенных в соединительной ткани. У низших двустворок имеются два желудочка, лежащие по бокам кишки, или непарный желудочек, лежащий над или под кишкой. У более развитых двустворок два желудочка срослись в один и кишка оказалась внутри сердца. Кровь обеспечивает внутренние ткани и органы кислородом и питательными веществами, уносит продукты обмена, поддерживает постоянство внутренней среды и способствует созданию в ноге гидравлического давления, необходимого при ползании и особенно при закапывании и регулируемого особым клапаном (кеберов орган). У септибранхий внутренняя поверхность мантии принимает на себя дыхательные функции, обогащая кровь кислородом. Лакуны и синусы представляют собой первичную полость тела, они не имеют собственных стенок.
Вторичная полость тела редуцирована; она представлена двумя небольшими полостями: перикардием, или сердечной сумкой, и полостью гонад. С перикардием связаны перикардиальные железы, выполняющие наряду с почками выделительную функцию. Почки имеют вид двух трубочек или мешков, которые сообщаются с одной стороны с перикардием, а с другой - через почечные отверстия с мантийной полостью. У низших двустворок почки имеют вид простых трубочек, покрытых выделительным эпителием; они связаны также с половыми железами. У большинства, почки отделены от половых протоков и устроены сложнее.
Нервная система развита слабо, что обусловлено малоподвижным образом жизни (см. рис. 166, 7); она симметрична и состоит из трех пар ганглиев, соединенных длинными тяжами (коннективами); одна пара лежит над ртом и иннервирует ротовые лопасти, передний мускул-замыкатель и переднюю часть мантии, вторая пара расположена глубоко в ноге, третья - позади заднего мускула-замыкателя, иннервируя: все внутренние органы, задний мускул и заднюю часть мантии; каждая, пара ганглиев соединена поперечными перемычками (комиссурами). Органы чувств развиты слабо; к ним относятся: органы кожного чувства - чувствительные клетки, выполняющие осязательные функции и расположенные на щупальцах, по краю мантии и сифонов, по краю ноги, на бугорках около анального отверстия; осфрадии - жаберные органы чувств, сходно устроенные с таковыми примитивных гастропод; статоцисты - органы равновесия, устроенные также сходно с подобными органами других моллюсков, они расположены около педального ганглия; глаза, или фоторецепторы, расположенные по краю мантии, на первых жаберных нитях, вокруг отверстий сифонов; они представляют собой не мозговые глаза в обычном смысле, а новообразования от простых эпителиальных до довольно сложных глаз с хрусталиком и сетчаткой.
Двустворчатые моллюски раздельнополы, очень редко гермафродиты; имеют парные гонады, расположенные внутри целома; протоки гонад связаны с протоками почек или чаще имеют самостоятельные отверстия. Оплодотворение происходит в воде. Развитие морских и пресноводных форм происходит разными путями. У морских форм развивается личинка трохофора, похожая на трохофору полихет. Раковина, выделяемая раковинной железой, лежит на спинной стороне личинки. Вначале это непарная кутикулярная пластинка, которая, постепенно разрастаясь, складывается вдоль, образуя две створки. Затем вся верхняя часть ноги личинки превращается в парус, или вел ум, - диск, покрытый ресничками; трохофора превращается в типичную для бивальвий личинку - велигер (рис. 169). Первичная раковинка велигера, или продиссоконх (гр. pro - префикс, означающий вначале, впереди, dissos - парный, konche - раковина), состоит из двух створок, соединенных вдоль спинного края тонкой конхиолиновой пленкой (лигамент еще не развит), личиночными зубами и одним, а позднее двумя мускулами-замыкателями. В течение планктонной стадии идет процесс формирования многих органов (органогенез), при этом часть личиночных органов исчезает, например велум. Затем личинка опускается на дно и превращается в молодого моллюска. Начинает интенсивно расти нога, служащая для передвижения или закапывания, начинает действовать биссусная железа, выделяющая клейкие нити. У прирастающих форм (устриц) биссусная железа выделяет вещество, прикрепляющее створку к подводным предметам. Постларвальная раковина (диссоконх) быстро приобретает облик взрослых форм; у нее начинают формироваться зубной аппарат и связка.
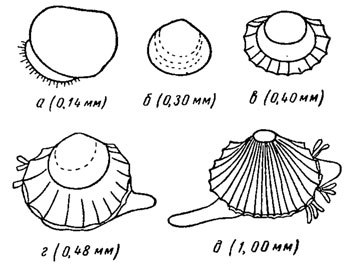
Рис. 169. Развитие Cardium edule L.: а - личинка-велигер, б-д - послеличиночные стадии
Эмбриональное развитие у пресноводных форм протекает иначе. У всех, за исключением дрейссенид, имеющих личиночные стадии, и унионид, эмбрионы развиваются в мантийной полости материнского организма и оттуда выходит вполне сформировавшийся моллюск (живорождение). У унионид в мантийной полости развивается личинка - глохидия, тело которой заключено в треугольную двустворчатую раковину, имеющую один мускул-замыкатель. Через выводной сифон глохидии выбрасываются в воду и прикрепляются особым зазубренным шипом раковины и биссусными нитями к телу рыб, превращаясь постепенно в эндопаразитов, которые питаются осмотическим путем с помощью личиночной мантии. Через два-три месяца глохидии, претерпев соответствующий метаморфоз и органогенез, покидают тело хозяина и молодые моллюски переходят к самостоятельному образу жизни. Остается неясным, когда у унионид впервые развилось подобное приспособление к паразитическому существованию, играющему важную роль в расселении пресноводных моллюсков. У южноамериканских пресноводных форм развивается личинка-лазидий, у африканских - гаусториальная личинка.
Строение раковины. Раковина бивальвий в типичном случае состоит из двух равных створок, охватывающих тело с правой и левой сторон, в соответствии с чем различают правую и левую створки. Обе створки соединяются вдоль смычного края лигаментом и замком. Замок расположен на спинной стороне и вместе с выступающей макушкой позволяет определить верхний (замочный), или спинной, край створок; противоположный край называется брюшным, или нижним; положение рта определяет передний край, а положение анального отверстия и сифонов - задний. Если макушка занимает центральное положение, то такие створки называются, равносторонними; если макушка сдвинута вперед или назад - неравносторонними. Раковина с загнутой вперед макушкой называется прозогирной (гр. gyros - округленный), с загнутой назад - опистогирной и ортогирной, если макушки двух створок направлены друг к другу. У прирастающих форм чаще всего раковина асимметричная, неравностворчатая: одна створка может иметь форму глубокой тарелки или конуса, а вторая - плоской крышки. Конусовидные раковины конвергентно сходны с рихтгофениями (из брахиопод) или скелетами одиночных кораллов; прирастающая створка отличается от свободной своей толщиной и массивностью.
В замкнутом состоянии створки обычно плотно прилегают друг к другу; у зарывающихся и сверлящих форм на переднем крае бывают развиты зияния для выхода ноги, а на заднем - для выхода сифонов.
Такие раковины называют зияющими; у некоторых сверлильщиков (тередо) для защиты длинных сифонов в процессе эволюции выработалась ложная раковина в виде длинной трубки. Редукция раковин наблюдается у тех бивальвий, у которых створки обрастают мантией; раковина в этом случае становится тонкостенной, зияющей, с редуцированным замком и мускулами-замыкателями.
Внешняя форма раковины бивальвии достигает большого разнообразия и отражает черты приспособления этих моллюсков к различным биотопам и условиям обитания (см. рис. 188).
В последние годы в связи с применением электронного микроскопа получены новые данные о структуре стенки раковины у бивальвий.
Стенка, как у всех моллюсков, состоит из органического и известкового слоев. Наружный органический слой, или периострак, образуется клетками эпителия внешней складки мантии; он постепенно утолщается от макушки к брюшному краю. Известковая стенка раковины обычно состоит из трех слоев - наружного, среднего и внутреннего; они образуются разными участками эпителия переднего края мантии (см. рис. 167).
Различают следующие микроструктуры: простая призматическая, сложная призматическая, перламутровая, перекрещенно-пластинчатая, сложная перекрещенно-пластинчатая, листоватая, гомогенная, гранулярная (рис. 170). Призматическая микроструктура построена из призм арагонита или кальцита с кристаллической осью С, расположенной вдоль длинной оси призмы; у некоторых наблюдается сложная призматическая микроструктура (нукулиды, венериды) - призмы первого порядка построены из призм второго порядка. Перламутровая микроструктура состоит из многочисленных многоугольных листочков или пластин арагонита, всегда параллельных внутренней поверхности раковины, налегающих друг на друга. Ось С всегда перпендикулярна плоскости таблички; ось В ориентирована в направлении роста. Наиболее распространенным типом микроструктуры является арагонитовая перекрещенно-пластинчатая, у которой отдельные пластины построены иглообразными удлиненными кристаллами, причем в соседних двух пластинах первого порядка пластины второго порядка имеют противоположное направление (рис. 170, 5). Сложная перекрещенно-пластинчатая микроструктура возникла из простой и отличается от нее более разнообразной и неправильной ориентировкой блоков таких пластин (рис. 170, 6). Листоватая микроструктура состоит из более или менее правильно расположенных листочков кальцита, лежащих рядом или черепицеобразно налегающих друг на друга. К гомогенной микроструктуре относится такая тонкокристаллическая структура, в которой отдельные кристаллы неразличимы; ее происхождение до сих пор не выяснено. Гранулярная микроструктура всегда арагонитовая, состоит из мелких карбонатных гранул, обычно с одинаковой кристаллографической ориентировкой.
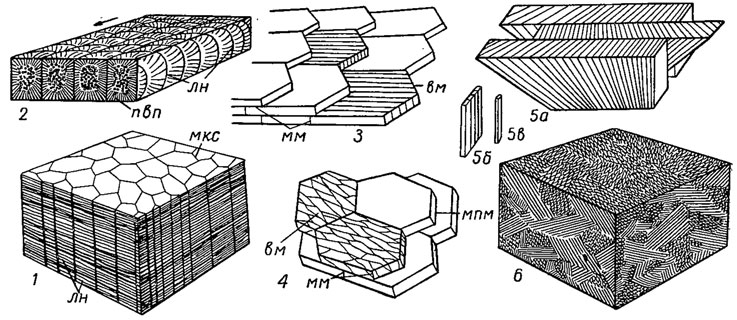
Рис. 170. Основные типы микроструктур стенки раковины: 1 - простая призматическая; 2 - сложная призматическая (стрелка - направление к макушке); 3 - листоватая; 4 - перламутровая; 5-6 - простая (5) и сложная (6) перекрещенно-пластинчатая (5б - пластина 2-го порядка, 5в - пластина 3-го порядка) (Тейлор, Кеннеди, Холл, 1969; 5 - оригинал Попова, 1974); вм - внутрикристаллическая органическая матрица, лн - линии нарастания, мкс - межпризматические конхиолиновые стенки, мм - межкристаллическая органическая матрица, мпм - межпластинчатая органическая матрица, пвп - призмы второго порядка
Особую структуру (миострак) образуют слои в местах прикрепления мускулов-замыкателей, мускулов ножных и паллиальных (рис. 171). Она построена из видоизмененных арагонитовых кристаллов, которые приобретают форму призм разных размеров, ориентированных нормально к поверхности слоя. Все кристаллы арагонита и кальцита, принимающие участие в образовании стенки раковины, окружены органической оболочкой, толщина которой очень варьирует. Известковая стенка раковины, как было отмечено выше, состоит обычно из трех слоев: наружного, среднего и внутреннего, но у многих можно различить только два слоя. По мнению Тэйлора (1973), исходным типом строения стенки раковины (рис. 171) у всех бивальвий была трехслойная стенка, состоящая из простого призматического слоя (наружного) и двух перламутровых (среднего и внутреннего). Она установлена у моноплакофор и у некоторых групп бивальвий (фоладомиид, унионин и тригониин).
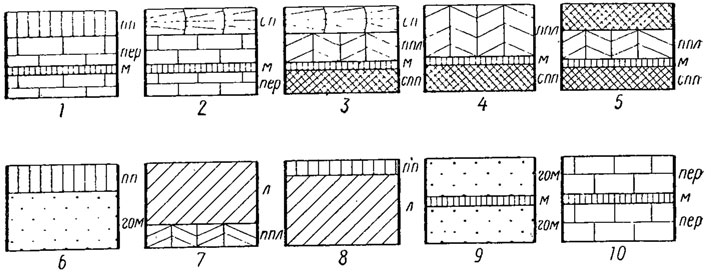
Рис. 171. Микроструктура стенки раковины некоторых групп бивальвий: 1 - Pteriacea, Pinnacea; 2 - Nuculacea; 3 - Lucinacea, Tellinacea, Veneracea; 4 - большинство гетеродонт; 5 - Zirfaea, Myidae; 6 - Solemya; 7 - Pectina- cea, Anomiacea, Limacea; 8 - Ostreacea; 9 - Cuspidariidae, Thraciidae; 10 - Mytilacea; микроструктуры: гом - гомогенная, л - листоватая, м - миострак, пер - перламутровая, пп - простая призматическая, ппл - перекрещенно-пластинчатая, сп - сложнопризматическая, спп - сложно перекрещенно-пластинчатая
Пока еще не установлено таксономическое значение отдельных типов микроструктур, которые наблюдаются в разных группах бивальвий, однако первые наметки уже можно сделать. Призматическая и перламутровая микроструктуры обнаружены у протобранхий, митилид; призматическая - у иноцерамид, гиппуритид; листоватая кальцитовая - у остреин (устриц), некоторых пектенид; гомогенная - у нукулан (протобранхий), хиателл; гранулярная и перекрещенно-пластинчатая - у миид (рис. 171); перекрещенно-пластинчатая простая и сложная - у большинства бивальвий с гетеродонтным замком. В строении раковины в малых количествах участвуют и такие элементы, как стронций, барий, магний. Концентрация элементов зависит от многих факторов и в первую очередь от химического состава, солености и температуры воды.
Скульптура. Поверхность раковины может быть гладкой, с линиями нарастания или скульптированной. Различают два типа скульптуры: радиальную и концентрическую. Радиальная представлена ребрами, расходящимися от макушки к брюшному краю; по размерам различают ребра первого, второго порядков, с широкими или узкими межреберными пространствами. Кроме ребер бывают развиты крупные складки (Lopha, см. рис. 185, 4), отражающиеся на строении всей раковины, а также сильные перегибы, или кили. Обычно имеется один хорошо выраженный киль, отделяющий переднее большое поле от малого заднего (тригонии). Концентрическая скульптура выражена концентрическими ребрами, идущими параллельно линиям нарастания; она отражает ритмическое изменение количества минерального вещества, отлагаемого краем мантии. При пересечении радиальных и концентрических ребер возникает канцеллятная скульптура. Часто на пересечении ребер развиваются бугорки, шипы и различной формы выросты, играющие защитную роль. У некоторых бивальвий сзади и спереди вдоль спинного края обособляются две площадки: передняя - луночка, задняя - щиток, отделенные от остальной поверхности раковины ребрышком или валиком. На щитке обычно расположен лигамент (см. рис. 180, 3).
Наружный слой, или периострак, часто окрашен в различные цвета, причем наиболее яркой и сочной окраской отличаются тропические бивальвии. Окраска иногда пропитывает и внешний известковый слой. У ископаемых бивальвий окраска сохраняется реже, чем у гастропод, тем не менее следы окраски обнаружены в ультрафиолетовых лучах при просмотре раковин девонских, каменноугольных и более молодых моллюсков. При этом также были видны паллиальная линия и отпечатки мускулов.
Внутреннее строение. На внутренней поверхности створок различают структуры, частично отражающие строение тела и расположение некоторых органов. Вдоль утолщенного спинного края протягивается замочная площадка, на которой расположены более или менее развитые зубы, разделенные зубными ямками, в которые входят зубы противоположной створки. Зубы и зубные ямки образуют замок, или замочный аппарат (рис. 172). Замок служит для прочного соединения створок; он корректирует угол раскрывания створок, препятствует их боковому сдвигу при закрытой раковине. Первичный замок, или проринкул, закладывается на стадии продиссоконха; он состоит из ряда небольших первичных зубчиков, которые образуются из радиальных ребер створок; иногда эта стадия первичных зубчиков выпадает из онтогенеза. В дальнейшем, при переходе к стадии диссоконха, на замочном крае образуется утолщенная замочная площадка, на которой закладываются по четыре первичных зубных пластинки; из них затем развиваются различные типы замков, среди которых можно выделить три основных типа: ктенодонтный, прегетеродонтный и гетеродонтный.
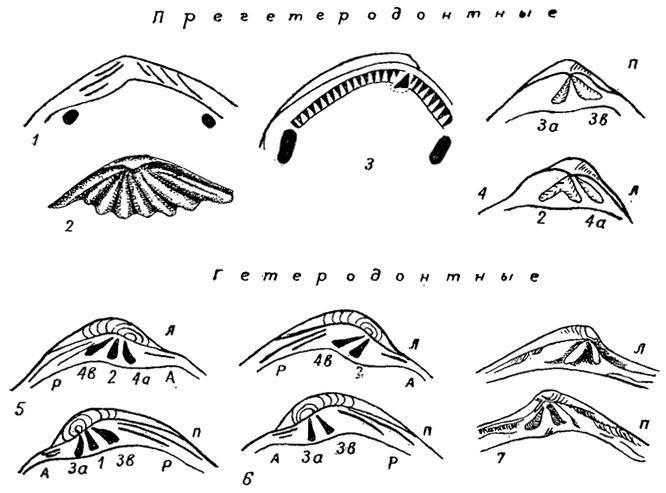
Рис. 172. Некоторые ткпы прегетеродонтных (1, 2, 4), гетеродонтных (5-7) и ктенодонтного (3) замков: 1 - актинодонтный, 2 - лиродесмоидный, 3 - ктенодонтный, 4 - схизодонтный, 5 - циреноидный, 6 - люциноидный, 7 - астартоидный; створки: л - левая, п - правая
Ктенодонтный (гр. ctenion - маленький гребень) замок рассматривается как наиболее примитивный тип замка (характерен для Nuculacea); он состоит из ряда зубов, расположенных по обе стороны от макушки в один прямой или изогнутый ряд. Для замка характерно увеличение числа радиусов по мере роста раковины. Он был ранее известен под названием тассодонтного, или рядозубого (гр. taxis - ряд). Однако подобный тип замка позднее независимо развился у другой группы двустворок (Arcina). Поэтому рекомендовано первичный таксодонтный замок называть ктенодонтным, а вторично возникший - псевдоктенодонтным; развившийся независимо рядозубый замок у некоторых пресноводных бивальвий предлагается называть псевдотаксодонтным (пермский Palaeomutela и современный Pleiodon).
К прегетеродонтному замку (рис. 172, 1, 2, 4) отнесены такие, у которых зубы представляют собой неизмененные или слабо измененные по форме и положению первичные зубные пластинки, радиально расходящиеся от макушки. Различают замки: лиродесмоидный, актинодонтный, схизодонтный, изодонтный, птеринеоидный, циртодонтный, дизодонтный. Лиродесмоидный замок состоит из равных или почти, равных зубов, веерообразно расходящихся от макушки; этот тип замка характерен для ранних бивальвий (Lyrodesma, ордовик). Актинодонтный замок отличается от лиродесмоидного укороченными зубами под макушкой. Схизодонтный замок (гр. schisma - расщеплять) характеризуется развитием зубов, идущих параллельно спинному краю, в левой створке имеет срединный крупный раздвоенный зуб, входящий в крупную зубную ямку правой створки, ограниченную двумя зубами; каждый зуб с боков снабжен насечками. По-видимому, описанный тип замка развился из актинодонтного и независимо возник у морских (тригониины) и пресноводных (унионины) бивальвий. Изодонтный замок (гр. isos - равный); в наиболее развитом виде состоит из двух равных зубов, симметрично расположенных по обе стороны от лигаментной ямки (Spondylus); у пектинид - узкие пластинчатые зубы, от одного до трех, расходящиеся радиально от макушки по обе стороны от лигаментной ямки; эти зубы названы кардинальными, или замочными, крурами (лат. crus, мн. ч. crura - нога). У птеринеоидного замка имеются лишь задние краевые зубы неравной длины. Циртодонтный замок развился из схизодонтного путем редукции примакушечных участков зубов; он характеризуется наличием только краевых зубов, идущих параллельно краю. Дизодонтный замок (гр. префикс dys - означает плохой, неэффективный) состоит из слабо или совеем неразвитых зубов.
Гетеродонтные, или разнозубые, замки (гр. heteros - разный) характеризуются дифференциацией зубов на кардиальные, расположенные под макушкой, и латеральные, лежащие параллельно краям (рис. 172, 5-7); различают передние и задние латеральные; их предлагают обозначать римскими цифрами, а кардинальные зубы - арабскими: четными для левой, нечетными для правой створок. Обычно бывает развито не более трех кардинальных и не более двух латеральных передних и задних зубов. Различают следующие типы замков: астартоидный, циреноидный, люциноидный, кардитоидный, пахиодонтный, десмодонтный, возникших, вероятно, от актинодонтного замка; Астартоидный замок (характерен для рода Astarte) состоит из хорошо развитых кардинальных зубов и переднего бокового; циреноидный, или корбикулоидный, замок отличается присутствием в каждой створке трех кардинальных зубов и парных передних и задних латеральных, причем на левой створке нижний задний латеральный зуб обычно не развит; люциноидный отличается наличием двух кардинальных и таких же, как в корбикулоидном, латеральных; кардитоидный отличается удлиненными косо направленными кардинальными зубами, отсутствием латеральных. Пахиодонтный, или толстозубый замок (гр. pachys - толстый) состоит из двух-трех толстых неправильной формы зубов, возникших из кардинальных или, возможно, латеральных. Десмодонтный замок отличается редукцией или полным исчезновением зубов у форм, ведущих зарывающийся образ жизни; и значительным развитием внутренней связки, расположенной иногда на особом выступе - хондрофоре. Подобно право- и левозавернутым раковинам гастропод у бивальвий иногда наблюдается так называемая инверсия замка, при которой зубы, свойственные левой створке, развиваются на правой, и наоборот. Это явление наблюдается у некоторых морских семейств (Astartidae, Crassatellidae) и некоторых пресноводных (Unionidae) и приводит к осложнениям при цифровых обозначениях зубов.
Лигамент, иди связка, расположенный вдоль спинного края, служит для соединения створок и для их открывания, когда ослабевают мускулы-замыкатели (рис. 173). Лигамент представляет собой упругий эластичный тяж, состоящий из периострака, наружного пластинчатого и внутреннего волокнистого слоев. Пластинчатый слой слагается из параллельно расположенных пластинок протеина, а волокнистый имеет волокнистую структуру, с фибрами, ориентированными длинной осью к поверхности роста в местах их образования. Конхиолин, из которого строится лигамент, состоит в основном из протеинов; волокнистый слой может частично обызвествляться с образованием плотной оси (литодесмы). К трем описанным слоям иногда еще добавляется четвертый - соединительный слой, развитый между периостраком и пластинчатым слоем. Связка, расположенная по обе стороны от макушки вдоль замочного края, называется, амфидетной (гр. amphi - двойной, с обеих сторон; detic - связанный); лежащая позади макушки, иногда на особой подпорке (нимфе) - опистодетной (гр. opisthen - задний). Отношение толщины лигамента к его длине может составлять 1 : 10. У большинства прикрепленных цементацией бивальвий (устриц) связка расположена между створками в особых ямках на связочной площадке, и здесь отношение длины лигамента в передне-заднем направлении к толщине в дорзо-вентральном направлении составляет 3 : 1 или 4 : 1. У форм, имеющих ктенодонтный или псевдоктенодонтный замок, обычно под макушкой развита площадка, или а рея, на которой к бороздкам, сходящимся под углом к макушке (шевронам), прикрепляется амфидетная связка. У пектинид лигамент разделен на две части - пластинчатый слой расположен вдоль ушек, а волокнистый - в особой ямке под макушкой. В сложной, или мультивинкулярной, связке (лат. multus - много, vinculum - тяж), развитой у иноцерамид и бакевеллид, волокнистый слой распадается на ряд частей - резилий, каждая из которых лежит в отдельной ямке на связочной площадке; резилии соединяются между собой пластинчатым слоем (рис. 173). У ряда бивальвий, ведущих сверлящий образ жизни, связка редуцируется или полностью исчезает; ее функцию принимает на себя передний мускул (см. рис. 181, 26).
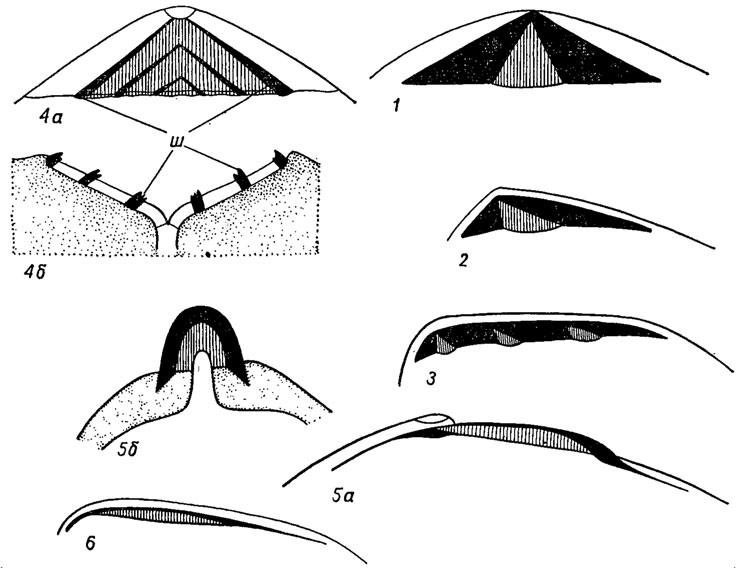
Рис. 173. Типы связок: 1-2 - амфидетная; 3-4 - сложная связка: 3 - у Isognomon, 4а, б - у Area; 5-6 - опистодетная: 5а, б - у Cardium, 6 - у Mytilus; зачернен пластинчатый слой связки, штриховка - волокнистый слой, точки - раковинный слой; ш - шевроны
На внутренней поверхности створок сохраняются места прикрепления разнообразных мускулов, оставляющие на раковине отпечатки (рис. 174). К ним относятся мускулы-замыкатели (аддукторы), мантийные, или паллиальные, мускулы, мускулы ноги и некоторые другие. Мускулы-замыкатели, два или один, служат для быстрого закрывания створок и действуют как антагонисты лигамента, стремящегося в силу своей эластичности открыть створки. В этом антагонизме заключено большое приспособительное значение подобного взаимодействия. В случае опасности моллюску необходимо быстро захлопнуть створки; после того как опасность прошла, можно понемногу ослабить напряжение мускулов и приоткрыть створки. После гибели моллюска и разрушения мягкого тела створки под действием лигамента приоткрываются. В зависимости от числа и размеров мускулов различают двустворок с двумя равными или неравными мускулами или с одним крупным задним мускулом. Бивальвии с почти равными мускулами называются изомиарными, гомомиарными, или равномускульными. У рудистов мускулы прикрепляются к особым подставкам,, называемым миофорами. Обычно оба аддуктора действуют одновременно, но у сверлящих моллюсков (фоладины) передний вынесен на отворот переднего края створки и его действия противоположны действию заднего мускула, лежащего внутри створок; при сжимании переднего мускула расширяется задний край (у фоладин раковина зияющая); при сокращении заднего мускула расширяется переднее отверстие.
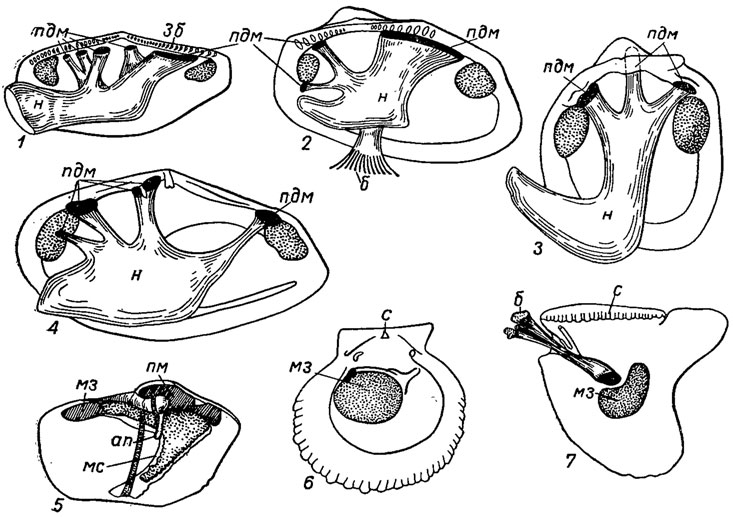
Рис. 174. Схема расположения мускулов-замыкателей (мз), ноги и педальных мускулов (пдм): 1 - Yoldia, 2 - Area, 3 - Cardium, 4 - Tellina, 5 - Zirfaea 6 - Chlamys, 7 - Isognomon; ап - апофиз, б - биссус, зб - зубы, мс - мантийный синус, н - нога, пм - передний мускул, с - связка
У большой группы бивальвий - мономиарий, или одномускульных, имеется один крупный задний мускул, расположенный почти в центре створки и обычно оставляющий большой отпечаток. Между изомиариями и мономиариями существует ряд двустворок, обладающих двумя мускулами разных размеров - гетеромиарии, и между ними можно найти все переходы; при этом наблюдается постепенное уменьшение переднего мускула и усиление и смещение к центру заднего. Каждый мускул состоит из двух типов пучков волокон; один пучок "стекловидный" (быстрый) служит для быстрого закрывания створок, а другой - "перламутровый", длительно удерживает створки в сомкнутом состоянии.
Мантия прикрепляется на некотором расстоянии от края к внутренней поверхности раковины с помощью мышечных волокон, назначение которых состоит в быстром втягивании края мантии, внутрь раковины перед ее закрытием. Внутри раковины эти мускулы образуют мантийную, или паллиальную, линию: у изомиарий она начинается у переднего мускула и идет почти параллельно краю до заднего аддуктора; у мономиарий мантийная линия следует параллельно краю раковины. У форм, обладающих сифонами, последние полностью или частично втягиваются внутрь раковины при помощи особых сифональных мускулов-ретракторов, оставляющих след прикрепления в задней части раковины в виде выемки - мантийного, или паллиального, синуса. Бивальвий с мантийным синусом называют синупаллиатными, без него - интегрипаллиатными.
Как было сказано выше, нога выполняет разнообразные функции и у большинства бивальвий довольно подвижна. Она приводится в движение с помощью системы мышц, среди которых различают парные; передние и задние ретракторы - мускулы, втягивающие ногу в раковину, мускулы-протракторы - способствующие выдвижению ноги из раковины. У форм, обладающих биссусом, задний ретрактор превращается в сильно развитую мышцу, которая выдвигается вперед и иногда распадается на несколько пучков. Все мышцы одним концом прикрепляются к раковине, другим - входят в основание ноги. Отпечатки этих мускулов Можно видеть вдоль спинного края и около мускулов-ретракторов (рис. 174). У форм мало или совсем неподвижных мускулатура ноги сильно упрощена или редуцирована. Очень редко сохраняются мускулы, прикрепляющие висцеральный мешок к раковине (у некоторых тригоний, унионин), и жаберные мышцы-ретракторы (у некоторых устриц и пектинид).
Основы систематики и классификация. До последних лет единой общепринятой систематики двустворчатых моллюсков не было. Палеонтологи использовали для выделения отрядов особенности строения замка, лигамента и аддукторов, но, как оказалось, близкие типы замков могут конвергентно возникать в разных эволюционных стволах, часто далеко отстоящих друг от друга. Зоологи для выделения отрядов использовали строение внутренних органов и в первую очередь жабр. В последние годы были предприняты попытки выделить более высокие категории, такие, как подклассы или надотряды. В американских "Основах палеонтологии" (1969) класс бивальвий разделен на 6 подклассов: Palaeotaxodonta, Cryptodonta, Pteriomorpha, Palaeoheterodonta, Heterodonta, Anomalodesmata. Советские малакологи (Невесская, Старобогатов, Скарлато и Эберзин, 1971) предложили для выделения надотрядов и отрядов использовать анатомические и конхиологические данные с учетом экологической дифференциации. Они предложили разделять всех бивальвий на три надотряда, в пределах которых строение жаберного аппарата определяет возможности адаптивной специализации: Protobranchia, Autobranchia и Septibranchia.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'