
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Общие положения тафономии
И. А. Ефремов [361] под тафономией понимал учение о закономерностях перехода органических остатков из биосферы в литосферу в результате совокупности биологических и геологических процессов. В тафономии рассматриваются, с одной стороны, причины массовой гибели организмов, перенос, разрушение и распределение их остатков в осадке, диагенетические изменения остатков в земной коре и другие явления, которые, как правило, лежат вне поля зрения палеонтолога при его обычных (морфологических, филогенетических и др.) исследованиях, а с другой стороны - процессы формирования и распределения современных, в первую очередь биогенных осадков, образования и изменения породы, что является предметом изучения седиментолога и литолога. Таким образом, для тафономии характерен комплексный подход исследований, основанный на биологических, палеонтологических, палеоэкологических, гидробиологических, седиментологических, литологических, геохимических и других данных. Она как бы объединяет биостратономию Й. Вайгельта и учение о фоссилизации В. Дееке.
Как следует из определения тафономии, главной ее проблемой является изучение процессов захоронения остатков организмов в биосфере и восстановление условий формирования их местонахождений в литосфере. Так как процесс перехода посмертных остатков организмов из биосферы в литосферу происходит в результате взаимодействия биологических и геологических явлений, а превращение захороненных остатков в окаменелости протекает под воздействием геохимического и других факторов, то можно представить всю сложность решения главной проблемы тафономии, слагающейся из целого ряда задач: 1) изучения условий и характера посмертного разрушения, переноса, распределения и захоронения остатков современных организмов в различных обстановках; 2) восстановления характера динамической переработки и сохранения остатков организмов в процессе их фоссилизации в земной коре и 3) изучения условий изменения окаменелостей в зоне поверхностного выветривания. Важными задачами также являются: совершенствование как общей, так и частных методик тафономического анализа; разработка тафономической классификации местонахождений и упорядочение терминологии.
Основной единицей тафономической классификации служит "захоронение" - "слой или часть слоя с остатками растений или животных" [520, с. 70].
Весь сложный путь остатков от момента гибели организмов в биосфере через их захоронение и фоссилизацию в литосфере и до момента их разрушения вновь в биосфере можно назвать тафономическим циклом [1243]. В его объеме выделяются четыре главных этапа, причем первые три являются основными в едином процессе образования местонахождений. На каждом этапе остатки организмов подвергаются воздействию определенных процессов, причем большинство из них действует не только в течение одного этапа, но начинает оказывать влияние в предшествующем или продолжает свою "работу" на последующем этапе, что обусловлено взаимосвязанностью процессов.
Тафономический цикл - образование, сохранение и разрушение местонахождения - может быть полностью завершен только при определенных условиях.
1. Этапы должны последовательно сменять друг друга (особенно важна последовательность первых трех этапов); выпадение хотя бы одного этапа ведет к нарушению всего цикла.
2. Смена трех этапов должна быть геологически одновременной. Если скопления остатков организмов, сформировавшиеся в биосфере (I этап), не будут быстро захоронены в осадке (II этап), то они неминуемо подвергнутся полному разрушению химическим, биохимическим или механическим путем. Если погребенные остатки не будут быстро фоссилизированы (III этап), то химические и физические процессы приведут их также к полному уничтожению.
3. Остатки организмов должны находиться в земной коре длительное время. Промежуток времени между III и IV этапами должен быть как можно большим, чтобы остатки, заключенные в толще породы, подверглись полной фоссилизации, т. е. превратились в окаменелости.
4. На IV этапе местонахождение должно быть выведено на дневную поверхность в результате горообразования и размыва покрывающих толщ. Небольшая выборка окаменелостей из местонахождения может быть также получена с керном при бурений скважин как на суше, так и в море.
5. На каждом этапе образования местонахождений в природе должны иметь место благоприятные стечения обстоятельств (совпадения, по И. А. Ефремову), как в биосфере, так и в литосфере, обусловленные первичными или вторичными факторами.
Большое значение для понимания как всего хода формирования местонахождений, так и процессов, действующих на определенных этапах тафономического цикла, имеет разработанное И. А. Ефремовым положение о выборочности. Оно основано на том, что каждый этап общего цикла образования местонахождения обладает своими решетками, под которыми понимается комплекс факторов внешней среды, воздействующих на остатки организмов или благоприятно (сохраняя и пропуская их на следующую ступень преобразования) или неблагоприятно (уничтожая их на каком-либо из этапов). На каждом этапе доминирующую роль играет определенный комплекс факторов.
Первый этап - подготовка исходного материала в биосфере. Решающая роль принадлежит здесь биологическим факторам, в первую очередь составу, количеству, динамике среды, распределению и плотности населения, массовой гибели организмов, первичной концентрации посмертных остатков, характеру химического и минерального состава организмов. Существенную роль в уничтожении скоплений посмертных остатков организмов на данном этапе играют также факторы биохимический (разложение органики) и химический (растворение минеральных скелетов), которые идут, как правило, под контролем климатического фактора.
Второй этап - перераспределение исходного комплекса остатков, его аккумуляция и захоронение в осадке - характеризуется наиболее сложным спектром разнообразных факторов. Доминирующую роль начинают играть физические, особенно механические факторы, обусловливающие перенос и переработку исходных остатков. Продолжают действовать биохимический и химический факторы. Вступают в цикл также и геологические факторы (тип бассейна, скорость и характер осадконакопления, состав и структура отложений и т. д.).
Третий этап - фоссилизация захороненных остатков. На этом этапе на первый план выдвигаются физические и геохимические факторы, приводящие к диагенетическим превращениям вещества остатков под воздействием больших давлений и высоких температур в связи с погружением на большие глубины. Степень преобразования остатков здесь зависит от тектонических условий, скорости литификации осадка, степени минерализации иловых и поровых вод и т. д. На переработку первичного органического материала на данном этапе продолжает также оказывать большое влияние биохимический фактор. Непременным условием для данного этапа является длительность воздействия всей совокупности процессов, поэтому время может рассматриваться здесь как один из главных факторов образования местонахождений.
Последний, четвертый этап - изменение и разрушение окаменелостей в зоне гипергенеза. На первое место вновь выходят такие факторы, как химический (растворение и замещение) и физический (механическая деструкция окаменелостей).
В результате неоднократного воздействия на исходные посмертные остатки организмов всех перечисленных факторов практически от начала до конца тафономического цикла происходит постепенное и устойчивое сокращение их общей массы, сопровождающееся изменением химического и минерального состава остатков, искажением количественных и качественных соотношений организмов, установившихся в прижизненном палеосообществе на соответствующем участке суши или моря.
Нарушение последовательности этапов тафономического цикла, отсутствие того или иного благоприятного условия или совпадения на каждом этапе, выборочность деструкции, захоронения и фоссилизации - все это в совокупности и определяет неполноту геологической летописи.
Для обозначения скопления живых и мертвых организмов и их фрагментов, места их жизни, смерти и погребения были введены многочисленные термины. Тем обзор 1243]. Скопление мертвых организмов Э. Васмунд [2455] назвал танатоценозом, а место их смерти - танатотопом. Аллохтонные танатоценозы, т. е. скопления организмов или их частей, перемещенные к месту захоронения, В. Квенштедт [2114] назвал тафоценозом. Одновременно появилось эквивалентное понятие некроценоза - ископаемого болотного биоценоза [2324]. В. А. Красилов [520] считает тафоценозом вообще совокупность остатков, содержащихся в захоронении, не уточняя их алло- или автохтонности. Скопление окаменелостей в слое предложено называть ориктоценозом [295]. Этот термин широко вошел в литературу. Его палеоботанический вариант - фитоориктоценоз - М. В. Ошуркова [792, с. 4] определяет как "совокупность фоссилизированных микро- и макроскопических остатков растений, захороненных в определенных фациальных условиях и приуроченных к конкретной литологической разности пород".
В. А. Собецкий [972] предложил для скоплений ископаемых другой термин - ориктокомплекс, поскольку термин "ценоз" подразумевает жизненно важные связи внутри современных сообществ организмов. По тем же соображениям он ввел термин "танатомасса" вместо термина "танатоценоз". А. Мюллер [1982] выделил два типа ископаемых танатоценозов: автохтонный, образованный из некроценотического танатоценоза, и аллохтонный, образованный за счет тафоценотического танатоценоза. Псевдоценозом названы скопления продуктов специфического цикла жизнедеятельности, например, панцирей членистоногих, продуктов линьки, листьев и пыльцы [1920, 1921]. Несколько раньше Л. Ш. Давиташвили [295] называл подобные остатки мероценозом. Ихноценоз - это сообщество следов жизнедеятельности [1876]. Х. Бёгер [1354] предложил называть тафоценозом аллохтонный танатоценоз. Этим же термином он обозначает переотложенные остатки организмов. Автохтонные танатоценозы он, как В. В. Друщиц и В. В. Меннер [347], называет некроценозами.
Говоря о тафоценозах, ориктоценозах и других типах ассоциаций ископаемых в захоронении, палеонтологи обычно не оговаривают, а часто может быть и не осознают важного обстоятельства: они имеют дело не со всеми организмами, имеющимися в захоронении, а лишь с выборкой. Чаще всего эта выборка таксономическая. Специалист по тетраподам изучает ассоциацию позвоночных, пропуская остатки растений и остракод, палеоботаник имеет в виду лишь растения и т. д. Нередко основанием выбора остатков для тафономического изучения служит их размер (например, раздельно изучается микро- и макрофоссилии) или техника их извлечения (мацерация породы в определенной жидкости, сепарирование по плотности, отбор остатков прямо на обнажении).
Сколько-нибудь полного анализа всех органических остатков, имеющихся в захоронении, почти никогда не делают. Предпринимались попытки комплексного изучения отдельных местонахождений, но и в этих случаях многие группы остатков выпадали из рассмотрения (например, остатки бактерий и грибов).
Тафономические объекты можно разбить на два класса. С одной стороны, это ископаемые остатки, давно захороненные в породах и являющиеся объектами любых иных палеонтологических исследований. С другой стороны, это современные или недавно жившие организмы, попадающие в захоронения на наших глазах или захороненные недавно, так что обстановка захоронения не успела измениться настолько, чтобы нуждаться в специальной реконструкции. Изучением таких захоронений занимается актуопалеонтология. Разновидностью ее можно считать экспериментальную тафономию, т. е. изучение процессов фоссилизации и деструкции, формирования захоронений на специально подобранных, порой даже искусственных объектах.
При изучении захоронений палеонтологу (актуопалеонтологу) приходится заниматься тафономией индивидов и тафономией сообществ (сравни деление экологии на аут- и синэкологию).
Тафономия индивидов. Изучает обстоятельства смерти, фоссилизации отдельных частей и деструкции остатков. Эти исследования в большей или меньшей мере выполняются палеонтологами при обычных морфологических и таксономических исследованиях, когда они реконструируют недостающие части, воссоздают прижизненный облик индивида, пытаются выяснить, можно ли считать данный признак прижизненным или обусловленным дефектами сохранности.
Обстоятельства смерти организмов важно знать для аут- и синэкологических реконструкций. Иногда на остатках организмов наблюдаются явные признаки старения. У раковинных морских беспозвоночных (моллюсков, брахиопод) к старости нарушается периодичность в нарастании раковины, резко меняется скульптура и прочие детали. Имеются документы смерти от хищников. Так, в полостях желудков акул и ихтиозавров находили остатки белемнитов.
Большие скопления озерных животных на отдельных плоскостях напластования считаются свидетельствами заморов. Резкие изменения температуры и солености вод ведут к массовой смерти обитателей.
Так, суровой зимой 1962-1963 гг. у северных берегов Западной Европы произошло катастрофическое вымирание литоральных и мелководных сублиторальных животных Последствия этого вымирания чувствовались 5-6 лет [1259]. Известна высокая смертность микропланктона теплых течений при встрече их с холодными водами высоких широт.
Показательна история неогеновой фауны Средиземного моря. В периоды океанизации (отложения илов) пелагиаль населял типично океанический планктон. В моменты изоляции, обмеления и осолонения (отложение эвапоритов) океаническая фауна быстро вымирала, сменялась галофильными моллюсками [1757]. Массовая катастрофическая гибель донной фауны на ограниченной площади может произойти из-за извержения вулкана. Так, после извержения на о. Десепшен (Южно-Шотландские о-ва) донные животные в бухте Порт Фостер систематически не обнаруживались дночерпателями и драгами. Гибель животных была вызвана накоплением 30-сантиметрового слоя пепла на дне, повышением температуры и наличием ядовитых газов [1280]. Смертность животных резко увеличивается при падении содержания кислорода в воде (anoxic events).
Процессам фоссилизации посвящена большая литература, и среди нее обзоры [1243, 2235]. Особенно детально они изучались у ископаемых растений, для которых знание типа фоссилизации иногда необходимо для принятия правильного таксономического решения (см. 4.1). Тип фоссилизации определяет и выбор исследовательской техники, например, изготовление пришлифовок, шлифов, микротомных срезов, получение слепков с полостей и др.; см. подробнее [683, 864].
Процессы фоссилизации изучаются путем сравнения остатков, испытавших разные степени преобразования исходных структур, при этом широко используются сведения о постседиментационных преобразованиях осадочных пород (рис. 105, 106). В некоторых случаях проводят эксперименты с искусственной фоссилизацией организмов. Например, Т. М. Гаррис [1708] поджаривал современные семена и затем их мацерировал, чтобы установить устойчивость отдельных оболочек к мацерации в зависимости от степени обугливания. Экспериментальное окремнение сине-зеленых водорослей оказалось важным для правильной интерпретации структуры микрофоссилий в докембрийских строматолитовых кремнях [1603]. Оказалось, что при окремнении слизистые чехлы имитируют клеточные оболочки, а содержимое клеток коллапсирует и имитирует ядро. Эксперименты выполнялись и на животном материале [1718, 2076].
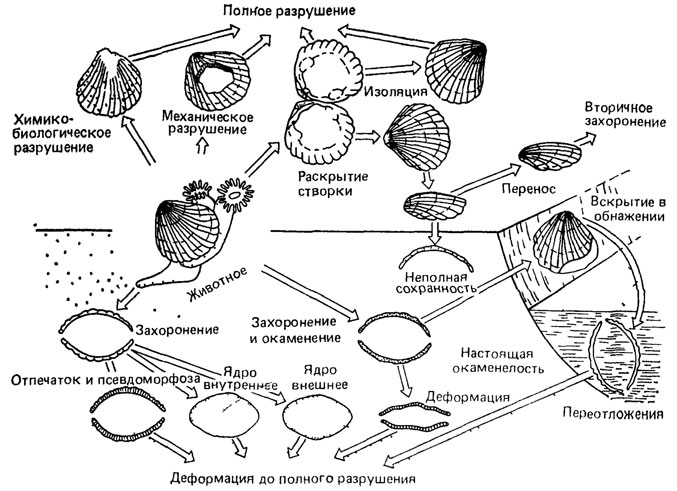
Рис. 105. Варианты послесмертных превращений раковины двустворчатого моллюска. По Э. Тениус из А. М. Циглера
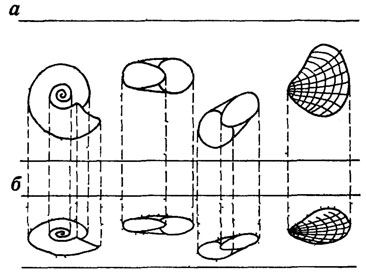
Рис. 106. Уплотнение породы (а) и вызванная этим деформация (б) окаменелостей. По А. М. Циглеру
Процесс, противоположный фоссилизации, можно назвать деструкцией. Она может быть биологической (деятельность бактерий, грибов, многоклеточных консументов), химической (растворение остатков) и механической (дезинтеграция остатков физическими факторами). С изначальной неполнотой при захоронении не следует путать неполноту при современной деструкции местонахождений. К разрушению остатков могут приводить и древние эрозионные срезы [1044]. Актуалистические и биостратономические наблюдения позволили наметить стадии распадения организмов. Например, для млекопитающих установлена такая последовательность распада [2390, 2464]: первоначально обычно отделяется нижняя челюсть - "закон нижней челюсти", затем череп, шейные и хвостовые позвонки, большинство фаланг, верхние и нижние части конечностей, ребра, позвоночный столб. Иные последовательности распада описаны у птиц, рыб и других животных. На распад влияют многие факторы, меняющие характер последовательности. Это климатические условия, посмертное перемещение остатков, деятельность хищников и трупоедов, гидродинамические условия на месте погребения и др. Обычно хорошая сохранность связана с быстрой седиментацией, но иногда и с медленной, если имела место ранняя консервация в сингенетических конкрециях на границе вода - осадок.
Морская среда редко бывает химически нейтральной по отношению к остаткам беспозвоночных. Пустые раковины растворяются как на дне, так и после погребения в мягком осадке, поскольку иловые воды нередко агрессивны по отношению к карбонату. Особенно легко растворяются карбонатные скелеты в пересыщенных углекислотой холодных водах. Наиболее стойки к растворению кремневые, фосфатные и богатые органическими компонентами (хитином, тектином, спонгином) скелеты беспозвоночных. Растворению подвергаются и окаменелости (иногда на их месте образуются пустоты). Экспериментальные исследования показали избирательность механического разрушения беспозвоночных (гастропод, кораллов, морских ежей, мшанок) и известковых водорослей. Деструкция растений при захоронении изучена более систематически, но лишь в отношении таких скоплений растительных остатков, которые дают начало торфу (а затем и углю), и применительно к деструкции тканей. Для растений в большей мере, чем для животных, характерно прижизненное отделение большого количества частей (листьев, семян, пыльцы, коры и др.). Совместное захоронение частей учитывается при реконструкции прижизненного сочетания частей и общего облика растений (см. подробнее 1.4).
Тафономия сообществ. Изучает не сохранность отдельных индивидов или их дисперсных частей, а обнаруженные в породе ассоциации ископаемых, классифицирует эти ассоциации по разным признакам, выявляет пути их образования.
Нами уже рассматривались некоторые понятия, относящиеся к тафономии сообществ. То, что при тафономическом исследовании мы имеем дело не с полными совокупностями фоссилий, данного захоронения, а с некоторыми выборками, не учитывается при классификации захоронений. Определенные типы захоронений всегда вводятся лишь для той выборки фоссилий, с которой имеет дело данный исследователь. Например, когда говорится о том, что сообщество рифовых известняков, состоящее из скелетов мшанок, кораллов и водорослей, автохтонное, то просто не учитывается присутствие в тех же породах микрофоссилий, принадлежащих планктонным организмам. Наоборот, совокупность костей и фрагментарных растительных остатков в русловых песчаниках может описываться как аллохтонная, хотя в том же слое могут встречаться не подвергшиеся переносу раковины двустворок и бактериальная флора, развившаяся в осадке после его накопления.
По-видимому, полностью аллохтонных или автохтонных захоронений вообще не бывает. Об аллохтонности и автохтонности можно говорить лишь в отношении интересующих нас организмов. Поэтому пока нельзя говорить о классификации захоронений как таковых. Мы просто не располагаем достаточным количеством таких местонахождений и захоронений в них, которые изучены с извлечением всего доступного комплекса фоссилий. Поэтому говоря далее о классификации захоронений, мы будем иметь в виду лишь те компоненты захоронений, которые захватываются традиционными методами сбора фоссилий.
Даже при таком ограничении классификации захоронений - очень трудная задача. Захоронения могут классифицироваться по разным основаниям. Можно учитывать таксономическую принадлежность фоссилий и выделять захоронения диатомей, фораминифер, двустворок, позвоночных, высших растений и т. д. Иногда в захоронении встречаются определенные морфологические компоненты организмов, так что получаются захоронения зубов акул, листьев (например, листовые кровли), древесных стволов. При этом используются термины, относящиеся к размерности фоссилий (комплексы микрофауны или миоспор). Учитываются количественные пропорции таксонов в захоронениях (моно-, олиго- и полидоминантные захоронения).
Наибольшее значение в классификации захоронений обычно придается транспортировке и сортировке остатков. Именно такой смысл придается перечисленным выше типам сообществ - танатоценозу, тафоценозу и др. Требующиеся для этого реконструкции обстановок, в которых формировались захоронения, всегда страдают гипотетичностью. В общем случае те классификации, которые требуют сложных реконструкций, менее объективны и им должны предшествовать более простые и более объективные классификации, тесно связанные с непосредственными наблюдениями. Однако было бы ошибкой требовать строгого разграничения наблюдаемых и реконструируемых признаков захоронений и соответственно структурных и генетических классификаций [520, 825, 1151]. Дело в том, что сама классификация захоронений по признакам, считающимся непосредственно наблюдаемыми, - окатанность обломков, частота встречаемости таксонов и другие, требует введения оценок, которые могут быть весьма субъективными. Например, при сравнении окатанности и фрагментированности остатков в разных захоронениях важно учитывать потенциальную устойчивость данных частей к деструкции. Ясно, что надо сравнивать части, близкие по такой устойчивости, а оценка этой близости неизбежно субъективна. С другой стороны, можно назвать очень сложные экосистемы, легко распознаваемые непосредственно на обнажении (устричная банка, кораллово-мшанковый риф и т. п.).
Учитывая возможность классификации захоронений по разным основаниям, общая классификация захоронений, которая пока не создана, должна быть многомерной. Её придется строить так, чтобы каждое конкретное захоронение попадало одновременно в разные классификационные ячейки, как это и принято в тафономических исследованиях. Например, известное Павлодарское местонахождение гиппариновой фауны характеризуется одновременно как аллохтонное, аллювиальное и полидоминантное. Каждая из этих характеристик одновременно является классификационной ячейкой.
В тафономической литературе разные группы критериев часто смешиваются и подменяют друг друга. Например, Г. П. Радченко [851] выделял шесть типов тафоценозов, отличающихся по удаленности захоронения от материнского фитоценоза и по участию в захоронении компонентов разных фитоценозов. При этом он допускал возможность распознавать по виду остатков, откуда они принесены - с верховьев речной долины или со склона близлежащей возвышенности. О несостоятельности этой и подобных классификаций уже шла речь в литературе [520, 643].
О том, как конкретно организуется исследование тафономии сообществ, писали многие исследователи; см. подробнее [1243]. Самая общая программа исследования разработана И. А. Ефремовым [362] преимущественно для захоронений наземных позвоночных. В целом в основе исследования лежат полевые наблюдения с последующей обработкой материала, широким привлечением данных фациального анализа, литологии, петрографии, палеоэкологии и других дисциплин. Палеоэкология нередко оказывается частью тафономических исследований, прежде чем получает от них интересующие ее данные. Существенным моментом при интерпретации материала является выбор актуалистической модели.
Изучение начинается с выяснения позиции захоронения в разрезе в стратиграфическом и фациальном отношениях. Детально описывается нужная часть разреза, вплоть до выявления мелких специфических фаций. Если органические остатки встречаются на большой площади, то используются различные методы опробования [973, 2386]. Полноценное изучение крупного захоронения нуждается в площадных раскопках. Опубликовано немало инструкций по их техническому осуществлению, особенно применительно к позвоночным [683, 1849 и др.]. Необходимо массовое извлечение всего комплекса органических остатков и следов жизнедеятельности, вплоть до микроскопических объектов, их строгая регистрация и фиксация в разрезе и на площади. С этой целью в палеонтологию целесообразно внедрять богатый опыт археологических исследований [1821, 1907, 2105]. При обширных раскопках важно предварительное снятие плана вскрытой площади, точная привязка к нему материала. На плане отражаются пространственные изменения мощности и литологии слоев, сохранность остатков, их ориентировка, соотношение со структурами и текстурами вмещающих пород Выявлению биостратономических закономерностей служат разрезы по направлениям, наиболее информативным в биостратономическом отношении.
Поскольку раскопки разрушают слой с органическими остатками, производится систематическое фотографирование и крупно масштабные зарисовки всех необходимых объектов (рис. 107, 108), снятие реплик с седиментологических структур, отбираются образцы на различные анализы, позволяющие детально охарактеризовать слой по площади. Разобранная порода иногда промывается и просеивается Дополнительная биостратономическая информация собирается в камеральный период путем анализа остатков в монолитах, фотографирования в различного рода лучах [882].
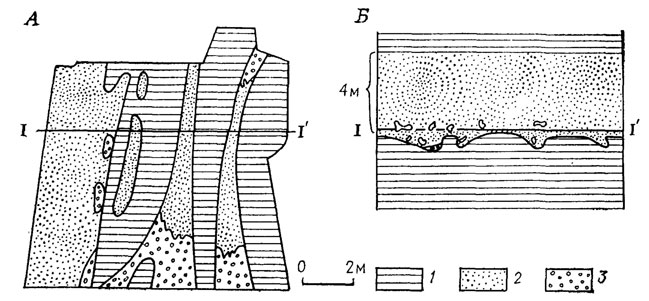
Рис. 107. План (А) и разрез (Б) местонахождения Букобай со среднетриасовыми позвоночными в букобайской свите Соль-Илецкого района Оренбургской области. По В. Г. Очеву. 1 - глина; 2 - костеносный песчаник, 3 - костеносный базальный конгломерат; I-I′ - линия сечения
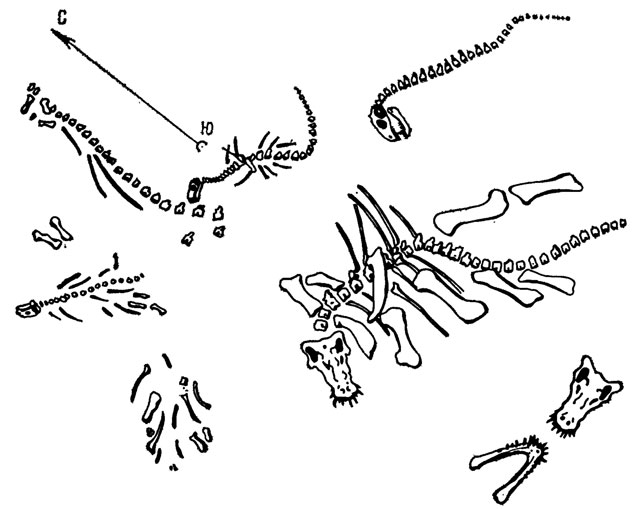
Рис. 108. Изображение в крупном плане положения скелетов зверообразных пресмыкающихся в местонахождении у д. Ежово в Пермской области. По П. К. Чудинову
Тафономическое исследование подразумевает изучение особенностей распределения органических остатков в слое (рассеянное, гнездами и т. д.), их ориентировку, сохранность, сортированность (рис. 109, 110) Например, Е. А. Иванова [421] выделила несколько типов распределения в разрезе морской каменноугольной фауны (прослоями, линзами, равномерное, выделены и более дробные подтипы). И. А Ефремов [362] в местонахождениях континентальных позвоночных различает два типа распределения в разрезе (послойное и перемешанное) и три типа распределения по площади (рассеянное, концентрированное и промежуточное). Другие авторы классифицируют форму скоплений органических остатков [283, 973], характер их упаковки.

Рис. 109. Танатоценозы раковин двустворчатых моллюсков в осадках мелководной части раннемелового моря на р. Боярке (Хатангская впадина). По В. А. Захарову, Ю. Г. Юдовному. а - автохтонный танатоценоз - групповое скопление целых раковин бухий, ув. 1/2, б - автохтонный танатоценоз - рассеянные по слою целые раковины и отдельные створки пектинид (борейонектесов, ув. 1/16, в - аллохтонный танатоценоз - ракушниковые скопления створок и раковин пектинид (борейонектесов)
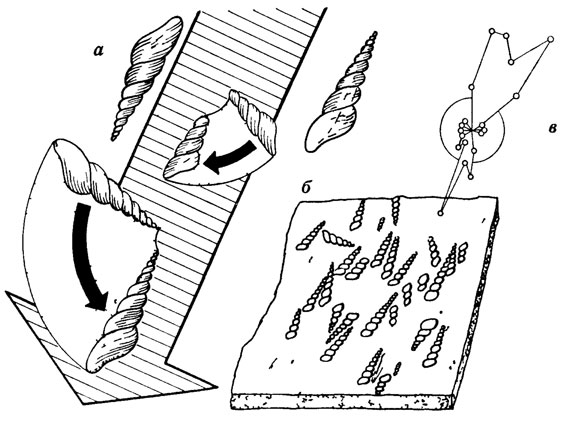
Рис. 110. Послесмертная ориентировка конусовидных раковин на примере брюхоногого Turritella из олигоцена Баварии. По А. Зейлахеру из А. М. Циглера. а - схема механизма ориентировки течением (зачерненные стрелы), б - часть плитки с Turritella, в - векторная диаграмма для 76 экземпляров
В отношении ориентировки ископаемых хороший пример классификации был дан в 1931 г. Р. Рихтером [1984]. Он различает (А) перевертывание (Einkippung), которое связано с движением вокруг более или менее горизонтальной оси и определяет гравитационную ориентировку, и (Б) выруливание (Einstenezung), которое связано с вращением вокруг вертикальной оси и приводит к азимутальной ориентировке. Перевертывание бывает свободным (под влиянием силы тяжести и собственной формы при погружении в водной среде или жидком осадке) и заторможенным (под влиянием приложенных со стороны механических сил). Азимутальная ориентировка бывает автономной, зависящей от сопротивления среды, которое обусловлено формой тела, и заякоренной. Выделяются и более дробные типы. Существует огромная литература по ориентировке остатков [868, 1737, 2263 и др.].
О сохранности фоссилий уже шла речь в разделе, посвященном тафономии индивидов. Тафономия сообществ интересуется не столько тем, как сохранились отдельные экземпляры организмов, сколько совокупной сохранностью всех фоссилий захоронения, степенью сохранности организмов, принадлежащих разным таксонам. С сохранностью фоссилий отчасти связана последующая характеристика захоронения, его сортировка, или выборочность. Так, И. А. Ефремов [362] назвал закономерное обеднение комплексов организмов в ходе захоронения. Факторы, определяющие выборочность, могут быть биологическими ("закон больших чисел" [360]; выборочная гибель), физико-географическими (выборочность при транспортировке к месту захоронения), геохимическими (выборочность при фоссилизации). При этом происходит как потеря [2361], так и искажение информации [522] об исходном сообществе организмов. Изучают формы выборочности по размерам, форме остатков, их механической прочности и устойчивости к растворению. Так вскрываются существенные процессы формирования местонахождений (гидродинамические условия, мера аллохтонности тафоценоза и др.), собирается материал для расшифровки выборочности по систематическому и экологическому составу реконструируемого сообщества.
Одна из главных задач тафономии - установление аллохтонности или автохтонности фоссилий, т. е. выяснение, подверглись ли фоссилий транспортировке до их захоронения. Часто роль посмертной транспортировки организмов переоценивается. Захоронение на месте жизни допускается как результат катастрофических событий, например извержение вулкана. Захоронение в прижизненном положении допускается для инфауны при аномально высоких темпах осадконакопления, застойной гидродинамике. Обычно прижизненное захоронение беспозвоночных в рифовых постройках.
Практика палеоэкологических исследований свидетельствует о гораздо более широком диапазоне условий, способствующих прижизненному захоронению. Такие захоронения описаны почти для всех типов животных. Вероятность прижизненного положения захороненных эпифаунных неприкрепленных или мягко прикрепленных форм, конечно, очень мала. Эти организмы часто захороняются после незначительной транспортировки в пределах места обитания. Тогда ископаемая популяция будет существенно близка прижизненной. В литературе приводились критерии автохтонности захороненных популяций [222, 225, 226, 392, 395, 1241, 1243, 1360, 1570 и др.].
Транспортировка остатков подробно рассматривалась и в палеоботанической литературе. В палеоботанике, как и при изучении планктонных и нектонных организмов, довольно трудно сформулировать сами представления об аллохтонности и автохтонности. Опадающие части растений, как и скелеты организмов, живущих в толще воды, до захоронения испытывают перемещение (падают в водоем, опускаются на дно), но могут и не разноситься по сторонам. Такие захоронения иногда называют гипавтохтонными. Для реконструкции экосистем гипавтохтонные и автохтонные остатки часто можно рассматривать совместно.
Если фоссилии и их ассоциации не несут прямых указаний на транспортировку, то отличить автохтонные, гипавтохтонные и аллохтонные остатки можно, лишь изучив образ жизни соответствующих организмов. Для этого нужна актуалистическая модель, выбор которой можно оспаривать. Например, для многих палеозойских фораминифер нельзя указать точные современные аналоги, а поэтому невозможно сказать, были ли данные фораминиферы бентосными, планктонными или псевдопланктонными. Не существует надежных критериев для опознания заведомо сухопутных и водных слоевищных и травянистых растений. Соответственно нельзя, например, сказать, были ли остатки многих палеозойских и мезозойских печеночников транспортированы с суши или захоронились в месте произрастания, а если они были водными, то плавающими или донными.
Нет необходимости пояснять, что перечисленные и все прочие компоненты тафономического исследования нуждаются в большем или меньшем использовании актуопалеонтологических и вообще актуалистических исследований. Правда, следует заметить, что актуалистические исследования нередко указывают на такие процессы, которые едва ли когда-нибудь удается расшифровать на палеонтологическом материале. В качестве примера можно привести весьма экзотический способ сортировки - селективное уничтожение морскими птицами левых створок Spisula solida [1412].
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'