
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Характеристика известковых водорослей, типы обызвествления слоевища
Обызвествление водорослей и возникновение раковинных животных организмов представляет собой одну из грандиозных загадок биогеохимии. На этот счет высказано много гипотез, среди которых явное предпочтение отдается скачкообразному изменению факторов внешней среды: состава атмосферы и (или) морской воды /Томмотский ярус, 1969; Tappan, 1974; Riding, 1984/. Возможно, именно эти обстоятельства и привели к тому, что представители всех отделов водорослей вблизи границы докембрия и кембрия приобрели возможность строить карбонатный скелет. Однако это не объясняет, почему некоторые из них (в частности синезеленые) впоследствии утратили эту способность (рис. I. 1), тогда как другие - зеленые и особенно красные - усилили. Следует иметь в виду, что соотношение водорослей, способных к обызвествлению слоевища и не обладающих такой возможностью, среди ископаемых водорослей разных отделов определить очень сложно. Тем более что это соотношение явно, и притом весьма значительно, изменяется, как это и было показано на примере синезеленых водорослей. Соответственно значительно изменялась во времени и породообразующая роль известковых водорослей. Для палеозоя эти данные недавно суммированы /Chuvashov, Riding, 1984/.
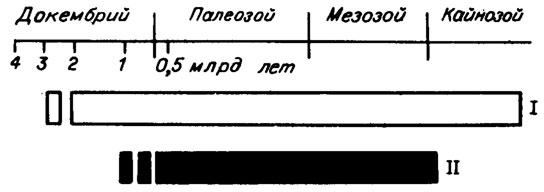
Рис. I. 1. Время существования синезеленых водорослей (I) и синезеленых водорослей с обызвествленным слоевищем (II) (по R. Riding, /1982/)
Разные отделы известковых водорослей обладают принципиально разными способами обызвествления. У синезеленых водорослей обызвествление представляет собой тонкую пленку, обволакивающую слоевище. Поверхностным, но более глубоким является обызвествление зеленых водорослей, причем у дазикладациевых внешние окончания ветвей и центральная часть слоевища обычно не обызвествлены. Кодиевые имеют более полно и равномерно обызвествленные талломы. Слоевища красных водорослей, если они подвергались обызвествлению, сохраняются в ископаемом состоянии почти полностью.
Таким образом, понятие "известковые водоросли" ни в коей мере не являются систематическим термином, а отражает только способность некоторых водорослевых слоевищ к частичному или полному обызвествлению. Эта искусственная группировка водорослей чрезвычайно важна для палеонтологии и геологии по ряду причин. Прежде всего, известковые водоросли наилучшим образом и чаще всего сохраняются в ископаемом состоянии, и подавляющей части информации по ископаемой альгофлоре мы обязаны именно им. Во-вторых, эти организмы-породообразователи являются и наиболее точными индикаторами фациальных обстановок.
Процессы биоминерализации вообще и у водорослей в частности изучены весьма слабо, хотя этими вопросами занимались многие исследователи. Итоги наблюдений относительно водорослей суммированы в ряде работ В. П. Маслова /1961 а, 1973 и др./.
При так называемом органическом осаждении извести карбонат кальция выделяется из клеточного сока на стенках клетки. Выделенный минеральный осадок состоит из мелких закономерно расположенных кристалликов кальцита, которые при хорошей сохранности дают в скрещенных николях волнистое угасание. Способностью органического осаждения извести обладают многие группы животных и красные водоросли семейства Corallinaceae.
Физиологическое отложение обусловлено усвоением углекислоты в процессе фотосинтеза ассимилирующими частями водорослей. Карбонат кальция выпадает на внешней поверхности этих частей слоевища в виде неправильно расположенных кристалликов кальцита. Такой процесс обызвествления, видимо, характерен для зеленых и некоторых групп синезеленых водорослей. Таким же образом обызвествляются вегетативные части харовых водорослей, тогда как репродуктивные органы харовых водорослей кальцитизируются путем " органического" обызвествления. Ооспорангии ископаемых и современных харовых водорослей имеют при хорошей сохранности слоистую текстуру стенки, в которой наблюдается многократное чередование микрослоев, образованных то "чистым" кальцитом с концентрически слоистой или радиально-лучистой структурой, то темным, обогащенным органикой. Образование ооспорангия начинается с относительно мощной органической пленки, которая всегда или почти всегда сохраняется в ископаемом состоянии, в то время как промежуточные микрослои темного, обогащенного органическим веществом кальцита часто преобразуются в процессе диагенеза с образованием одного мощного слоя радиально-лучистого кальцита (рис. I. 2). Органическая пленка явно стимулирует образование микрослоя "чистого" кальцита, но только до определенной толщины. Для наращивания следующей пленки "чистого" кальцита вновь необходимо образование карбонатно-органической темной пленки.

Рис. I. 2. Основные типы строения известкового чехла водорослей. 1 - обызвествленные части слоевища сложены темным пелитоморфным карбонатом: а - Girvanella, б - Epiphyton, в - Koivaella, г - Renalcis, д - Tubiphytes, e - Parachaetetes; 2 - стекловатый прозрачный кальцит: а - Devonoscale, б - Kamaena, в - Donezella, г - Claracrusta; 3 - двухслойные карбонатные чехлы: а - Wetheredella, б - Sphaeroporella, в - Asphaltina, г - Nuia; 4 - многослойные чехлы, сложенные чередованием слоев темного пелитоморфного и светлого прозрачного кальцита: а - первичная структура утрикулов харовых водорослей, б - вторичная структура; 5 - известковые чехлы, сложенные мелкозернистым кальцитом: Hedstroemia, Garw odia и т. д. в продольных (а) и поперечных (б) сечениях
Третий тип отложения карбонатного кальцита называют биохимическим. Водоросли в данном случае играют косвенную роль, изменяя рН среды в тонком слое воды, примыкающем к слоевищу, что способствует осаждению извести.
Следует упомянуть также о смешанном процессе осаждения извести при большом влиянии водорослей. Водорослевые пленки и нити механически, биохимически и химически связывают осаждающийся карбонат кальция. При отмирании органических тканей водорослей и уплотнении осадившегося карбоната образуется его корочка. Последний (смешанный) тип обызвествления характерен для строматолитов.
Разные способы обызвествления слоевища, толщина обызвествленной части, последовательность процесса, а также, вероятно, последующие изменения обызвествленной части в ископаемом сочетании определяют большое разнообразие типов строения известковых чехлов. Кальцит, фиксированный разными группами ископаемых известковых водорослей, можно по структурным особенностям разделить на несколько типов (см. рис. I. 2).
1. Темный пелитоморфный карбонат, фарфорово-белый в отраженном свете и бархатисто-черный в проходящем. Такой тип кальцита характерен для наиболее древних водорослей, относимых к синезеленым, и может быть прослежен с позднего докембрия. Это трубчатые (Girvanella, Obruchevella, Tubomorphophyton, Koivaella и др.), нитчатые (Epiphyton), пузырчатые (Renalcis, Chabakovia), массивные и палочковидные (Jkella, Tubiphytes) слоевища водорослей. Подобный пелитоморфный карбонат образует стенки клеток красных водорослей (Solenopora, Parachaetetes, Katavella), микрослой в стенках репродуктивных органов харовых водорослей. Есть основания считать, что это наиболее древний, "первичный" тип обызвествления.
2. Прозрачный или желтоватый кальцит слагает непористые чехлы слоевищ родов Proninella, Devonoscale и др.; пористые чехлы зеленых водорослей семейств Palaeoberesellidae, Beresellidae и др.; стенки клеток красных водорослей (Solenopora, Paralancicula).
3. Тонковолокнистый радиально-лучистый кальцит входит в состав многослойных известковых чехлов. Так, водоросли родов Wetheredella, Asphaltina имеют двухслойные чехлы, причем верхний слой образован пелитоморфным карбонатом, а внутренний, более мощный, слагается радиально-лучистым пористым кальцитом. При трехслойном чехле (Nuia) срединный слой представлен темным пелитоморфным карбонатом, а внешний и внутренний образованы волокнистым желтовато-прозрачным кальцитом.
4. К этому типу примыкает строение скорлупы репродуктивных органов харовых водорослей при одном существенном различии - стенки репродуктивных органов образованы многократным чередованием светлых и темных слоев кальцита, причем те и другие имели первоначально концентрически-слоистую текстуру. При вторичных изменениях "чистый" кальцит наиболее легко приобретает радиально-лучистое строение. В следующую стадию вторичных преобразований вся скорлупа органа, кроме внутреннего слоя, теряет свое слоистое строение; образуется единый мощный радиально-лучистый слой (см. рис. I. 2).
5. Мелкокристаллический серый в проходящем свете карбонат, образованный мелкими изометричными, по-разному ориентированными кристалликами кальцита. Такой карбонат слагает известковые чехлы водорослей родов. Garwoodia, Hedstroemia, Uraimella и др., а также вегетативные части харовых водорослей.
Из сказанного следует, что водоросли, относимые сейчас к одному систематическому типу, могут иметь известковые чехлы с разным характером обызвествления. Это обстоятельство имеет два объяснения: либо водоросли одного типа (отдела) могут менять способы обызвествления своего чехла, либо из-за несовершенства систематики мы объединяем в одном таксоне разные группы. При использовании тонких методик изучения вещества и его структуры можно получить интересные данные относительно систематики и эволюции ископаемых водорослей.
Намеченные выше типы обызвествления свойственны не только водорослям. Хорошие аналогии наблюдаются прежде всего у фораминифер. Карбонат первого типа слагает огромное множество раковин фораминифер, в том числе и наиболее древних примитивных - Archaesphaera, Parathuramminet, Earlandia, Tikhinella и др. Прозрачный желтоватый кальцит слагает стенки фораминифер родов Nanicella, Protonodosaria и др. Двухслойные стенки, сложенные тонкой пленкой черного карбоната и мощным слоем радиально-лучистого кальцита, обращенного или внутрь (Nodosaria, Geinitvzina), или наружу (Multiseptida) раковины, также характеризуют большое число родов фораминифер разного возраста. Мелкокристаллический серый карбонат свойствен многим родам. Наиболее характерный пример - турнейские турнейлиды.
Следовательно, структурные типы обызвествления отражают какие-то общие биохимические процессы, происходящие в клетках как животных, так и растительных организмов.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'