
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Размножение красных водорослей
Сложные процессы размножения красных водорослей резко выделяют этот отдел водорослей среди других представителей альгофлоры. Багрянки обладают разными формами бесполого размножения, сложным строением половой системы, многообразием циклов развития. На этих признаках строится классификация современных представителей отдела.
Вегетативное размножение происходит в результате роста дополнительных побегов, которые берут начало от подошвы слоевища или от его стелющихся ветвей. Оно свойственно представителям обоих классов красных водорослей. Примитивные представители класса бангиевых размножаются только вегетативно. У одноклеточных и колониальных форм бангиевых водорослей при вегетативном размножении клетки делятся на дочерние (две или более).
Бесполое размножение. У низкоорганизованных багрянок бесполое размножение осуществляется моноспорами. Таким образом размножаются почти все бангиевые и редкие представители флоридиевых. Моноспоры - споры, которые образуются в результате превращения всего протопласта клетки в одну спору. Моноспоры не имеют жгутика и оболочки, передвижение их после выхода из клетки осуществляется за счет амебовидных движений. У бангиевых моноспоры могут образоваться в любой клетке слоевища, которая по форме не отличается от других вегетативных клеток. В классе флоридиевых моноспорангии могут образовываться на веточках ограниченного роста. У водорослей, слоевище которых образовано обособленными нитями, в моноспорангии превращаются конечные клетки боковых ветвей (см. рис. IV. 1). У водорослей с дифференцированным слоевищем моноспорангии образуются из внешних клеток ассимиляционных нитей. Моноспорангии отличаются от вегетативных клеток яйце- или шаровидной формой. Голые моноспоры выходят через отверстие в вершине созревшего спорангия, быстро одеваются оболочкой и начинают прорастать.
Однако более характерны для флоридиевых тетраспоры, которые образуются по четыре в клетке, называемой по этому признаку тетраспорангием. По способу деления клетки различаются три типа тетраспорангиев: разделенные крестообразно, зонально и тетраэдрически (см. рис. IV. 1). В крестообразно разделенных спорангиях вся клетка делится, пополам поперечной перегородкой, а затем каждая из двух образовавшихся клеток делится еще раз продольными перегородками, которые располагаются или в одной плоскости, или под прямым углом одна к другой (см. рис. IV. 1). При зональном делении все перегородки проходят параллельно одна другой и тетраспоры располагаются в один ряд (см. рис. IV. 1, В). При тетраэдрическом делении перегородки располагаются косо по отношению друг к другу (см. рис. IV. 1, Б). Способ деления тетраспорангиев - постоянный признак для определенных таксонов. Для кораллиновых водорослей характерны так называемые биспоры, образующиеся при разделении спорангия только на две части.
Для систематики багрянок используется и такой признак, как положение тетраспорангиев в слоевище. Тетраспоры обычно образуются в конечных клетках веточек ограниченного роста. Положение тетраспорангиев между клеточными нитями (интеркалярная позиция) встречается сравнительно редко (см. рис. IV. 1, Г).
Половое размножение характерно для всех представителей флоридиевых, но только наиболее организованные бангиевые обладают этим способом воспроизводства. Половой процесс багрянок оогамный. Мужские и женские гаметы лишены жгутиков; при оплодотворении женские остаются на растении (гаметофите), а мужские (спермацеи) выбрасываются в воду. Спермацеи - шаро- или яйцевидные клетки, бесцветные, без хлоропластов. Спермацеи образуются в клетках, функционирующих как мужские органы размножения.
У некоторых багрянок сперматангии расположены в слоевище беспорядочно. У большей части водорослей они собраны в более или менее обширные группы - сорусы. Нитевидные багрянки имеют сорусы в виде густых пучков иногда шаровидной формы. Водоросли с массивным слоевищем обладают сорусами в виде подушковидных образований на поверхности слоевища (рис. IV. 2). У кораллиновых сорусы расположены в специальных углублениях или пустотах - концептакулах. Иногда наружные коровые клетки дают начало однорядным, параллельно расположенным нитям, выступающим над поверхностью слоевища. Клетки этих нитей затем отчленяют множественные сперматангии. Иногда сперматангии образуются на специальных коротких веточках - стихидиях, или трихобластах (см. рис. IV. 2).
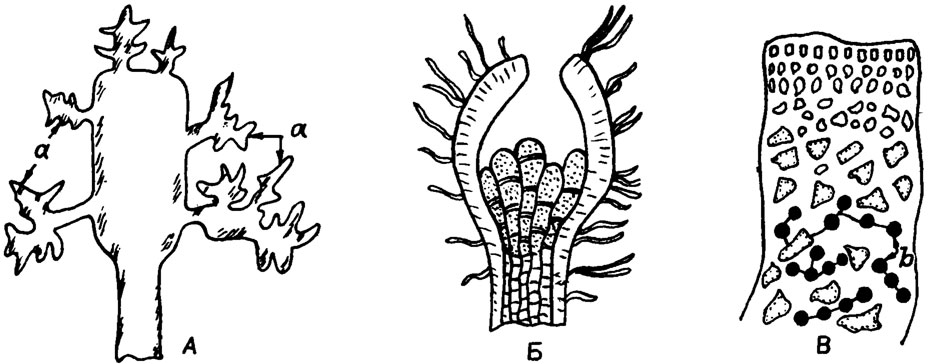
Рис. IV. 2. Тетраспорангии некоторых красных водорослей (по К. Л. Виноградовой /Жизнь растений, 1977/). А - расположение тетраспорангиев (а) на выростах вокруг цилиндрического слоевища у Geiidium; Б - концептакулы с тетраспорангиями у Corallina; В - сгруппированные в цепочки интеркалярные тетраспорангии (в) у Iridiae
Женские репродуктивные органы красных водорослей (оогонии) называют карпогоном, которой представляет собой клетку, заполненную цитоплазмой без хлоропластов. Верхняя часть этой клетки преобразована в длинный трубчатый вырост (трихогон), при помощи которого улавливаются спермацеи. Форма выроста варьирует; он может быть коротким кеглевидным, длинным узким, спирально закрученным. У водорослей с нитчатым слоевищем карпогон располагается открыто.
Карпогон после оплодотворения спермацеями превращается в зиготу, которая начинает прорастать, развиваясь в новое растение, принадлежащее уже к бесполому поколению - спорофиту.
У красных водорослей зигота после сложных превращений преобразуется в особые споры - карпоспоры - голые, лишенные жгутиков клетки, способные к амебовидным перемещениям. Только перед началом прорастания карпоспоры приобретают оболочки, образуются они в карпоспорангиях.
Развитие зиготы происходит неодинаково у разных багрянок. У бангиевых карпоспорангии делятся и образуется от 4 до 32 карпоспор. У флоридиевых после оплодотворения карпогона развиваются специальные нити гонимобласта. Конечные клетки этих нитей превращаются в карпоспорангии.
У большинства флоридиевых оплодотворенный карпогон сливается с особыми (ауксилярными) клетками, богатыми питательными веществами. Только после этого процесса развивается гонимобласт. Зрелый гонимобласт с карпоспорами называют цистокарпом. Место его расположения, форма, характер оболочки, способ выхода карпоспор - важные признаки, используемые в таксономии современных красных водорослей на уровне родов.
Следовательно, в ходе жизненного цикла наиболее высокоорганизованных багрянок наблюдается чередование полового и бесполого размножения, но последовательность их часто нарушается. Гаметофит и спорофит могут повторяться каждый по нескольку раз. У большинства багрянок чередуются тетраспорофит, гаметофит и карпоспорофит, причем разные формы могут давать слоевища, заметно отличающиеся по морфологии.
Большинство альгологов считают, что первичный цикл развития красных водорослей состоял из смены трех морфологически одинаковых форм, которые существовали независимо. В дальнейшем сформировались гетероморфные циклы развития, при которых гаметофит и спорофит приобрели существенные морфологические различия. По другим представлениям, карпофит - более позднее эволюционное приобретение, а на ранних стадиях существования красных водорослей существовали только два поколения.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'