
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Морфология слоевищ красных водорослей
Клетки и клеточные нити. Основным элементом строения слоевища красных водорослей является клетка. В. П. Маслов /1962/ различает сферическую, бочковидную, цилиндрическую, параллелепипедальную, многоугольную, серповидную или в виде колпачка, конусовидную формы клеток.
Сферические чрезвычайно редко встречаются у известковых багряных водорослей. Бочковидные имеются в гипоталлии и в свободных нитях у мелабезиевых. Цилиндрические распознаются с трудом, так как в продольных сечениях они дают прямоугольники, как и параллелепипедальные клетки. Для определения формы таких клеток необходимо иметь два сечения слоевища. Параллелепипедальные наиболее часто встречаются у каменистых багрянок. Более правильно, может быть, говорить о призматических клетках, так как в поперечных сечениях часто встречаются не только четырехугольники, но и разнообразные по числу граней многоугольники. У большинства соленопор, у лысьвелл и других клетки всего слоевища или его части призматические. Колпачковидные встречаются на концах растущих нитей. Конусовидные наиболее характерны для нитей базального гипоталлия. Многоугольные (в любом допустимом сечении) встречаются в случае сильного в процессе роста "сдавливания" клеток во всех направлениях. Наилучшим примером клеток подобного рода являются клетки периталлия у водоросли рода Lysvaella (табл. XXII фиг. 1-3; см. рис. IV. 5).
Кроме того, в коре некоторых ископаемых багрянок встречаются яйцевидные клетки. Они иногда приострены на апикальном конце и завершаются короткими волосками (табл. XXII, фиг. 4). Этот тип клеток В. П. Маслов не различал.
Существует обширная группа водорослей, у которых клеточное строение настолько трансформируется за счет исчезновения одних стенок клеток и видоизменения других, что клеточные нити или устанавливаются с большим трудом (см. рис. IV. 6, Ungdarella, Komia), или не улавливаются (см. рис. IV. 4, IV. 8).
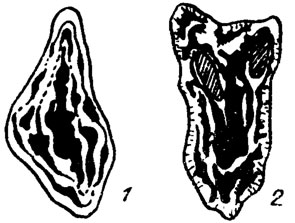
Рис. IV. 8. Схема строения слоевища с сильно видоизмененным обликом периталлия и гипоталлия родов. 1 - Pseudostacheoides; 2 - Epistacheoides (по В. Mamet, A. Roux /1977/)
Такое аномальное строение большой группы организмов, сближенных с красными водорослями, у ряда исследователей вызывает сомнение в их водорослевой природе. Мы имеем в виду организмы типа Stacheoides, Parastacheoides и др.
Слоевище редко сложено клетками одной формы. Сравнительно однообразны по форме клетки слоевища соленопорацей, что же касается других багрянок, особенно с субцилиндрическим слоевищем и хорошо развитым срединным гипоталлием, то здесь формы клеток очень разнообразны. Например, у лысьвелл (табл. XXII, фиг. 1-4) различаются узкие призматические клетки срединного гипоталлия, многоугольные клетки периталлия и яйцевидные с волосками клетки коры.
Даже в пределах единой клеточной нити форма клеток изменяется. Так, например, у катавелл (табл. XX, фиг. 1, 4) клеточные нити сложены низкими широкими клетками, но верхушечные клетки нитей становятся высокими и относительно широкими. Именно они определяют рост слоевища, отчленяя вниз клетку за клеткой (табл. XX, фиг. 4). Подобные же клетки являются и органами размножения. Равным образом меняется форма клеток в единой нити у монилипорелл (табл. XVIII, фиг. 1-5). Во внутренней части периталлия клетки здесь тесно прижаты и являются призматическими по форме, в периферийной части, где клеточные нити расположены рыхло, клетки становятся субцилиндрическими.
В основе слоевища любой формы лежит клеточная нить. Простая клеточная нить может в единственном числе представлять собой слоевище багрянки, которое чаще образовано группой клеточных нитей, обладающих, как правило, морфологическими различиями в разных частях слоевища, что определяется их функциональным назначением. Сближенные нити клеток образуют сложную ткань, напоминающую строение современных высших растений. Выше упоминалось, что форма клеток в значительной степени зависит от того, насколько тесно они взаимодействуют с соседними клетками. Все это остается, естественно, в силе и по отношению к нитям клеток. При наличии тесно сжатых многих нитей появляются новые признаки. Поперечные перегородки, отделяющие клетки нитей, располагаются на разных уровнях - обособляются нити клеток. В случае расположения перегородок на одном уровне образуется "решетчатая структура" - различаются нити и ряды (рис. IV. 9, Б). При слиянии поперечных перегородок в единую линию образуется серия клеток (рис. IV. 10, IV. 11).
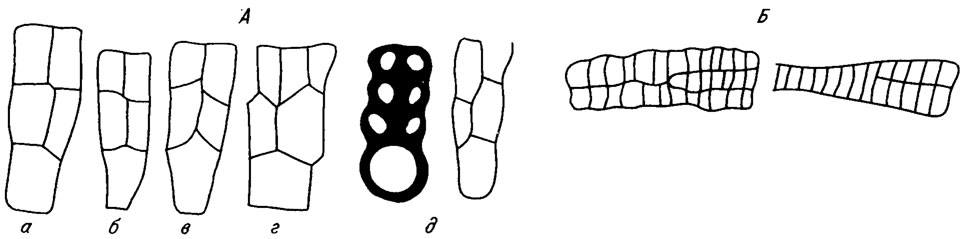
Рис. IV. 9. Способы дихотомического деления нитей (А) и увеличения количества серий в слоевище (Б) (по В. П. Маслову /1962/). а - Lithothamnium; б - Paleothamnium; в - Melobesia; Lithophyllum; г - Karpathia
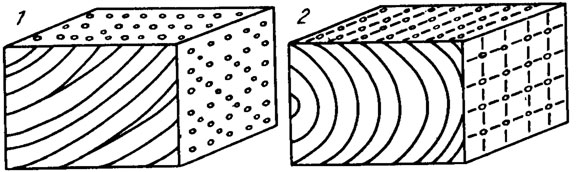
Рис. IV. 10. Строение слоевища некоторых кораллиновых красных водорослей /Poignant, 1979 a, b/. Базальный гипоталлий при расположении клеток в нитях (1) и рядах (2)
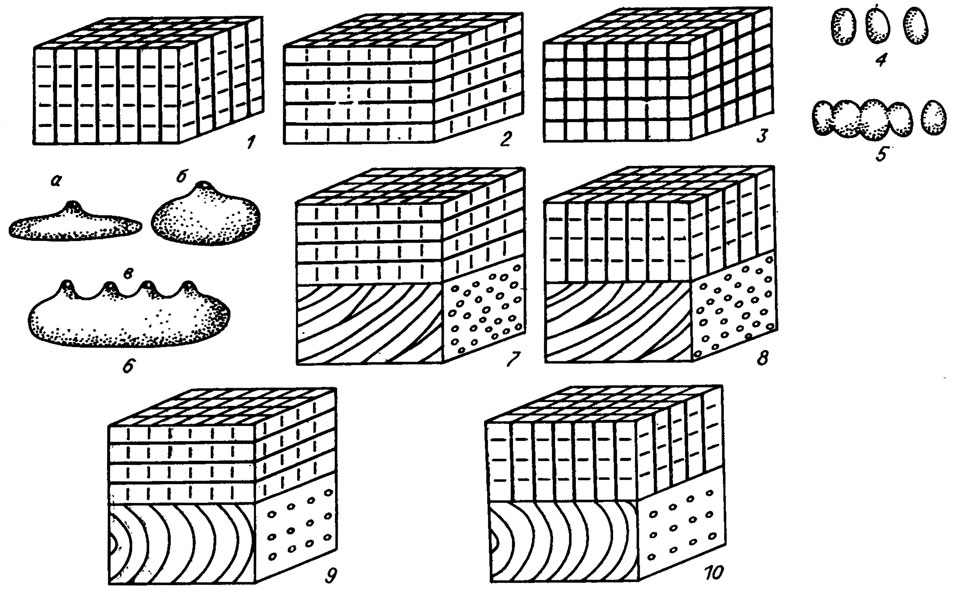
Рис. IV. 11. Схема строениям 1-3 - периталлия некоторых красных водорослей (по A. F. Poignant /1979 а, b/) при расположении клеток в нитях (1), рядах (2), рядах и сериях (3); 4-6 - репродуктивных органов кораллиновых водорослей; 4 - изолированные спорангии; 5 - изолированные спорангии и сорус; 6 - концептакул (а - моноперфорированный сильно уплощенный; б - моноперфорированный, в - полиперфорированный); 7-10) - слоевищ некоторых кораллиновых водорослей (7 - Archaelithothamnium; 8 - Lithothamnium; 9 - Mesophyllum; 10 - Lithophyllum)
Дихотомическое ветвление нитей осуществляется при продольном делении клетки. При этом рядом с материнской клеткой появляется тонкая клиновидная дочерняя, в другом случае появляются две меньшие дочерние клетки (рис. IV. 9, А). Очень часто верхушечная клетка, располагающаяся в основании дихотомически разделившейся нити, резко увеличивается в размерах и изменяет свою форму (см. рис. IV. 9, А).
Существуют слоевища, состоящие из клеточных серий и клеточных нитей. Первая из них развивается делением клеток в поперечном и продольном направлениях. При этом образуются горизонтальные или дуговидные ряды клеток, в которых нити не прослеживаются. При "решетчатой ткани" выражены одинаково хорошо и клеточные нити, и серии (см. рис. IV. 10; IV. 11).
В дифференцированном слоевище красных водорослей различают базальную ткань, или гипоталлий, и периферическую - периталлий.
Клетки нитей, располагающихся в базальной части прикрепленного слоевища, заметно отличаются по толщине стенок, форме, их взаимному расположению от клеток более молодой части таллома. Часто нити в базальной части слоевища имеют и другое направление роста. Такие видоизмененные клеточные нити в базальной части слоевища называют базальным гипоталлием. У палеозойских форм базальный гипоталлий обычно выражен слабо, но у мезо-кайнозойских багрянок обособляется достаточно отчетливо (см. рис. IV. 10; IV. 11).
Довольно редкие среди красных водорослей формы с пластинчатым слоевищем, например Archaelithophyllum (рис. IV. 12), имеют четкое деление на гипоталлий и периталлий. Близкие к ним морфологически Principia имеют очень слабо развитый (редуцированный) периталлий.

Рис. IV. 12. Соотношение гипоталлия (слева) и периталлия в слоевище Archaelithophyllum, ув. 100 (по J. Johnson /1960/)
Красные водоросли, обладающие прямостоящим субцилиндрическим слоевищем, часто имеют осевой, или срединный, гипоталлий, образованный одиночной нитью (см. рис. IV. 7), небольшой группой нитей или значительным по диаметру их пучком (см. рис. IV. 5, IV. 13).
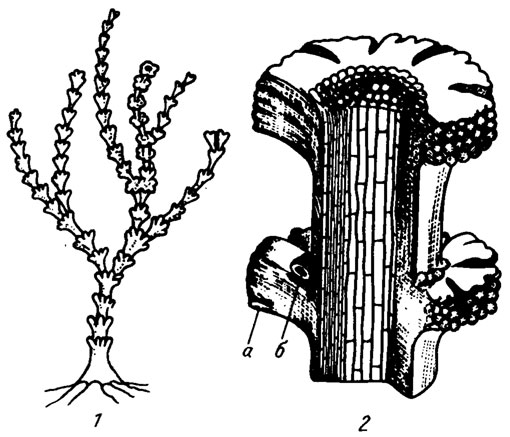
Рис. IV. 13. Реконструкция водоросли Paralancicula fibrosa Shuysky (1973). 1 - внешний облик слоевища, 2 - увеличенный фрагмент слоевища в разрезе, видны нити крупных клеток срединного гипоталлия и более мелкие клетки периталлия: а - грибовидные кольцевые расширения, б - более слабо развитые кольцевые расширения второго яруса. Реконструкция В. П. Шуйского
Срединный гипоталлий слагается клеточными нитями, морфологически отличными от остальной части слоевища. Как уже упоминалось выше, клетки срединного гипоталлия иногда почти не обызвествлены (см. рис. IV. 5) или обызвествлены значительно слабее клеток периталлия.
В очерке о морфологии современных багрянок уже говорилось, что в периферийной части слоевищ может обособляться более или менее мощный слой клеток, образующих так называемый коровый слой, или кору. Срединному и базальному гипоталлию противопоставляется остальная часть слоевища - периталлий (см. рис. IV. 5).
Органы размножения. Выше мы кратко описали сложные процессы размножения современных красных водорослей, многообразие существующих у них органов размножения. Наши познания в этой области об ископаемых багрянках чрезвычайно неполны.
У нитчатых красных водорослей типа budlovia и Tubomarphophyton /Корде, 1973/ органы размножения в виде мелких шарообразных тел располагаются на апикальных концах веток или при дихотомическом делении одна из веточек преобразуется в спорангий.
В слоевищах водорослей семейства Solenoporaceae давно были подмечены крупные овальные полости, которые обычно рассматриваются как органы размножения - концептакулы (рис. IV. 14, А, Б, В).
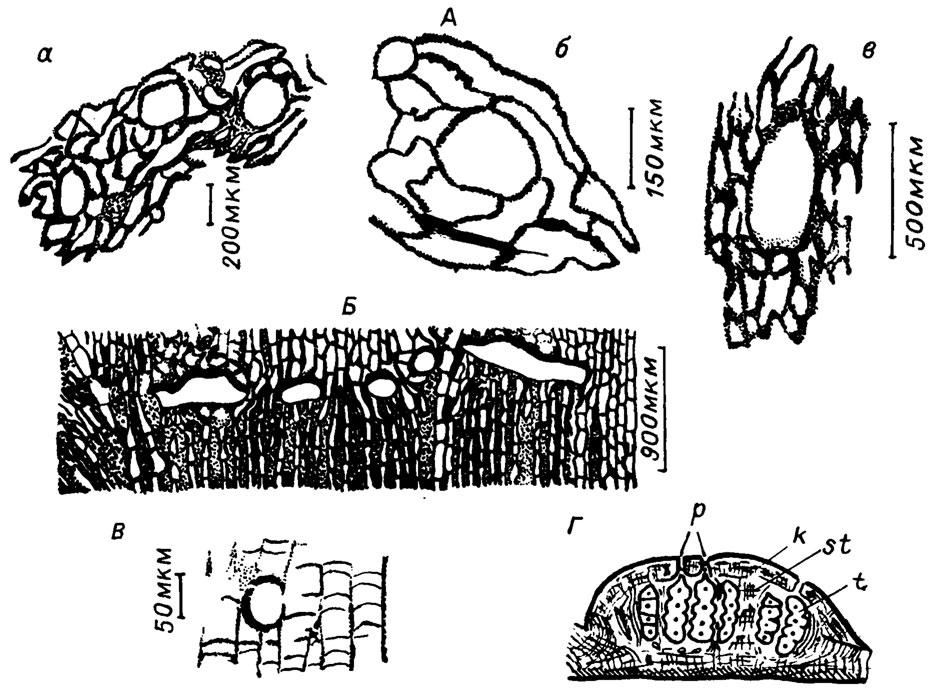
Рис. IV. 14. Органы размножения некоторых ископаемых красных водорослей. А - спорангии Solenopora spongoides (Dyb.) (по В. П. Маслову /1956а/): а - в поперечном сечении, б - при большом увеличении, в - продольное сечение через спорангий; Б - строение клеточных нитей, форма и расположение спорангиев у Solenopora (Neosolenopora) multiformis Bel. (по Л. С. Белокрысу /1966/); В - спорангии (?) среди нитей периталлия У Parachaetetes (Tomilithon) johnsoni Masl. (по В. П. Маслову /1962/); Г - сорус современного литотамния с тетраспорами (по В. П. Маслову /1956/): t - тетраспорангии, k - крыша, p - поры, st - стерильные нити
Довольно часто встречаются концептакулы в слоевищах архелитофиллюмов (рис. IV. 15). Они представляют собой крупные полусферические образования, располагающиеся среди клеток периталлия. Несколько отличаются от концептакулов архелитофиллюмов крупные яйцевидные концептакулы лысьвелл, которые располагаются в периферийной части слоевища под тонким однослойным покровом (см. рис. IV. 5) коровых клеток.

Рис. IV. 15. Две концептакуловые камеры Archaelithophyllum, окруженные мелкими клетками периталлия, внизу виден переход к крупным клеткам зоны гипоталлия, ув. 50 (по J. Johnson /1960/)
Другой тип органов размножения устанавливается у водорослей рода Katavella. Здесь на концах клеточных нитей располагаются очень крупные клетки, по крайней мере в два раза превосходящие по размерам обычные вегетативные (табл. XX, фиг. 4). По всей вероятности, эти образования также можно рассматривать в качестве концептакулов. Они близки по форме к таковым у Ludlovia.
Наличие или отсутствие органов размножения у палеозойских багрянок не учитываем как систематический признак, поскольку концептакулы встречаются очень редко и далеко не всегда однозначно идентифицируются.
Совсем иначе обстоит дело у кораллиновых водорослей. Здесь учитываются форма и взаимоотношение спорангиев, строение концептакулов (см. рис. IV. 11, IV. 16). Для этих водорослей строение органов размножения, их наличие или отсутствие - важнейший родовой признак. Это можно подтвердить всей системой кораллиновых водорослей. В одной из последних таксономических схем А. Поунана /Poignant, 1979 а, б/ придается важнейшее значение органам размножения для систематики мезо-кайнозойских багрянок. А. Поунан различает несколько типов строения репродуктивных органов (см. рис. IV. 11). Именно по этому признаку все мезо-кайнозойские красные водоросли разделены на две большие группы. Одна из них имеет изолированные спорангии и мультиперфорированные концептакулы, другая - моноперфорированные концептакулы (рис. IV. 17), По представлениям А. Поунана, систематика мезо-кайнозойских багрянок должна базироваться на внешней морфологии, текстуре клеточной ткани, строении репродуктивных органов. Из сказанного выше следует, что систематика палеозойских красных водорослей основана только на первых двух признаках. Значение морфологического признака также неустойчиво. Например, многие соленопорацеи обладают чрезвычайно изменчивой внешней формой слоевища. Соленопоры могут иметь таллом, меняющийся по форме от плоской пластины до шара с рядом промежуточных форм. Только внутренние детали строения клеточных нитей и их взаимоотношение позволяют диагностировать водоросли этого рода. Для соленопорацеи практически не имеет значения соотношение в слоевище гипоталлия и периталлия, как и полное отсутствие первого. Но эти признаки приобретают существенное значение для классификации красных водорослей с субцилиндрическим слоевищем. Для подобных форм соотношение срединного гипоталлия и периталлия - признак родового уровня или даже более высокого ранга.
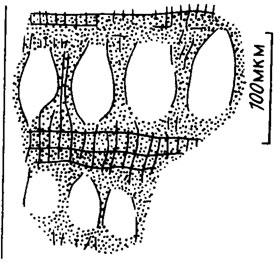
Рис. IV. 16. Расположенные рядами спорангии у Archaelithothamnium, видна деформация нитей клеток между спорангиями (по В. П. Маслову /1962/)
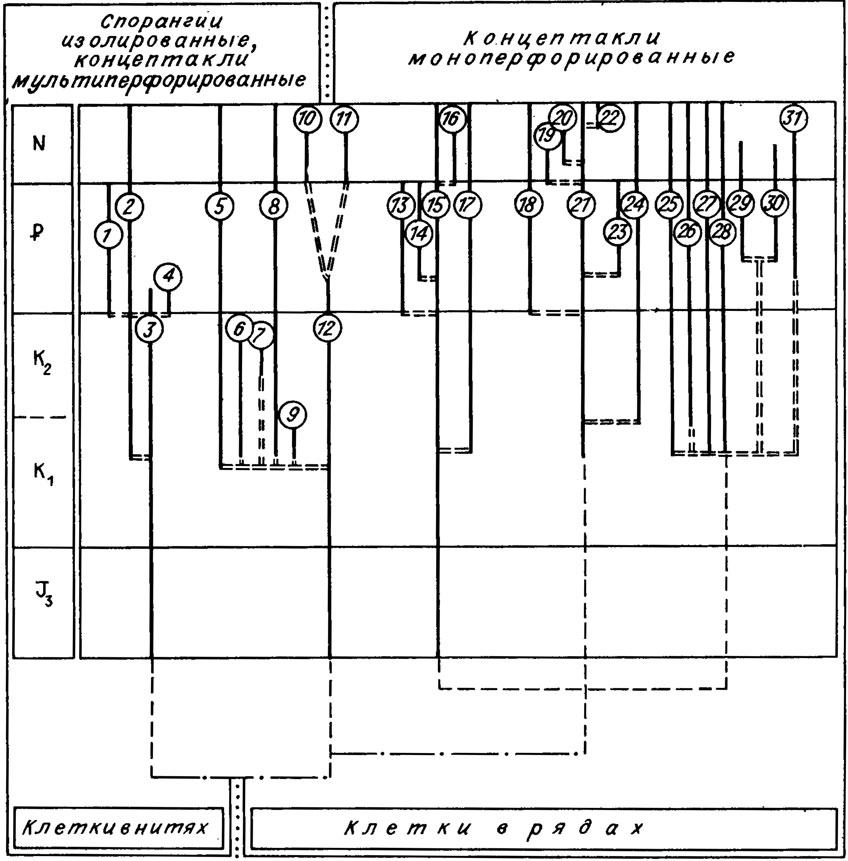
Рис. IV. 17. Филогенетические взаимоотношения и развитие мезозойских и кайнозойских кораллиновых красных водорослей, принципы их классификации (по A.F. Poignant /1979 а, b/). Цифрами в кружках обозначены: 1 - Paleophyllum; 2 - bithophyllum; 3 - Solenopora (Neosolenopora); 4 - Mesolithon; 5 - Archaelithothamnium; 6 - Paleothamnium; 7 - Hemiphyllum; 8 - Mesophyllum; 9 - Kymalithon; 10 - Pseudoaethesolithon; 11 - Aethesolithon; 12 - Parachaetetes; 13 - Distihoplax; 14 - Litholepis; 15 - bithoporella; 16 - Tenaria; 17 - Melobesia; 18 - Neogoniolithon; 19 - Paraporolithon; 20 - Porolithon; 21 - Litholepis; 22 - Paleolithophyllum; 23 - Leptolithophyllum; 24 - Dermalithon; 25 - Corallina; 26 - Arthrocardia; 27 - Jania; 28 - Amphiroa; 29 - Subterraniphyllum; 30 - Metagonolithon; 31 - Calliarthon
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'