
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Развитие красных водорослей
Наши представления об эволюции и филогенетических связях красных водорослей в значительной мере зависят от принятой системы. Первые, к сожалению не подтвержденные позднее, находки красных водорослей сделаны в верхнедокембрийских отложениях А. Г. Вологдиным. Из синийских отложений Приангарья описана Pustularia Vologdin, отнесенная к отделу красных водорослей на основании участия в стелющейся части слоевища клеточных нитей. В верхнесинийских отложениях Прибайкалья А. Г. Вологдиным найдены соленопоры. Представители рода Bija, описанные из среднекембрийских отложений Алтая, указываются сейчас из верхнедокембрийской формации Sarnyere Республики Мали /Bertrand-Sarfati, 1979/. Наличие Bija в кембрийских отложениях Сибири подтвердили К. Б. Кордэ /1973/ и Л. Г. Воронова /1976/. Все эти авторы относили Bija к красным водорослям, что кажется нам не правомерным. В принятой нами системе этот род отнесен к отделу синезеленых.
Богатейший мир красных водорослей описала К. Б. Кордэ /1961, 1973/ из кембрийских отложений СССР. Принадлежность к красным водорослям всего комплекса, изученного К. Б. Кордэ, неодинаково достоверна. Наиболее правильно, на наш взгляд, отнесение к багрянкам форм с клеточноподобным строением слоевища и хорошо выраженными терминальными органами размножения - роды Tubomorphophyton, Gordonophyton, Kosvophyton, Ludlovia, которые мы объединили в семейство Ludlovia и вывели из состава порядка Epiphytales.
Из нижнекембрийских отложений Сибири, Алтая и Монголии описаны /Рейтлингер, 1959; Кордэ, 1966, 1973; Дроздова, 1980/ два рода красных (? Б. Ч.) водорослей - Batenevia и Botominella. Слоевища этих водорослей сложены пучком гирванеллоподобных трубок - то очень плотно сжатых, то сравнительно рыхло уложенных. Наличие перегородок отличает клеточные нити названных родов от клеточных нитей гирванелл; у батеневий отмечены также овальные пустоты - спорангии. Отметим, что далеко не все палеоальгологи признают принадлежность этих двух родов к красным водорослям /Лучинина, 1975/.
Таким образом, в настоящее время наши представления о водорослях позднего докембрия и кембрия в значительной степени противоречивы. Если следовать К. Б. Кордэ, то кембрий можно без преувеличений назвать периодом пышного развития нитчатых багрянок. По другим представлениям, в кембрии имеются только проблематичные находки соленопорацей, а основной фон его альгофлоры составляют синезеленые водоросли.
С учетом отмеченных условностей можно сказать, что красные водоросли в кембрии были представлены несколькими морфологическими типами: формами с однорядным нитчатым слоевищем (Tubomorphophyton, Grordonophyton), массивными желвакоподными Solenopora и многорядными субцилиндрическими Batenevia.
Палеоальгологами неоднократно предпринимались попытки проследить во времени эволюцию красных водорослей и филогенетические связи между таксонами разного ранга /Johnson, 1960; Маслов, 1962; и др./. В большинстве филогенетических построений важная роль отводится соленопорацеям из родов Solenopora и Parachaetetes, которые образуют самостоятельную долгоживущую ветвь - от докембрия (?) до палеогена.
В. П. Маслов /1962/ рассматривал соленопоры как предков целой группы родов мелобезиевых - представителей триб Archaeolithothamnieae и Lithothamnieae. В этих построениях важное место занимает попытка проследить происхождение членистых багрянок. От рода Parachaetetes В. П. Маслов намечает боковую ветвь к нижнедевонскому роду Bicorium Maslov, 1956, который рассматривается, с одной стороны, как предковый род Lithoporella, а с другой - Archaelithophyllum. Архелитофиллюмы являются, по построениям В. П. Маслова, предковыми формами родов Lithophyllum, Porolithon, мелобезиевых и кораллиновых водорослей. Вся эта схема в высшей степени гипотетична, поскольку опирается в филогении членистых красных водорослей на проблематичный род Bicorium, не встреченный более ни автором, ни другими палеоальгологами*.
* (Мы считаем, что В. П. Маслов описал под этим родовым названием сечения водорослей типа. Rothpletzella или Plabellia.)
В филогенетических построениях Дж. Джонсона /Johnson, 1960/ в качестве предковых форм использованы роды Solenopora и Parachaetetes (рис. IV. 19). В середине каменноугольного периода от парахететеса отчленяется боковая ветвь рода Cuneiphycus, который рассматривается в качестве предковой формы всех членистых багрянок, начинающих свое существование в юре. В качестве гипотетического предка членистых багряно? принимается организм, принадлежность которого к водорослям вообще сомнительна (см. разд. "Система красных водорослей").
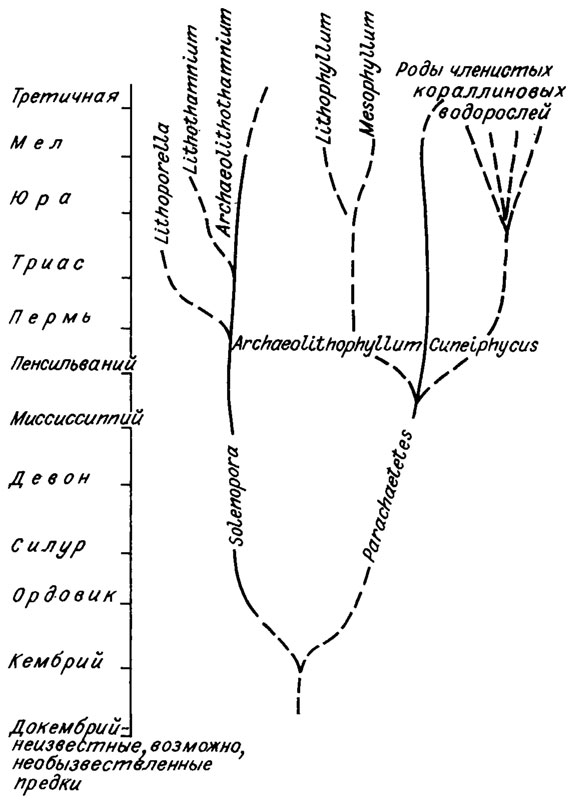
Рис. IV. 19. Схема происхождения кораллиновых водорослей (по J. Johnson, /1960/)
В качестве предковых форм других родов кораллиновых водорослей приняты Solenopora и Archaelithophyllum. В значительной мере базируется на соленопорациях филогенетическая схема (см. рис. IV. 17) А. Поунана /Poignant, 1979 а, b/. Накопленный материал по ископаемым красным водорослям позволяет по-иному реконструировать филогенетические связи ископаемых багрянок (рис. IV. 20). Мы намечаем несколько филогенетических рядов.
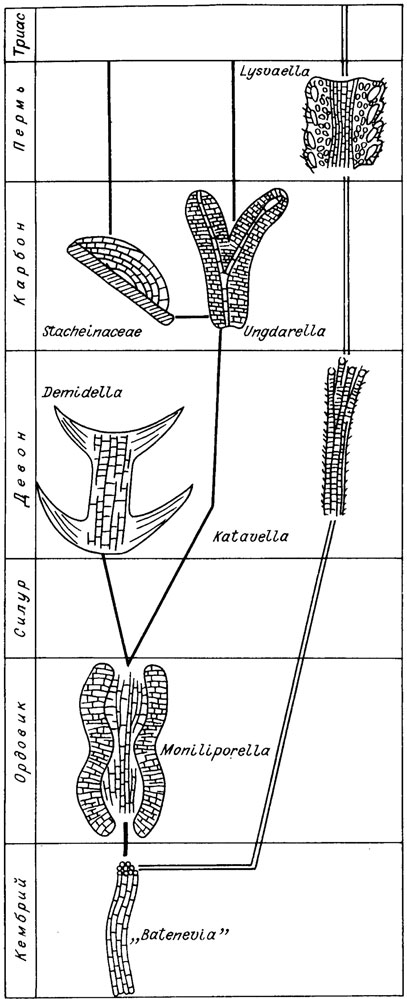
Рис. IV. 20. Филогенетические взаимоотношения между важнейшими родами палеозойских красных водорослей
В качестве предковых форм принимаются примитивные багрянки, обладающие слоевищем субцилиндрического типа, подобные Batenevia, из которых можно вывести принципиально новые по морфологии водоросли ордовикско-девонского семейства Moniliporellaceae, слоевище которых четко подразделяется на две зоны - центральную из вертикального пучка нитей (срединный гипоталлий) и зону периталлия, образованную клеточными нитями, расположенными почти перпендикулярно к осевой зоне.
Дальнейшее развитие этого ряда можно видеть в раннедевонских водорослях семейства Demidellaсеае (роды Demidella и Paralancicula). Это направление привело к возникновению очень высокоорганизованных багрянок.
Другое направление прослеживается от монилипорелляцей к унгдарелляцеям. Водоросли родов Ungdarella и Komia можно с достаточным основанием рассматривать как потомки Moniliporella, учитывая их морфологические сходство. Их сближает общность генерального плана строения слоевища: наличие вертикального пучка или одиночной нити срединного гипоталлия и отходящего под углом (иногда прямым) к оси слоевища пучка нитей периталлия, В качестве специализированной боковой (тупиковой?) ветви унгдарелляцей можно рассматривать (рис. IV. 21) довольно разнообразные в морфологическом отношении водоросли семейства Stacheinaсеае, как приспособившиеся к специализированному существованию в качестве тонких инкрустирующих корочек на разных предметах или на раковинных организмах. Можно предполагать их происхождение от унгдарелляцей способом неотении. Водоросли рода Archaelithophyllum мы рассматриваем как специализированную ветвь унгдарелляцей. Действительно, многие детали морфологии архелитофиллюмов противоречат их сопоставлению с членистыми кораллиновыми водорослями. Членистые багрянки имеют очень тонкие удлиненные клетки срединного гипоталлия и более крупные многоугольные - периталлия. У архелитофиллюмов, наоборот, срединный гипоталлий сложен крупными многоугольными клетками, а периталлий - мелкими.
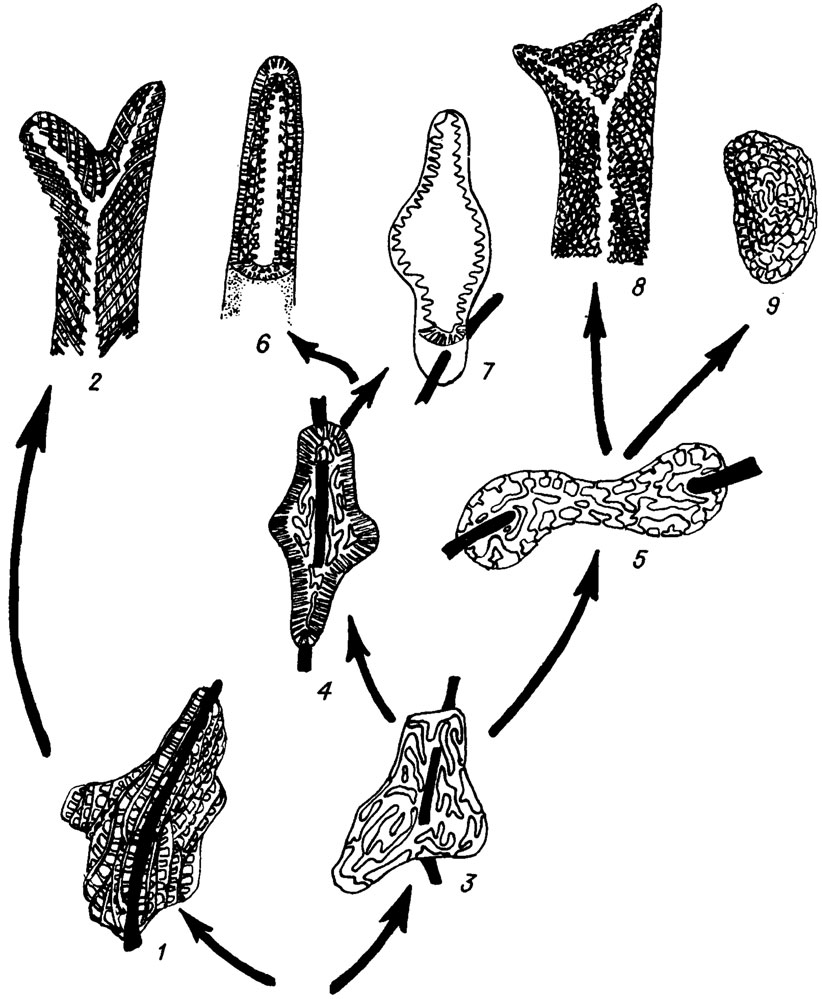
Рис. IV. 21. Филогенетические связи между родами семейства Stacheinaсеае (по М. Е. Perret, D. Vacharcl /1977/). Цифрами обозначены роды: 1 - Stacheoides; 2 - Ungdarella; 3 - Sinustacheoides; 4 - Epistacheoides; 5 - Dromastacheoides; 6 - Pseudokomia; 7 - Roguesselsia; 8 - Komia; 9 - Amorphia
Представляет интерес другой гипотетический ряд, который также можно начинать с водорослей типа Batenevia. Девонским потомком этого рода являются катавеллы, которые имеют субцилиндрическое слоевище, нечетко дифференцированное на гипоталлий и периталлий с одним рядом коровых клеток и внешними органами размножения. Дальнейшая линия развития может быть намечена и пермскому (конец ранней перми, кунгурский век) роду Lysvaella, представители которого имеют сложнодифференцированное слоевище субцилиндрической формы. Органы размножения расположены в периферийной части слоевища под однослойным коровым слоем клеток. Лысьвеллы долгое время оставались эндемичным уральским родом, но недавно /Vachard, 1980; Vachard, Moutenat, 1981/ они были описаны из артинских отложений Центрального Афганистана, что очень важно для обоснования выводов о происхождении членистых багрянок.
Из всех описанных палеозойских красных водорослей именно Lysvaella увереннее всего может быть помещена в качестве проблематичного предка кораллиновых водорослей. Лысвеллы (см. рис. IV. 5) имеют срединный гипоталлий, сложенный длинными узкими полигональными в поперечных сечениях клетками, а зона периталлия слагается крупными изометричными полигональными клетками. Именно такое строение имеют и слоевища членистых багрянок. Органы размножения у кораллиновых (Amphiroa), как и у Lysvaella, расположены рядами по боковым поверхностям. Оба сравниваемых объекта имеют высокие субцилиндрические слоевища. Сказанного достаточно, чтобы рассматривать Lysvaella и близкие к ним формы в качестве проблематичных и наиболее вероятных предков членистых кораллиновых водорослей. Мы далеки от мысли считать, что именно лысьвеллы - непосредственные предки членистых багрянок, но предположим, что эта линия развития привела к возникновению богатой альгофлоры членистых красных водорослей мезо-кайнозоя.
Для завершения общей картины филогенетических связей между красными водорослями необходимо рассмотреть развитие еще одной важной ветви - нитчатых однорядных водорослей семейства Ludloviaceae, т. е. эпифитоноподобных водорослей с признаками клеточного строения. Эта группа водорослей родов Gordonophyton, Tubomorphophyton и других появляется в кембрии и быстро занимает здесь важное место в составе альгофлоры. В силурийско-ордовикское время явно намечается резкое сокращение ареала их распространения и снижение значения в общем составе альгофлоры; только единичные находки родов Ludlovia и Kosvophyton подтверждают наличие водорослей этой группы. Мы пока не располагаем сведениями о существовании подобных форм в раннем и среднем девоне, но в позднем девоне недавно установлены в рифовых фациях представители (надо думать, последние) этой ветви - водоросли рода Tubomorphophyton /Чувашов и др., 1985/.
Рассматривая историю развития красных водорослей в общем плане, можно наметить несколько крупных этапов их эволюции. Первый, кембрийско-девонский этап - время существования нитчатых однорядных багрянок. В этот же период существуют высокоорганизованные красные водоросли семейств Moniliporella, Demidellaceae, Katavellaceae. Следующий, каменноугольно-пермский этап развития красных водорослей характеризуется стремительным увеличением морфологического разнообразия, что сопровождается вторжением красных водорослей во все биотопы мелководных шельфовых морей карбона и перми. Основу и наиболее яркую часть альгофлоры представляют унгдарелляцеи и стахеинацеи. Начало мезозоя - триасовый период - не выделяется ничем примечательным в истории красных водорослей. В триасе из всего комплекса пермских красных водорослей продолжили свое существование лишь два рода соленопорацей - Solenopora и Parachaetetes. Столь же бедны красными водорослями нижне- и среднеюрские отложения. На это время приходится заметное снижение роли багрянок в общем комплексе известковых водорослей. Это снижение компенсируется пышным расцветом зеленых водорослей, обильных и разнообразных в триасовых и юрских образованиях.
С поздней юры начинается последний, продолжающийся и поныне этап в развитии известковых красных водорослей. Первые членистые багрянки рода Archamphiroa появляются в начале позднеюрской эпохи. Вместе с ними начинают свое развитие мелобезиевые. В меловой и третичный периоды разнообразие альгофлоры красных водорослей непрерывно расширяется, они занимают все более разнообразные экологические ниши, осваивают большой диапазон глубин: морского бассейна, активно участвуют в создании крупных рифовых массивов. Настоящее время без преувеличения можно считать эпохой расцвета красных водорослей. В графической форме этапы развития известковых красных водорослей приведены на схеме (рис. IV. 22).
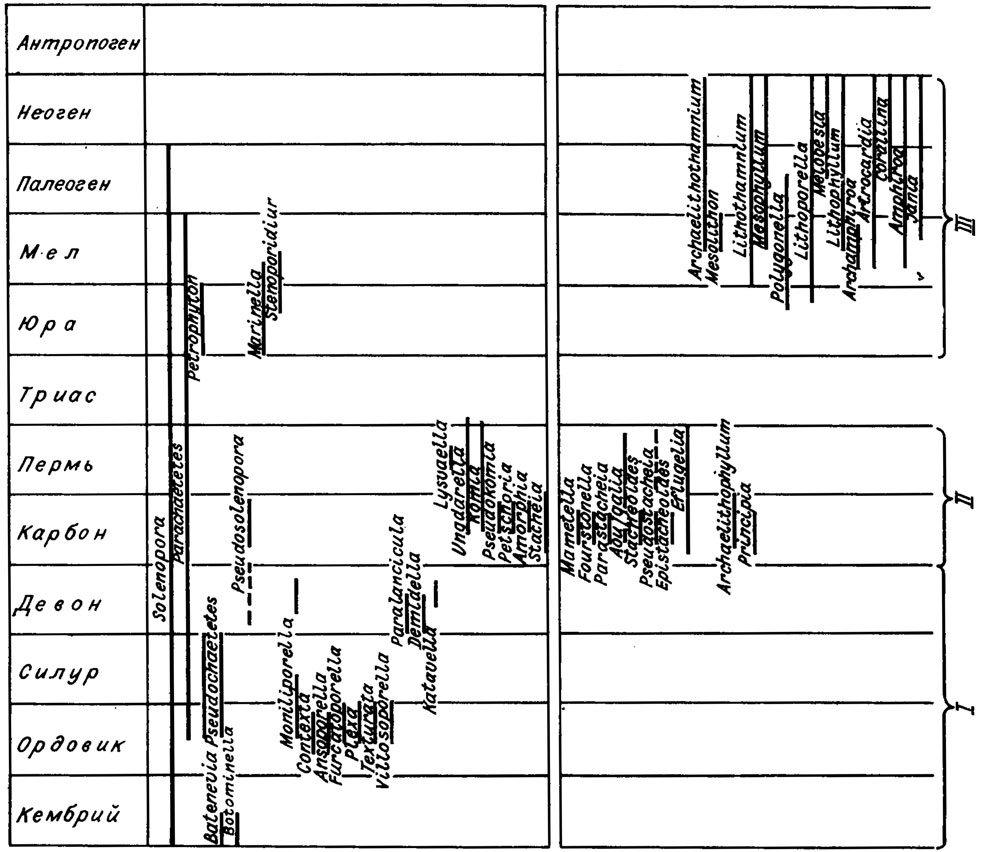
Рис. IV. 22. Стратиграфическое распространение палеозойских и важнейших родов мезо-кайнозойских (см. также рис. 17) красных водорослей
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'