
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Подсемейство Nitelloideae Braun, 1897
Гирогониты со сложной базальной пластинкой (2- и 3-членные), у ныне живущих форм - с десятью коронарными клетками.
Гирогониты толипеллоидного типа с заостренной вершиной, субшаровидные или сжатые сбоку; поверхность часто имеет зернистую орнаментацию (см. рис. V. 3).
В составе подсемейства шесть родов: Sphaerochara, Maedlerisphaera, Kaskyechara, Collichara, Tolypella и Toiypella ныне живущая с необызвествляющимися оогониями.
Распространение. Верхняя юра - ныне.
Основные черты палеоэкологии. Харофиты считаются типично пресноводными растениями. Большинство современных харовых водорослей растет на малой глубине в пресных водах (прудах, каналах, озерах, лиманах и по берегам рек со спокойным течением). Однако как в другие геологические эпохи, так и в настоящее время некоторые харофиты приспособились к условиям солоновато-водных небольших водоемов, озер и морских прибрежных участков и лагун. В основном они произрастают в умеренной и аридной климатических зонах, но встречаются также в жарких и холодных поясах.
Ископаемые харофиты характеризуются достаточно разнообразными условиями произрастания и захоронения. В силурийских отложениях, в наиболее древних находках харофитов, они встречены совместно с остатками наземных растений, а также мохообразных и плауновидных, существовавших в условиях солоновато- и пресноводных водоемов, т. е. в континентально-лагунной обстановке. Здесь обызвествление происходило, очевидно, замедленно, чем, по-видимому, объясняется своеобразное состояние сохранности позднесилурийских харофитов Подолии, захороненных частично в виде отпечатков, частично в форме слабоминерализованных ядер вегетативных осей и ооспорангий. Наличие массовых скоплений однотипно ориентированных вегетативных осей харофитов, в изобилии встречающихся на плоскостях наслоения породы, позволяет сделать вывод, что они образовывали густые заросли на дне заселенного ими водоема /Ищенко, Сайдаковский, 1975/.
В девонских отложениях Тянь-Шаня харофиты приурочены к прослоям пород лагунного и прибрежно-морского происхождения /Поярков, 1966/.
В верхнем карбоне Донбасса харофиты обнаружены в глинах и глинистых известняках, образовавшихся в опресненных лагунах регрессирующего каменноугольного моря /Шайкин, 1966/.
Харофиты перми и триаса приурочены к огромным толщам пестроцветных глин и алевритов, отлагавшихся в континентальных водоемах в условиях аридного климата.
Остатки меловых и палеогеновых харофитов характерны для отложений внутренних континентальных водоемов; это фации обширных озер, лагун и реже прибрежно-морских опресненных зон. Харофиты встречаются в глинистых красно- и пестроцветных породах, а также в породах, обогащенных карбонатом кальция и образованных при аридном и реже семиаридном климате, в прибрежных освещенных зонах глубиной от 1-2 до 3,5 ми при большой прозрачности воды (до 30 м) /Кянсеп-Ромашкина, 1981/.
В верхнем меду Монголии харофиты встречаются только в мелкозернистых песчаниках и легких серых песках, в которых содержание карбоната кальция достигает 42%. Высокое содержание карбоната объясняется, вероятно, высыханий ем осадочного бассейна, что подтверждается сильной кальцификацией корковых клеток и наличием дополнительных ребер на партекальцинах. В комплексе харофитов встречается много очень мелких (от 200 до 300 мкм) гирогонитов с сильно утолщенными стенками и характерной микроструктурой поверхности, что свидетельствует о неблагоприятных условиях в период их развития /Karczewska, Ziembinska-Tworzydlo, 1983/.
В эоцене харофиты обнаружены на Украинском щите в прослоях серых глин между горючими сланцами в локальной Болтышской впадине; геологическое строение свидетельствует, что здесь в палеогене находилось озеро.
В неогене Казахстана обильные находки гирогонитов заключены в светло-серых и светло-коричневых глинах, суглинках и супесях, перекрытых красноцветами континентального происхождения.
В Северном Причерноморье харофиты встречаются в отложениях лиманных бассейнов с пресной и полупресной водой. В аналогичных условиях ныне живут харовые водоросли в лиманах Причерноморья, озерах Казахстана, устьевых частях р. Амударьи и т. д. /Маслов, 1966/.
Харофиты играют некоторую роль в образовании горных пород. Известняки и мергели образуются в результате обильного роста харофитов в озерах третичного и современного периодов. Обычно карбонат, выделяемый вегетативными частями (главным породообразующим элементом харофитов), представляет собой довольно чистую известь. Как показывает рентгенокристаллографический анализ "спелых" известковых оболочек современных хар /Horn af Rantzien, 1959 а, b/, гирогонитов и утрикулов мезозойских харофитов (рис. V. 6), они состоят из кальцита. Отложения, образуемые современными харами, эксплуатируются в озерах с достаточно мощным осадком. Встречающиеся иногда породы, образованные в основном известковыми остатками харофитов, называются харацитами. В СССР известен харацит из плиоцена (Южная Тува), образующий метровый пласт писчего мела, используемого для местных нужд. Харациты встречаются и в других районах СССР, в Западной Европе и Америке, но их роль в образовании осадочных толщ незначительна /Маслов, 1963 а, б/.
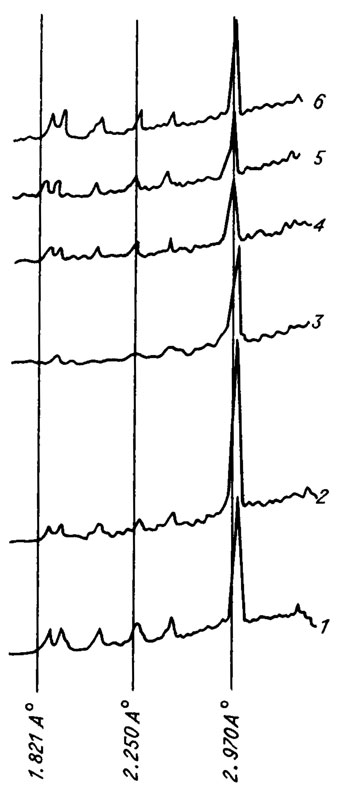
Рис. V. 6. Результаты рентгеноструктурного анализа гирогонитов и утрикулов мезозойских харофитов (составил И. М. Шайкин). 1 - эталон кальцита; 2 - гирогонит Porochara (J3km-ti); 3 - утрикул Flabellochara(J3v); 4 - утрикул Nodosoclavator (J3v); 5 - гирогонит Nodosoclavator (J3v); 6 - гирогонит Maslovichara (T1)
Филогения и стратиграфическое значение харофитов. Харовые водоросли рассматриваются как отдельный тип, который давно дифференцировался от зеленых водорослей (сифоней?), и как первая попытка развития новых свойств, близких к высшим растениям /Grambast, 1973/.
К. Мэдлер /Madler, 1952, 1953, 1955, 1957/ отметил у древних харофитов наличие простой открытой апикальной поры, а у более молодых - ее отсутствие и предположил, что древние харовые водоросли оплодотворялись с помощью спор, как и другие водоросли, а оплодотворение сперматозоидами продукт эволюции, приведшей к уменьшению количества партекальцин и изменениям морфоструктуры вершины.
Современное состояние изучения харофитов, показывает, что они являются одной из групп растений, хорошо вписываемых по филогенетическим признакам в ряд окаменелых популяций, поддающихся определенным принципам систематики. Вопрос о происхождении и филогенетических связях между тремя порядками харофитов является дискуссионным. Филогения внутри каждого порядка установлена более или менее определенно, хотя требует дальнейших доработок.
Древнейшие представители Sycidiales и Trochiliscaies известны из силура, но они уже имеют довольно высокую степень организации женского органа размножения. Что же появилось раньше: формы с завивающимися партекальцинами или с вертикально расположенными элементами известковой оболочки? К. Мэдлер /Madler, 1952, 1953, 1957/, рассматривая этот вопрос, напоминает, что у современных харофитов эмбриональные клетки спорофидия вначале не имеют завивания (см. рис. V. 1). Это следует трактовать как явление, соответствующее одному из основных биологических законов, когда онтогенез проливает свет на филогенез. И вслед за Р. Пеком /1934 b/ К. Мэдлер Matdler, 1952, 1953, 1957/ считает сицидиумы наиболее примитивными харофитами не только на том основании, что клетки оболочки у них не завиваются, но и потому, что эти клетки имеются в наибольшем количестве. У них насчитывается не менее 20 клеток оболочки, которые образовались, полагает Р. Пек, благодаря тому, что в эмбриональном состоянии имелось 10 клеток, из которых позднее каждая разделилась по длине надвое. У более поздних харофитов произошла редукция количества клеток оболочки: у трохилисков до 7-12, у палеохар - 6, харацей - 5. У них возможно также повторное деление, причем для лучшего укрепления возникли витки.
На основе новых данных Л. Грамба /Grambast, 1962, 1966, 1973/ дает несколько иное толкование филогении древнейших харофитов. В отличие от современных харофитов древние, особенно в девонском периоде, обнаруживают большое разнообразие в строении остатков известковых оболочек женского органа размножения. В это время существуют четыре семейства (по данным 1985 г., не менее семи), представляющие все три порядка харофитов. У трохилисков партекальцины продолговатые и завернуты вправо, у сицидиумов известковая оболочка образована вертикальными элементами, каждый из которых состоит из мелких ячеек. У хованелл наружные элементы оболочки также вертикальны, но, как у трохилисков, образуют одну клетку. Следовательно, строение известковой оболочки хованелл можно считать наиболее примитивным среди харофитов, хотя в додевонских отложениях они не обнаружены. Вероятно, хованеллы - прямые потомки основного ствола, от которого, с одной стороны, произошли сицидиумы, а с другой - трохилиски. Возможно также, что эти особые формы имели независимое происхождение от общего примитивного типа, у которого не образовывался гирогонит, поскольку орган размножения не имел наружных стерильных клеток. Женский спорофидий, вероятно, впервые появился путем сращения вегетативных элементов вокруг простой женской яйцеклетки.
Хованеллы, сицидиумы и умбеллы оказались неприспособленными к дальнейшей эволюции и вымерли в конце девонского и начале каменноугольного периодов. И лишь новые формы харофитов - эохары, появившиеся в средне девонскую эпоху и имеющие сходство с трохилисками по большому числу партекальцин, но с левым их завиванием, дали новую филогенетическую линию харофитов, дошедшую до наших дней. И, как отмечает Л. Грамба /Grambast, 1973/, несмотря на незначительную роль среди харофитов в девоне, эохары оказались растениями с более значительным эволюционным потенциалом, чем пышно развившиеся в то время харофиты двух других порядков.
Эволюция харофитов, начало которой положили эохары, характеризуется прогрессивным уменьшением и фиксацией количества партекальцин с 8-13 до 6 у палеохар и стабилизацией на 5 у порохарацей и всех последующих поколений, вплоть до современных.
Porocharaceae, сохранившие от своих предков/апикальную пору, стали связующим звеном между палеозойскими и мезо-кайнозойскими харофитами. В пермский и триасовый периоды они были, по-видимому, единственным семейством, представлявшим харофиты в континентальных водных бассейнах. Отдельные их представители дожили до эоцена, но еще в юрский период это обширное семейство дало ряд новых филогенетических линий (рис. V. 7.). Одни (стеллятохары и масловихары), очевидно, были предками Clavatoraceae, гирогониты которых имеют апикальную пору и вытянутую в шейку вершину, но покрыты утрикулом; другие, вероятно из подсемейства Cuneatocharoideae, дали начало и ныне живущему семейству Characeae, у которого исчезла апикальная пора; наконец, третьи (род Musacchiella с сегментированной базальной пластинкой) могли развиться в сторону семейства Raskyallaceae путем создания "крышечки" над апикальной порой /Feist et al., 1984/.
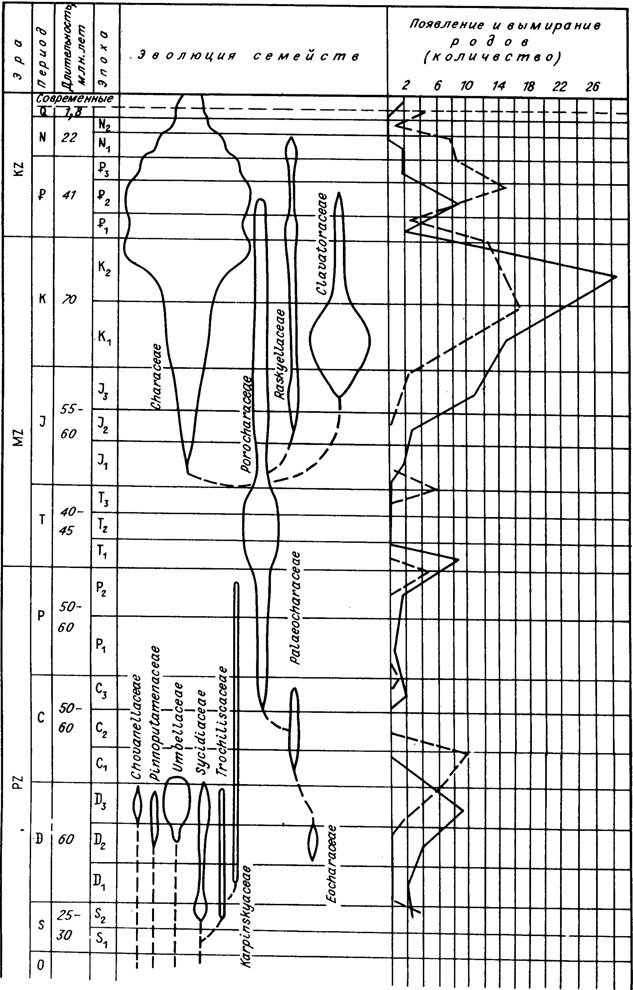
Рис. V. 7. Эволюция харофитов в фанерозое (составил И. М. Шайкин). Сплошной линией обозначено появление родов, штриховой - вымирание
Верхнеюрская эпоха и меловой период стали временем пышного развития своеобразного семейства Clavatoraceae, имевшего большой эволюционизирующий комплекс родов и видов, несмотря на сравнительно недолгое существование (см. рис. V. 7).
В этом семействе четкие морфологические изменения и филогенетические линии характерны для утрикулов при устойчивости морфологии гирогонитов. Основное направление филогенеза утрикулов - это модификация и дифференциация составляющих элементов, что приводит к большой сложности их строения. Л. Грамба /Grambast, 1962, 1966, 1973/ прослеживает как общую филогению этого семейства, так и отдельные ряды и линии, дающие последовательное развитие новых родов и видов (см, рис. V. 5). Развитие утрикулов во времени происходит от простых, не уплотненных обволакивающими гирогониты вегетативными элементами Echinochara (верхняя юра) до сложных спаянных "настоящих" утрикулов со сложной симметрией за счет дополнительных выступов, пор и поверхностной орнаментации, закономерно фиксирующих филогенетические связи у родов и видов Perimneste, Atopochara, Flabellochara-Clypeator, Glovator и др.
Развитие семейства Characeae заключалось главным образом в изменении морфоструктуры вершины и общей формы гирогонитов, появлении и исчезновении орнаментации на их поверхности. Занимая незначительное место в верхней юре и нижнем мелу, это семейство быстро дифференцируется на многочисленные роды и виды в конце мелового периода, достигая расцвета в эоценовую и олигоценовую эпохи. В кайнозое Characeae - единственное семейство харофитов, но представлено оно значительным количеством таксонов. Начиная от миоцена заметно сокращается видовой и родовой состав, который в современных водоемах весьма скромен в сравнении с предыдущими эпохами.
Таким образом, можно констатировать, что во все геологические эпохи харофиты как растительное сообщество с момента их появления находились в постоянном развитии, при котором новые формы сменяли старые. Благодаря этому они получили определенное стратиграфическое значение, особенно в связи с полным или частичным отсутствием другой ископаемой флоры и фауны в толщах континентального и прибрежно-морского происхождения. По харофитам хорошо фиксируется и коррелируется ряд горизонтов в девонских, юрских, меловых, палеогеновых и неогеновых отложениях. А для расчленения триасовых, нижнемеловых, эоценовых и олигоценовых отложений многих регионов по харофитам разработаны и используются весьма детальные биостратиграфические схемы.
Primochara ellipsoidea A. Istchenko, sp. nov.
Табл. XXIII, фиг. 1 а, б
Видовое название от Ellipsoideus - эллипсоидальный (лат.)
Голотип - экз. № 120/846. ИГН АН УССР. Подольское Приднестровье, р. Баговица; лудловский ярус, баговицкая свита, мукшинская подсвита.
Диагноз. Гирогонит эллиптический, зауженный кверху. Партекальцины многочисленные, тонкие.
Описание. Форма гирогонита в продольном срезе эллипсоидальная, в поперечном - округлая. Высота гирогонита - 1,07 мм, ширина в экваториальной части - 0,57 мм. К вершине гирогонит постепенно суживается, образуя невысокую горловину шириной 0,17 мм, которая заканчивается вверху заостренными зубцами. В продольном срезе видны четыре зубца: два расположены вертикально, два наклонены в стороны. Высота зубцов 0,04-0,05 мм, ширина у основания 0,03-0,04 мм,
Базальная часть гирогонита слегка расширенная, снизу уплощенная. Базальное отверстие диаметром 0,03 мм ограничено выступающими наружу острыми зубцами длиной 0,02 мм. Базальная пластина не наблюдается. По обе стороны от базального отверстия на некотором расстоянии от него расположен шипообразный вырост, суживающийся к концу, длиной 0,114 мм, шириной у основания 0,034 мм.
Внутренние полость и стенки партекальцин заполнены перекристаллизованным кальцитом. Внешние стенки разрушены. Их поперечные сечения (12) четко видны только в верхней половине продольного среза гирогонита. Диаметр партекальцин не превышает 0,018-0,020 мм.
Сравнение. Отличается от типового вида Primochara calvata T. Istchenko et Saidakovsky размерами гирогонитов, их формой и количеством партекальцин.
Замечание. Описанный вид отмечался нами ранее как Primochara sp. /Ищенко, 1985/ с кратким диагнозом. Дополнительный материал позволил описать его детальнее. Однако в связи с перекристаллизацией карбоната, из которого состоял гирогонит, невозможно установить количество витков партекальцин. Судя по тому, что в верхней части гирогонита с одной его стороны отмечаются 12 поперечных срезов партекальцин, возможно, и в нижней части их было не меньше. Тогда общее количество срезов партекальцин должно составить около 20-24. Возможно также, что в нижней части гирогонита обызвествление партекальцин было слабее, чем в верхней, и поэтому в ископаемом состоянии их стенки не сохранились.
Местонахождение. Подольское Приднестровье, левый приток р. Днестр, р. Баговица; лудловский ярус, боговицкая свита, мукшинская под свита.
Материал. Пять сечений в шлифах.
Primochara smotrichi A. Istchenko, sp. nov.
Табл. XXIII, фиг. 2, 3, а, б.
Видовое название от местонахождения на р. Смотрич.
Голотип - экз. № 121/120. ИГН АН УССР. Подольское Приднестровье, левый берег р. Смотрич у с. Заводянка; лудловский ярус, коновская свита, голосковская подсвита.
Диагноз. Гирогонит яйцевидный, уплощенный в базальной части, в продольных срезах сбоку 12-14 поперечных сечений партекальцин.
Описание. Гирогонит высотой до 0,63 мм, шириной до 0,36 мм. В продольных сечениях четко видна внутренняя полость высотой 0,40-0,45 мм, шириной 0,26-0,29 мм, заполненная перекристаллизованным кальцитом с редкими сгустками микрита. В апикальной части у типового экземпляра расположены шипообразные вытянутые вверх выросты с закругленными концами. Выросты плотно примыкают друг к другу; высота их 0,13-0,15 мм, ширина каждого 0,05-0,06 мм. Имеется несколько продольных сечений без выростов или с короткими лопастевидными. Базальная часть уплощена снизу. Базальное отверстие шириной 0,034 мм ограничено с двух сторон выступающими окончаниями партекальцин.
В продольных срезах с обеих сторон внутренней части гирогонита расположены поперечные сечения партекальцин. Их стенки перекристаллизованы в различной степени, сложены желтовато-серым мелкокристаллическим кальцитом, С каждой стороны продольных срезов гирогонита отмечаются 12-14 поперечных срезов партекальцин. Толщина стенки партекальцин 0,010 мм, диаметр внутреннего отверстия 0,026 мм.
Сравнение. От типового и нового видов Primochara. ellipsoidea sp. nov. отличается размерами гирогонитов, количеством и диаметром партекальцин.
Замечание. Продольное сечение гирогонита было кратко описано нами /Ищенко, 1985/ как Primochara, sp. 2. Дополнительный материал, полученный из скважин, позволил описать новый вид.
Местонахождение. Хмельницкая обл., скв. 3664, гл. 294 м; венлокский ярус, суршинская свита, гл. 231 м - лудловский ярус, коновская свита шутновская подсвита. Река Смотрич у с. Заводянка; лудловский ярус, коновская свита, голосковская подсвита, Днестр у с. Лука Врублевецкая; баговицкая свита, мукшинская подсвита.
Материал. Около десяти сечений в шлифах.
Sycidium sp.
Табл. ХХIII фиг. 8
Описание. В поперечном срезе известковая оболочка эллиптическая. Внутренняя полость заполнена округлыми зернами кальцита, частично микритизированными, ближе к стенке - перекристаллизованными. Высота внутренней полости 0,30 мм, диаметр в центральной части 0,22-0,23 мм. Стенка желтовато-серая, сложена криптокристаллическим кальцитом. Толщина стенки 0,024-0,030 мм. Наружная поверхность неровная, с остроугольными выступами длиной 0,036 мм. Стенка состоит из 12-14 субквадратных полей шириной 0,018-0,026 мм, разделенных швами толщиной 0,004-0,005 мм.
В апикальной части имеется четыре лопастевидных выроста высотой 0,040 мм, шириной у основания 0,020, вверху 0,050 мм, расположенных в виде розетки. В базальной части отмечаются два лопастевидных выроста, направленных в противоположные стороны друг от друга. Базальное отверстие из-за перекристаллизации зерен кальцита не просматривается.
Замечание. В силурийских отложениях Подольского Приднестровья выявлены остатки харофитов, отнесенные к новому роду Pratesycidium T. Istchenko et A. Istchenko /Т. А. и А. А. Ищенко, 1982/, который отличается наличием вегетативных осей и крупными (до 3-4 мм в диаметре) известковыми оболочками (гирогонитами?). Описанный нами поперечный срез известковой оболочки имеет микроскопические размеры (почти в 10 раз меньше) и не может быть отнесен к этому роду. Поскольку это единственный экземпляр такой известковой оболочки из силурийских отложений, мы не можем пока описать его детальнее. Но принадлежность его к роду Sycidium несомненна.
Местонахождение. С. Дарахов Тернопольской обл., скв. 1, гл. 552 м; пржидольский ярус, звенигородская свита.
Материал. Один поперечный срез в шлифе.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'