
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Глава третья. Сибирь и Африка (гиппарионовая фауна)
"...В северных краях в древние веки великие жары бывали, где слонам родиться и размножаться, и другим животным, также и растениям около екватора обыкновенным держаться можно было; а потому и остатки их здесь находящиеся не могут показаться течению натуры противны..."
М. В. Ломоносов. "О слоях земных 1757-1759.
Первая половина лета 1928 г. в Западной Сибири и Северном Казахстане была дождливая. Мы с Е. Д. Шлыгиным, одним из лучших знатоков геологии Казахстана, тогда еще совсем молодым геологом, подолгу отсиживались в палатке на берегу Бе-текея. Глинистая почва сильно набухала от воды и так размокала, что колья, на которые была натянута палатка, во время ветра упорно вылезали из земли. Палеонтологические сборы были скудны. Но мне предстояла поездка па Иртыш для осмотра его берегов и поисков третичных млекопитающих, и в июле л уехал в Омск. Здесь меаш приветливо встретил геолог проф. П. Л. Драверт (1879-1945), поистине влюбленный в Сибирь, начиная еще со времен своей дореволюционной ссылки. К этому времени П. Л. Дравертом на Иртыше были сделаны палеонтологические сборы и найдена интересная неогенов а я флора. Она, очевидно, когда-то "кормила" разнообразных неогеновых копытных, на которых охотились их неизменные спутники - хищники. Но в Омском музее не нашлось никаких следов неогеновых млекопитающих. Где они все и где найти их кости?
Еще в Ленинграде после совещания с П. И. Преображенским и А. А. Борисяком было намечено осмотреть берега Иртыша, представляющие до известной степени естественный меридиональный разрез через "третичные" и "четвертичные" отложения Западной Сибири. Для начала я обследовал несколько крупных обнажений по высокому правому берегу Иртыша, от Омска вверх по реке до Павлодара, в который прибыл на пароходе.
Еще издалека с Иртыша, при подходе снизу к Павлодару, бросалось в глаза длинное обнажение желтых, охристых песков и розовых пятнистых глин правого берега. Предположительно им приписывался неогеновый возраст, но они не были охарактеризованы палеонтологически.
Павлодар, в наше время крупный областной центр, тогда был скромным степным уездным городом. Железная дорога доходила до Иртыша, но железнодорожного моста еще не было.
Устроившись в гостинице, я вышел к пароходной пристани и пошел по бечевнику Иртыша вниз по течению, тщательно осматривая разрез песков и глин правого берега. Дойдя до места, носившего тогда название "Гусиный перелет" или "Разгуляй", я увидел на бечевнике, частью в воде, белые куски костей млекопитающих, зубы, среди них вскоре нашел характерную лопатовидную, слегка раздвоенную коронку зуба крупного жвачного. Сомнений быть не могло - это зуб жирафы! (рис. 4). Осмотревшись, я увидел, что берег на протяжении метров двадцати сплошь усыпан обломками костей, челюстей и разрозненными зубами млекопитающих. Кое-где вперемежку с ископаемыми лежали и кости домашних животных - бараньи, коровьи и лошадиные; но ископаемые легко отличались от современных ярко-белым цветом со ржавыми пятнами от окислов железа, тяжелым весом и хрупкостью, высохшие давали сильную "отлипь" на языке. Осмотрев обнажение, я увидел вверху слои песков и глин, из которых торчали и вываливались кости. Жирафа, носорог, небольшая лошадь с очень извилистым рисунком тонкой эмали на коренных зубах, какие-то гиенообразные хищники - легко устанавливались при самом беглом взгляде на это скопление. Сомнений быть не могло: передо мною было кладбище "пикермийской фауны" (по ее первому раскопанному в прошлом столетии местонахождению близ деревни Пикерми около Афин в Греции), иначе "фауны гиппариона"; она чаще называется так по ее характерному члену - гиппариону, небольшой трехпалой ископаемой лошади, обычно преобладающей в этой фауне над другими копытными по количеству костных остатков. Я был так поражен находкой, притом в самом городе на берегу Иртыша, где геологи бывают ежегодно, что даже растерялся. Придя в себя, стал спешно собирать найденное; набрав рюкзак, снял часть одежды, превратил ее наскоро в мешки, которые, наполнив костями, едва смог поднять. Какие-то мальчуганы, катавшиеся на лодке, довезли меня до пристани, откуда я доставил коллекцию на извозчике в гостиницу. Переодевшись, я направился в столовую, где меня нашел приехавший в Павлодар П. И. Преображенский. В тот же день я показал ему и свои первые сборы и местонахождение. Решено было поставить небольшую пробную раскопку; она сразу же показала обилие костных остатков. Отправка этой коллекции по железной дороге не обошлась без курьеза: с меня настойчиво требовали справку ветеринара о том, что жирафы, носороги и прочие погибли не от эпизоотии. Впоследствии сборы в Павлодаре производились неоднократно, по наиболее крупные коллекции были добыты раскопками Палеонтологического института Академии наук в 1929 и 1930 гг. (см.: Природа, 1930, № 1; рис. 5, 6, 7) и в последние годы Академией наук Казахстана. Материалы из Павлодара изучены еще не во всех подробностях. Сделаны новые находки гиппарионовой фауны в Казахстане и Сибири, недавно обнаружено большое местонахождение в Киргизии. Имеющиеся данные о фауне Павлодара представляют нешмнениый интерес, так как сведений о третичных млекопитающих Сибири очень мало.
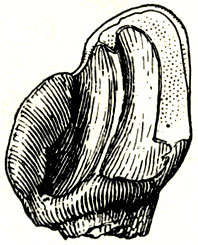
Рис. 4. Левый нижний клык жирафы самотерия (Samotherium sp.) Павлодар. Палеонтологический институт АН СССР. Увеличение 1,5
Берег Иртыша на месте раскопок имеет следующее строение (рис. 5). На выходящий из-под уровня воды Иртыша белый мергель (палеонтологически немой, по Н. К. Высоцкому (1896) - предположительно морской верхний олигоцеи) налегают снизу вверх:

Рис 5. Правый берег Иртыша у Павлодара на месте раскопок 1929-1930 гг. а - общий вид роскопок с левого берега (фото Д. П. Багаева).
1. Пески средне- и мелкозернистые, диагонально-слоистые с большим количеством блесток слюды (мусковит), местами образующей сплошные прослои; до 8-10 м.
2. Суглинистый песок с блестками мусковита; до 1,5 м.
3. Глина мергелистая, очень плотная, светло-серая; до 1 1/2 м.
4. Глина мергелистая, розовая с голубыми пятнами; ее верхний край со следами размыва; 2 м.
Четвертичные отложения:
5. Пески желтовато-бурые, горизонтально-слоистые, немые; от 1 до нескольких метров.
6. Карбонатный горизонт вышележащей погребенной почвы и погребенная почва; до 1,5 м.
7. Современные образования (дюны, тонкий слой почвы); до 1 м.
Кости залегают в слоях 1, 2 и 3; таким образом, общая вертикальная мощность костеносной толщи достигает 10-12 м, но главная масса костей захоронена в верхней части диагонально-слоистых песков (слой № 1, где залегает лучший по сохранности материал и где встречаются кости во взаимном сочленении), в глинистом песке (слой № 2), и в нижней части мергелистой глины (слой № 3). В этом последнем слое, помимо целых костей и черепов, имеется огромное количество мелких обломков, местами сплошь заполняющих глшту, частично сильно окатанных, а также разрозненных зубов. Диагонально-слоистые слюдистые пески тянутся по берегу Иртыша на несколько километров и обязаны своим отложением скорее всего крупной водной артерии, проходившей здесь в неогене после освобождения от морской воды и превращения в сушу "иртышского залива" палеогенового моря. Сильное течение несло и нагромождало в беспорядке черепа и тяжелые кости (носорогов и других копытных); отложение же налегающих па диагонально-слоистые пески глинистых слоев, видимо, шло уже в водоеме с илистым дном, быть может типа заводи или старицы. Следовательно, богатое скопление костей павлодарского местонахождения, как и многих других этой фауны, имеет водное происхождение, и образование этого скопления, быть может, шло не один сезон.

Рис 5. Правый берег Иртыша у Павлодара на месте раскопок 1929-1930 гг. б - берег на месте раскопок (с картины В. П. Батурина в Музее Палеонтологического института АН СССР)
Количество захороненного здесь костного материала огромно: подсчет одноименных костей одного и того же вида животных показал, что при вскрытии раскопками Палеонтологического института небольшой части местонахождения, размываемого Иртышом многие годы, были обнаружены остатки не менее 20 особей оленей, 40 жираф, 130 носорогов, более 200 гиппарионов и т. д. Между тем раскопками захвачена лишь незначительная часть этого кладбища, разрушаемого половодьями Иртыша, как выяснилось, многие десятки лет...

Рис. 6. Раскопки гиппарионовой фауны у Павлодара (1929 г.)
Различные животные этой фауны представлены в сборах, очень неравномерно. Условия образования местонахождения (перенос сильным течением тяжелых черепов и костей носорогов, жираф и других копытных) не были благоприятны для сохранения в целости хрупких костей газелей, мелких хищников, грызунов, насекомоядных, ящериц п т. п. Остатки мелких животных частью оказались раздробленными и полностью разрушенными, а частью могли быть отнесены течением дальше и погребены в другом месте. То же относится и к пустотелым или очень пористым, легким и хрупким костям птиц.
Грызуны и насекомоядные павлодарской фауны в первых сборах совсем отсутствуют; встречаются остатки черепах, но в виде отдельных щитков панциря, нет никаких следов ящериц и других позвоночных, несомненно входивших в состав фауны в целом. В коллекциях из Павлодара на сотни индивидов копытных проходятся единицы хищников.

Рис. 7. Обмазка костей глиной, смешанной с соломой. Павлодар, 1929 г.
Впрочем последнее можно объяснить и тем, что хищники всегда малочисленны по сравнению с огромными стадами травоядных копытных, кроме того, они могли захороняться в других местах. Таким образом, мы имеем типичный пример "выборочного" захоронения.
Самыми крупными животными гиппарионовой фауны были хоботные - мастодонты и динотерии. Однако материал по хоботным, добытый в Павлодаре, пока недостаточен для точного определения. Скудость остатков хоботных могла бы стоять в связи с их относительной малочисленностью сравнительно с жирафами, носорогами и особенно с гиппарионами, которые, вероятно, паслись в конце миоцена и в нижнем плиоцене на простоpax Западно-Сибирской равнины такими же табунами, как еще совсем недавно зебры и другие копытные в саваннах Африки. Однако современные слоны в Африке и в Южной и Юго-Восточной Азии до начала их истребления огнестрельным оружием и развития промысла "слоновой кости" (т. е. бивней самцов) жили в очень большом числе, да еще и сейчас многочисленны в некоторых африканских заповедниках. Поэтому вполне возможно, что малочисленность хоботных в Павлодаре только кажущаяся и объясняется но фактической малочисленностью животных и самой фауне, а захоронением их массивных черепов и костей где-то в другом месте, скорее всего, вверх по течению потока, образовавшего павлодарское кладбище. Не исключено и другое объяснение - хоботные могли избежать по каким-либо причинам массовой гибели, по-видимому, имевшей место для гиппарионов, жираф, оленей и других копытных.
Носороги павлодарской фауны (рис. 8, 9) - главным образом из безрогого рода хилотерий - с широким симфизным отделом нижней челюсти и большими торчащими из нее вперед и в стороны, косо сточенными со стороны симфиза резцами, напоминающими стамески. Хилотерий принадлежит к широко распространенному в миоцене - плиоцене Северной Америки, Азии и Восточной Европы семейству толстых, приземистых носорогов, коротконогих, с боченкообразным, как у бегемота, туловищем, с зачаточным рогом на конце носовых костей, о чем можно догадываться по совсем слабому развитию шероховатости для основания рога, иногда совсем безрогих. Очень характерны большие, торчащие вперед, нижние резцы хилотериев, которые, вероятно, помогали добывать растения (см. рис. 8). У современного крупного индийского носорога острые резцы играют роль оружия, подобно клыкам кабана, и он наносит ими тяжелые повреждения даже слонам. Тем более опасным оружием должны были быть мощные острые нижние резцы хилотерия.
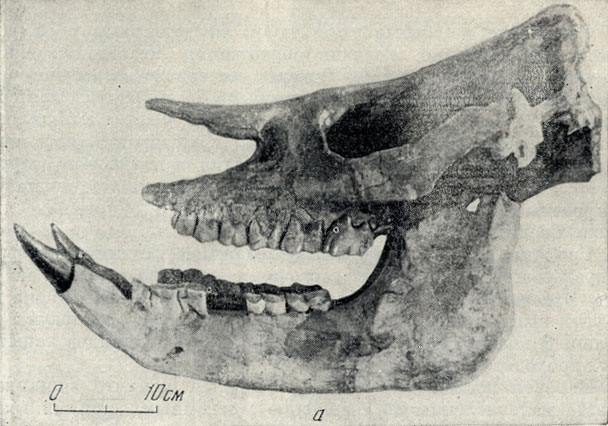
Рис. 8. Носорог хилотерий (Chilotherium) из Павлодара. а - череп
Крупные коренные зубы хилбтериев сложного строения, хорошо приспособлены для измельчения листвы, молодых побегов: деревьев и кустарников, возможно, и мягкой травянистой растительности (см. рис. 8). На молочных резцах встречаются как бь? насечки в основании коронки, пропиленные травой, продергивавшейся при пастьбе между зубами (известны и на зубах других копытных).
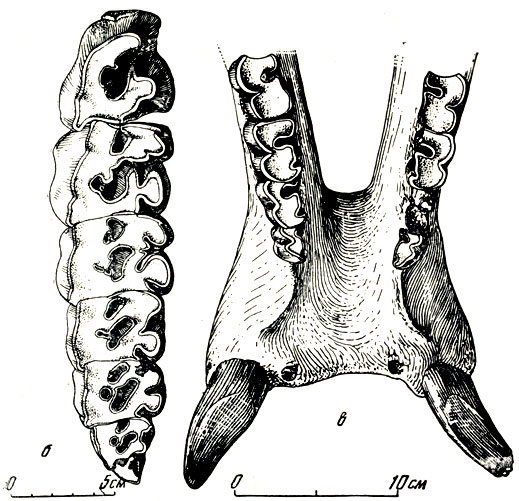
Рис. 8. Носорог хилотерий (Chilotherium) из Павлодара. б - верхние ложнокороенные и коренные зубы; в - жевательная поверхность и передний конец нижней челюсти (вид сверху) Павлодар. Палеонтологический институт АН СССР
По словам Е. И. Беляевой (устное сообщение), носорог павлодарской фауны близок к хилотерию, известному в гиппарионовой фауне Китая. Немногочисленные остатки какого-то очень крупного носорога из Павлодара, возможно, принадлежат синотерию, найденному ранее в Китае.
Большие массивные черепа и другие кости, естественно, сохраняются легче и в большем числе, чем остатки мелких животных; неудивительно поэтому, что в Павлодаре и в других местонахождениях гиппариоыовой фауны остатков носорогов всегда много "по объему и по весу".

Рис. 9. Носорог хилотерий (Chilotherium) Реконотрунция внешнего вида (рис. К. К. Флерова). Музей Палеонтологического института АН
Гиппарион, по числу найденных остатков и особей обычно преобладающий во всех крупных местонахождениях этой фауны, представлен и в сборах из Павлодара очень обильно (несколькими тысячами отдельных зубов, а всего - более 15 000s различных остатков). Помимо трехпалости гиппариона отличают от лошадей однопалых, доживших до современности, некоторые черты архаичности в строении черепа, менее развитая, чем у лошадей, затылочная связка, поддерживающая череп, относительно большая величина височной впадины; следовательно, относительно более сильный височный мускул, чем у лошадей (вероятно, черта примитивности); меньшая редукция клыков присуща животным обоего пола (у современных лошадей обычно имеются только у самцов). Очень типично строение верхних; коренных зубов: на не стертых до основания верхних коренныж внутренний "столбик" зуба ("протокон", предположительно БОЗНИК из первичного конуса простого одноконусного зуба) oобособлен от остального зуба, тогда как у лошади они слиты (рис. 10). Конечности гиппариона, по данным В. И. Громовой (1952), были более согнуты в суставах, менее выпрямлены, чем у лошадей. Гиппарион, по мнению Громовой, был функционально трехпалым животным; об этом говорит хорошая оформленность фаланг и суставных поверхностей на боковых пальцах, вообще, все их строение и ряд сравнительных наблюдений над oбоковыми пальцами современных копытных (косули, северные олени и др.), у которых они редуцированы.
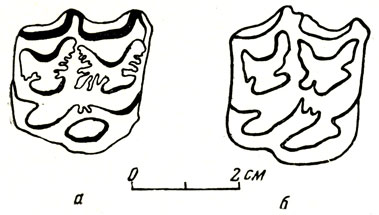
Рис. 10. Жевательная поверхность верхнего правого коренного зуба а - гиппариона (Hipparion elegans Grromova) из Павлодара (В. И. Громова, 1954) и б - лошади (Equus stenonis) (В. И. Громова, 1952)
Боковые пальцы гиппариона при беге по твердому субстрату, видимо, не касались земли, но должны были быть полезны при ходьбе по влажному, заболоченному грунту (возможно, и по песчаному), по скользкой тропе и т. п. Для гиппариона характерна способность боковых пальцев широко раздвигаться (что увеличивает площадь опоры стопы), сильная сгибаемость и отведение назад среднего пальца и боковых. Подобно северному оленю и лосю, у гиппариона более острые углы сгиба конечностей, чем у однопалой лошади (приспособленной для бега по твердому грунту). Эти особенности говорят, по мнению К. К. Флерова (1954), о явной приспособленности гиппариона скорее всего к влажным степям, широким речным долинам с богатой пойменной и луговой растительностью. Здесь могли быть и прибрежные леса, и участки лесостепи с кустарниками, и отдельные заболоченные места. Зубы гиппариона сравнительно с зубами однолалых лошадей имеют значительно более низкую коронку и приспособлены к более мягкой травянистой пище. чем у лошади. Все это, по мнению К. К. Флерова, свидетельствует, что гиппарион проводил большую часть своей жизни на мягком влажном грунте, как это предполагает и В. И. Громова для малого гиппариона из Павлодара (видовое название элегантный - Н. elegans Gromova) - тонкого, стройного и относящегося к числу мелких видов, ростом с осла. Для малого гиппариона, жившего на территории нынешних Западной Сибири и Казахстана большими табунами, характерна очень сильная складчатость эмали коренных зубов, малая разделенность коронки цементными прослойками, следовательно, и большая прочность зубов, и, видимо, прекрасная приспособленность их к перетиранию травянистой растительности. (В этом отношении; гиппарион напоминает современную лошадь, хотя и уступает ей.)
Другой крупный !вид из Павлодара - длинноногий гиппарион (найдены остатки не менее 9 особей) - был не только крупнее, но и относительно более долгоногим и достигал размеров средней лошади. В нем: сочетаются очень высокий для гиппариона рост с относительно длинными и тонкими ногами; он отличается этим от всех остальных известных гиппарионов, которые-либо много меньше (большинство), либо, если крупнее, то с массивными конечностями.
Жирафы павлодарской фауны обладали умеренно вытянутой шеей, лишь слегка удлиненными ногами и, следовательно, более или менее тем же обликом, что и современные окапи лесов Центральной Африки, сравнительно недавно (1900) ставшие известными зоологам. Родина современных жираф, вероятно, Азия, давшая большой и разнообразный материал по ископаемым жирафам (Китай, Монголия, Индостан). Запутанная в сложная систематика ископаемых жираф подверглась обстоятельному пересмотру Кольберта (1935) и Крюзафон-Пайро (1952) (рис. 11)., По-видимому, все жирафы, ископаемые и современные, могут быть разделены на три подсемейства: 1) наиболее архаичные в своем строении палеотраги, куда относится "живое ископаемое" - окапи, в некоторых отношениях самая примитивная из всех известных жираф (даже сравнительно с наиболее древними известными верхнемиоценовыми); 2) жирафы собственно - современные длинноногие африканские жирафы с длинной шеей и сильно шжатой назад спиной, а также и немногие ископаемые (Орасиус); 3) ископаемые неогеновые (или нижнечетвертичные) сиватерии - куда, кроме самого "сиватерия", первоначально найденного в Сиваликских холмах Индии, а сравнительно недавно и на Балканском полуострове, относятся "браматерий", "вишнутерий" (Сива (Шива), Брама, Вишну - божества индусской мифологии) и др. Это в большинстве массивные, буйволообразные жирафы, самые большие парнокопытные, но имеющие на первый взгляд ничего общего с жирафами в обычном понимании этого слова: В павлодарской фауне пока обнаружены два рода из палеотрагов. Один из них - самотерий (Samotherhim, впервые найденный на о-ве Самосе). К самотерию же относятся первоначально выделенные в самостоятельные роды и геологически приблизительно одновозрастные жирафы, описанные под названием альцицефалус из Ирана (Matpara) и "херсонотерия" из Ново-Елизаветовки на юге Украины (А. К. Алексеев, 1915). Другая павлодарская жирафа принадлежит к роду палеотрагус, также широко распространенному в составе пикермийской фауны (рис. 12). Палеотрагус известен те китайской фауне гиппариона (Bohlin, 1927), где имеется вид, очень близкий к сибирскому, цели не тот же самый. На территории СССР палеотрагус был описан под названием ахтиария (название по деревне Ахтиар) из верхнесарматских отложений Севастополя (А. А. Борисяк, 1915) и Эльдара в Закавказье (А. К. Алексеев, 1930).
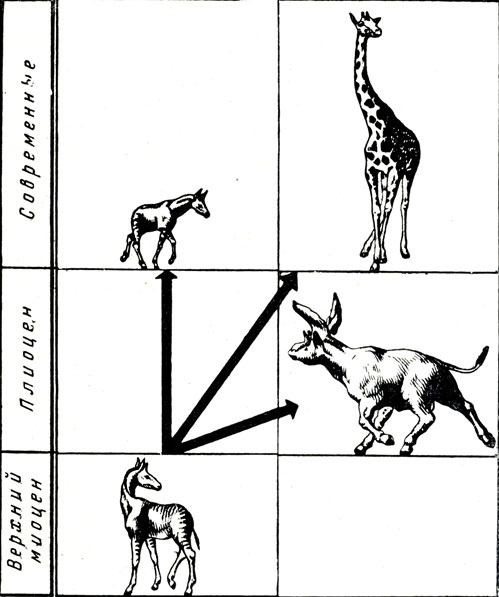
Pис. 11. Схема эволюции жираф (по Э. Кольберту) Верхний миоцен - нижний плиоцен, палеотрагус (Palaeotragus).. Плиоцен, сиватерии (Sivatherium). Современные окапи (Okapia) и камелопардалис (Camelopardalis)
В павлодарской фауне пока не обнаружено сиватериев. Некоторое указание на их распространение в Сибири очень крупный астрагал, найденный на одном из алтайских рудников и хранившийся долгие годы в Семипалатинском музее. Вероятно, к сиватериям относятся некоторые кости конечностей какого-то огромного парнокопытного из Касякинского карьера около Ставрополя Кавказского, хранящиеся в Палеонтологическом институте АН СССР. Не найдено в Павлодаре, возможно случайно, и длинноногих жираф из подсемейства Giraffinae, дожившего до современности в Африке, а в ископаемом состоянии они известны из Греции (орасиус) и Китая (хонанотерий).

Рис. 12. Жирафы рода палеотрагус (Palaeotragus) Реконструкция внешнего вида (рис. К. К. Флерова). Музей Палеонтологического института АН СССР
В павлодарской фауне довольно многочисленны остатки oоленей, относящиеся к роду цервавитус, впервые установленному И. П. Хоменко (1913) для Бессарабии и широко распространенному в гиппарионовой фауне от Бессарабии до Китая (К. К. Флеров, 1950). В отличие от бессарабского вида, обладавшего ребристыми рогами, у павлодарского, как и у близкого к нему китайского, описанного Зданским (О. Zdansky, 1925), рога гладкие. "Лопата" на рогах павлодарского оленя слабая, с тремя-четырьмя отростками (рис. 13). Характерно, что при относительной многочисленности рогов остатки черепа и зубов павлодарских оленей очень редки; видимо, кость рота оружие боя у оленей значительно прочнее остальных костей оленьего скелета.
Из антилоп имеются остатки трагоцероса (рис. 14), довольно крупной, антилопы, близкой к виду Tragoceros amalteus, известному и Греции и Малой Азии. Павлодарская газель неотличима от газели, известной из Пикерми и других местонахождений Балканского полуострова, Малой Азии и Казахстана (К. К. Флеров, 1935). Газель из гшшаржшовой фауны Китая, по данным К. К. Флерова, вряд ли заслуживает выделения в особый вид, отличный от газели с Балканского полуострова и Малой Азии.
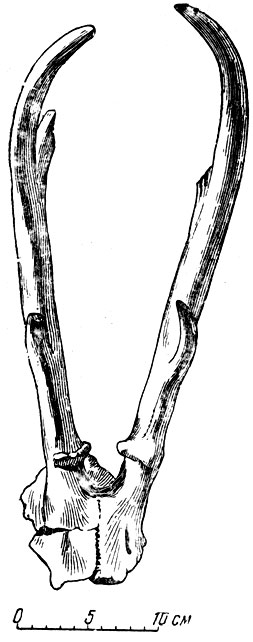
Рис. 13. Часть черепа с рогами оленя цервавитуса (Cervavitus orlovi Flerov). Павлодар
В павлодарской фауне не обнаружено никаких следов обезьян, встреченных в более южных местонахождениях гиппарионовой фауны (Греция, Закавказье, Бессарабия, Индия). Возможно, что причина этого отсутствия кроется не только в условиях захоронения в Павлодаре, неблагоприятных для хрупких костей обезьян, но и в северном положении павлодарского местонахождения, вместе с ишимскими (у Селим-Джевара и на р. Кара-Кол по среднему течению р. Интим) одного из самых северных местонахождений этой фауны. Не найдены и свиньи, обычные в гиппарионовой фауне.
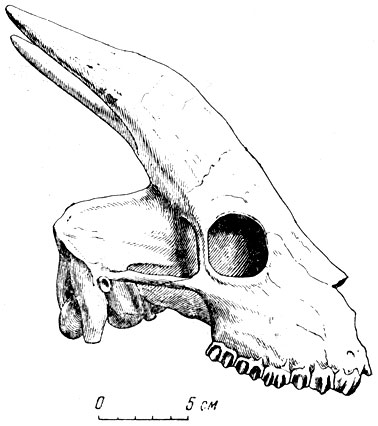
Рис. 14. Часть черепа трагоцероса (Tragoceros amalteus Gaudry), антилопы фауны гиппариона?из южноевропейских местонахождений
Многочисленные растительноядные млекопитающие павлодарской фауны, в частности копытные, служили обильной пищей разнообразным хищникам. Самым грозным из них, несомненно, был очень крупный саблезуб - махайрод (рис. 15). Судя по уцелевшей от него большой массивной нижней челюсти, это один из самых крупных известных махайродов. По сравнению с тигром (и львом) крупные саблезубы отличаются более длинной мордой с огромными кинжалообразными уплощенными верхними клыками, могучей шеей, мощными передними конечностями при относительно слабых задних, явно не приспособленных для прыжков и нагона добычи (как это имеет место у тигра и льва). Видимо, саблезубы представляли экологический тип, резко отличный от "настоящих кошек".. Наряду с очень большими размерами павлодарскому саблезубу присущи некоторые черты архаического характера, например меньшая, чем у других махайродов, специализация послеклыковых зубов. Небольшие остатки близкого по размерам махайрода были найдены на р. Дженама вместе с зубом нижнеплиоценового "мастодонта Бореона", описанным М. В. Павловой. Вообще говоря, саблезубы, образующие особое вымершее подсемейство кошек, включали в себя, подобно современным кошкам, не только очень крупных животных размеров тигра, но и меньших - величиной с леопарда, рысь и еще более мелких. В павлодарской фауне, кроме огромного махайрода, обнаружены кошки величиной примерно с рысь и с дикого кота.

Рис. 15. Саблезуб (рис. К. К. Флерова)
Из других хищников самым сильным была гиена из рода крокута (см. рис. 16, б), по скелету сходная с современной африканской пятнистой гиеной (рис. 16, а) и известная из местонахождений гиппарионовой фауны во Франции, Германии, Венгрии, Греции, Бессарабии, на Эгейских островах, в Малой Азии. Эта ископаемая крокута все же отлична от современных гиен. Верхний плотоядный зуб (Р4), обычно у хищников трехбугорчатый, превращен у нее в двухбугорчатое "лезвие"; другие отличия от современной крокуты - длинноногость, меньшая, чем у современных гиен, укороченность задних ног сравнительно с передними, значительная длина пяточной кости, что должно было делать гораздо более сильным сокращение икроножных мышц (следовательно, более быстрый бег), большая стройность животного. Известно, что современная гиена не только поедает падаль и нападает на слабых и больных животных, включая льва, но нередка, соединившись в небольшие стаи, не прочь преследовать и здоровую добычу. Тем больше оснований считать очень активным и подвижным хищником стройную и проворную крокуту гиппарионовой фауны. Возможно, лишь местной расой этого вида является и очень крупная гиена, описанная Зданским (1924) из содержащих гиппарионовую фауну красных глин Китая. Таким образом, ареал распространения крокуты гиппарионовой фауны был очень велик - от Франции до Восточного Китая и от южной Греции до Сибири.

Рис. 16. Крокута. а - современная пятнистая гиена (Crocuta crocuta Zimmermann), Южная и Восточная Африка
Из остальных хищников в павлодарской фауне преобладает иктитерий, известный из местонахождений западной и юго-восточной Европы (Сицилия, Венгрия, Балканский полуостров) и Ирана (рис. 17). "Иктитерий" (греческое иктис - лисица, oтэр - зверь) относится одними авторами к виверрам, другими - к гиенам. Действительно, эти хищники, закончившие свое существование в илиоцене, во многам промежуточны между виверрами и гиенами. Наиболее мелкий вид настолько виверрообразен, что его правильнее отнести к семейству виверр (рис. 18); что же касается крупного, гиппарионового иктитерия, то изучение павлодарского материала и других сборов (Pilgrim, 1931, Ю. А. Орлов, 1938) показывает с несомненностью, что этот вид следует считать гиеной, но менее специализированной на пути к крупным: гиенам типа пятнистой, чем упомянутая ископаемая крокута; гиппарионовый иктитерий обнаруживает сходство с ней в строении конечностей. Стройный, тонконогий гиппарионовый иктитерий достигал размеров мелкого степного волка и при своей подвижности мог быть еще более активным охотником, чем крокута. На шлифах копролитав иктитерия хорошо сохранились куски оставшихся непереваренными костей млекопитающих с костными канальцами и щелями, заполненными окислами железа.
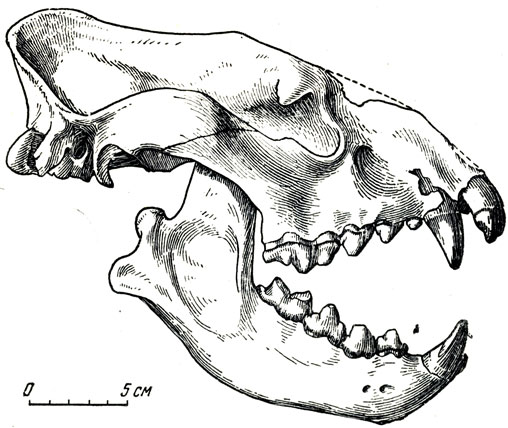
Рис. 16. Крокута. б - череп Crocuta eximia Roth. et Wagner. Павлодар. Музей Палеонтологического института АН СССР
Из семейства куниц в павлодарской фауне с достоверностью найдены росомаха, барсук и куница (Ю. А. Орлов, 1941). Росомаха Павлодара мельче описанной Зданским (1924) в фауне из Китая, но принадлежит тому же короткомордому роду и виду плезиогуло, что и найденные в гиппарионовой фауне Китая и Индии.
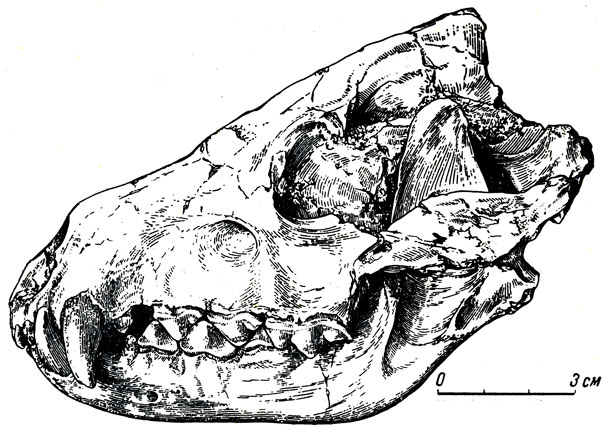
Рис. 17. Инкитерий (Ictitherium hipparionum Gervais). Деформированный неполный череп. Павлодар (Ю. А. Орлов, 1936)
Барсук павлодарской фауны тот же, что и в пикермийской фауне Китая (Зданский, 1924) и Ирана (Kittl, 1887), выделенный Зданским в род паратаксидэа ввиду сходства его черепа с черепом современного короткомордого североамериканского барсука таксидэа (рис. 19). В павлодарском материале оказались и конечности барсучьего типа (ранее были известны только череп и нижняя челюсть), несущие все черты приспособления к рытью, хотя и в меньшей степени, чем у современного барсука.

Рис. 18. Современная виверра
Куница павлодарской фауны неотличима от описанной Зданским в качестве нового вида. В описании Зданского нет признаков, которые подтверждали бы правильность выделения в особый вид, отличный от современной лесной куницы. По-видимому, куштца из гиппарионовой фауны Сибири и Китая, если

Рис. 19. Паратаксидэа (Parataxidea crassa Zdansky). Реконструкция К. К. Флерова
В итоге обзора следует упомянуть о семанторе, своеобразном представителе семанторид, нового семейства ластоногих (Орлов, 1933, рис. 20 и 21). От семантора найдена лишь задняя половина скелета, изучение которой убеждает в промежуточном положении этого ластоногого, с одной стороны, между выдрообразными куницами, с другой - тюленями; это подтверждает предположения Майварта (Mivart, 1862) о возможности происхождения тюленей от куниц. Тем не менее сам семантор имеет особенности строения, исключающие его из прямых предков тюленей, хотя R общем и мог бы служить одним из примеров "связующего звена" (семантор найден слое № 1, вниз по течению от главного местонахождения).
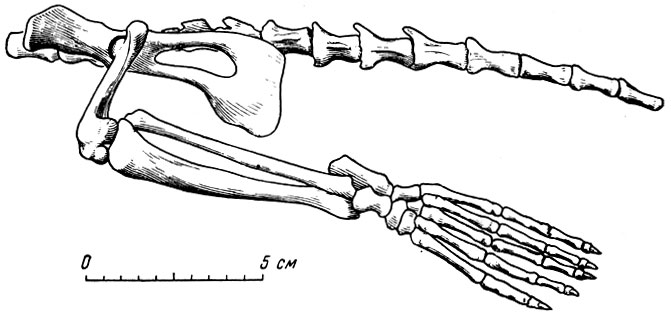
Рис. 20. Семантор (Semantor macrurus Orlov) Часть скелета. Павлодар (Ю. А. Орлов, 1933)
В Павлодаре обнаружены остатки черепах, по-видимому, из триониксов (кожистых трехшготных). Из птиц А. Я. Тугариновым (1935) описаны незначительные остатки крупного страуса, предположительно одинакового с "херсонским страусом", описанным в Бессарабии (А. Брандт, 1873), и нового школа сушкиния, близкого к одному из современных африканских соколов и названного в честь академика П. П. Сушкина.

Рис. 21. Семаптор (Semantor macrurus Orlov) Реконструкция (рис. А. П. Быстрова) (Ю. А. Орлов, 1933)
В последнее время (письменное сообщение В. С. Бажаяова, 1960) работниками Академии наук Казахской ССР путем промывания через сита пород среднего и нижнего горизонтов в местонахождении на "Гусином перелете" обнаружены остатки зайца, пищухи, мышовки, тушканчика, змеи, земноводных (типа жабы и лягушки-чесночницы) и мелких птиц.
Помимо местонахождения у "Гусиного перелета" (ныне У железнодорожного моста), остатки той же фауны, перемешанные с комьями пятнистой глины, часто включенные в них, были встречены невдалеке от раскопок вниз по течению, во вторичном залегании, вместе с костями четвертичных млекопитающих, а также рассеяны по отмелям и побережью островов вниз по Иртышу.
Большое количество костного материала павлодарской фауны погибает в течение многих десятков лет при ежегодных разливах Иртыша. Тем не менее и до сих пор местонахождение далеко не "исчерпано". Очевидно, общее количество скелетного материала, когда-то захороненного в неогеновой песчано-глинистой толще Павлодара, было огромно и, вероятно, соответствует тысячам особей, главным образом копытных, которые, может быть, стали жертвой стихийного бедствия.
Трудно сказать, чем была вызвана эта гибель. Причиной могли быть: внезапное наводнение, вызванное ливнями и сопровождавшееся ураганом; большой лесной пожар от удара молнии в засушливое лето, перекинувшийся на степь; длительная катастрофическая засуха, при которой животные гибнут не только от недостатка влаги и пищи, но и губят друг друга, толпясь на водопое, а тяжеловесные копытные погибают, увязая в топком прибрежном иле; не исключена была и смерть многих животных, особенно копытных, живших стадами, от эпизоотии. Наконец, большие разливы древней реки, так сказать "пра-пра Иртыша", могли время от времени омывать с лугов и побережий реки скелетные остатки жившей по его берегам фауны.
Если считать, что основная часть захороненного материала представляет все же итог массовой единовременной гибели животных, то можно попытаться сделать догадку о времени года, когда погибли, по крайней мере, те животные, от которых известен молодняк. Такая попытка была, например, сделана одним автором для местонахождения гиппарионовой фауны, где можно было установить примерный возраст жеребят гиппариона, допустив, что период размножения его и современной однопалой лошади совпадают, т. е. в основном время рождения большинства жеребят обоих родов лошадиных одно и то же. Для павлодарской фауны такой приблизительный расчет пока еще не сделан и число жеребят со сменой зубов незначительно. По мнению К. К. Флерова (1950), можно предположить, что массовая гибель оленей павлодарской фауны произошла во второй половине лета, так как найденные в Павлодаре остатки рогов представляют не спавшие рога, а сидящие на выростах лобных костей и вполне сформировавшиеся, окостеневшие, т. е. очищенные от "бархата" (покрытой шерстью кожи).
Следовательно, животные погибли, вероятно, незадолго до периода спаривания, который у "оленьков" и у наиболее примитивных оленей (цервавитуса, мунтжаков и элафодусов) происходит летом; можно предположить, что период спаривания у цервавитуса, еще очень близкого к мунтжакам, приходился также на вторую половину лета, а не на осень, как у большинства современных оленей.
Само павлодарское местонахождение, очевидно, лишь место захоронения перенесенных водою костных остатков, но не место гибели всей этой массы животных. В состав животных этой фауны, попавших в захоронение, входят и виды, широко распространенные в Европе и Азии в конце миоцена или нижнем плиоцене (гиена, иктитерий, газель) и "чисто азиатские" (паратаксидэа) и, возможно, "чисто Западно-Сибирские", эндемичные (махайрод, семантор).
Однако общий характер фауны очень типичен для описываемого времени. Некоторые "чисто-азиатские" представители этой фауны в последнее время, по-видимому, обнаруживаются в Европе. Некоторые азиатские виды при ближайшем рассмотрении, быть может, придется принять тождественными с ранее известными европейскими, а вместе с тем сделать вывод о несколько большем однообразии фауны на обширных лесостепных и степных пространствах Европы и Азии, чем казалось ранее (рис. 22).
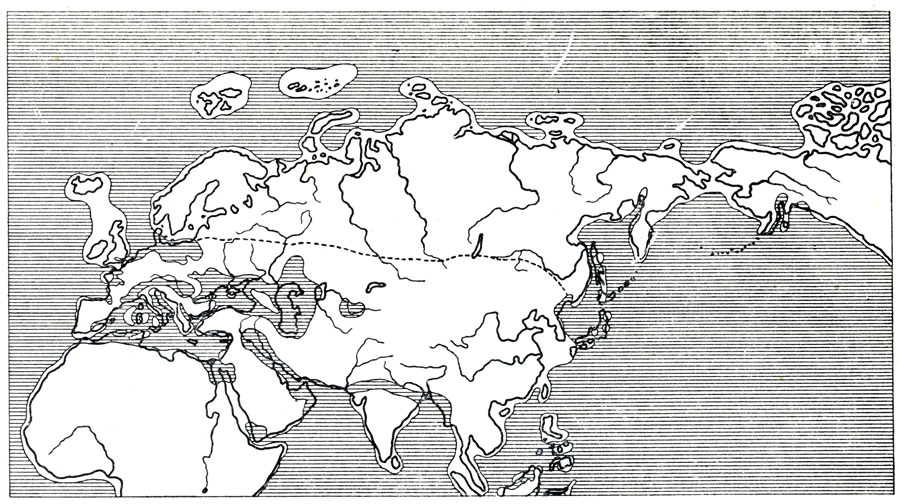
Рис. 22. Северная граница (пунктирная линия) известных местонахождений фауны гиппариона и некоторые палеогеографические связи (по Термье, 1952) в конце миоцена-нижнем плиоцене
В настоящее время ареалы "свободного от человека" обитания диких животных очень сужены, часто до пределов заповедников, правда, в некоторых случаях огромных (Африка). Обширные степные пространства Азии, саванны Африки и Южной Америки уходят безвозвратно под земледелие и скотоводство; многие миллионы голов диких копытных (Еще во второй половине прошлого века в США за несколько лег было перебито около 4000000 бизонов. По данным современных американских и французских журналов, в Африке ежегодно убивают около пяти тысяч слонов) погибли от огнестрельного оружия, а некоторые виды млекопитающих: и многие птицы исчезли с лица земли, т. е. истреблены человеком. Нам уже становится трудно представить себе массовость жизни, т. е. многочисленность многих животных в прошлом. Еще сравнительно недавно некоторые газели в Азии крупные антилопы, газели и зебры в Африке производили свои перекочевки на новые пастбища или сезонные откочевки десятками, а то и сотнями тысяч голов, так что за километр и более путешественник слышал запах этих гигантских диких гуртов, которым могли бы позавидовать все скотоводы мира (рис. 23-26). И мы можем лишь по аналогии с этой "вчерашней современностью" догадываться о той огромной "биомассе", которая была и "прошла" через территорию ныне Западной Сибири и Казахстана несколько миллионов лет тому назад, во времена фауны гиппариона. Разумеется, эта фауна с течением времени меняла свой облик тем более, что она была распространена от Испании до Тихоокеанского побережья Китая и от Центральной Африки до Западной Сибири и Забайкалья. Но, именно Центральная и Южная Азия, по мнению многих, были родиной фауны, так напоминающей современную африканскую, хотя отдельные члены фауны гиппариона могли быть и, видимо, были совершенно разного происхождения в смысле первичной родины их предков.

Рис. 23. Зебры (Equus burchelli Gray). Африка (М. Johnson, 1936)
Фауна гиппариона справедливо считается в значительной степени предковой для современной африканской, но ее формирование как комплекса животных разного палеогеографического происхождения не совсем ясно. Некоторыми учеными считалось, что родина этой фауны Центральная и Южная Азия, где представлены крупные местонахождения (Китай, Монголия. Казахстан, Сибирь, Индия и прилегающие острова). Возможно, что именно на территории Центральной и Южной Азии сформировался этот поразительно характерный и в общем однородный "пра-африканский" комплекс, откуда фауна и расселилась в Европу, Переднюю Азию и Африку. Но пока это все же скорее лишь наполовину обоснованная гипотеза, хотя наибольшее разнообразие, например, ископаемых жираф характерно именно для Азии. Однако история палеонтологии указывает на необходимость большой осторожности в этих вопросах, хотя для некоторых групп млекопитающих не приходится сомневаться в том, на каком материке они впервые появились.

Рис. 23. Зебры (Equus burchelli Gray). Африка (М. Johnson, 1936)
Ландшафт, в котором жила фауна гиппариона, в общем должен был носить лесостепной характер; это можно допускать и для Западной Сибири и Северного Казахстана в описываемое время. Но отчетливых данных офлоре Западной Сибири для этого времени мы не имеем. Лишь одно ясно - широкое распространение фауны, ее "теплолюбивость", казалось бы, скорее всего субтропический в общем характер (за 'исключением самых северных местонахождений). Широкое распространение фауны может говорить не столько об однообразии климата для такой большой территории, сколько о широком распространении сходных ландшафтов; при этом могло бы идти относительно быстрое расселение разных будущих (Компонентов фауны из мест возникновения в другие ареалы, и вся, в целом однообразная, фауна могла бы относительно быстро возникнуть на весьма обширной территории "почти одновременно" - в геологических масштабах времени. Разумеется, это допущение не противоречит разнородности фауны в смысле происхождения отдельных ее представителей. Не вполне ясен возраст сибирско-казахстанской фауны; скорее всего - это понт. За последние десятилетия были сделаны попытки, по-видимому, достаточно обоснованные, приурочить разные фауны гиппариона из их многочисленных местонахождений Европы, Азии и Африки к разным по возрасту отложениям, хотя еще недавно некоторыми исследователями указывалось на необоснованность этого подразделения и на то, что пока правильнее говорить о единой фауне млекопитающих верхнего сармата - понта. Однако за последние годы накопились данные, говорящие о нескольких исторических этапах фауны гиппариона, от верхов среднего миоцена до конца нижнего плиоцена; позднее сам гиппарион еще встречается, но вся эта "пра-африканская" фауна уцелевает в сильно измененном виде лишь в самой Африке, где ее прямые потомки живут и поныне. Не совсем бесспорно и происхождение гиппариона, обычно считаемого потомком северо-американской трехпалой лошади мерикгиппус. По мнению же некоторых (Пилгрим, 1913), гиппарион ведет начало от более древних лошадей Старого Света (возможно, неизвестного еще азиатского мерик-гиппуса). Против этого категорически возражают Мэтью (1929) и Кольберт (1935), указывающие на то, что североамериканский гиппарион, происходящий от североамериканского же мерин-гиппуса, имеет все основания считаться непосредственным предком гиппарионов Старого Света. Вместе с тем, по мнению этих авторов, гиппарион своим внезапным появлением и быстрым распространением в Старом Свете знаменует наступление нового исторического момента в большей степени, чем почти вся остальная часть фауны, главным образом азиатского происхождения. Таким моментом, по мнению Кольберта, является граница миоцена и плиоцена.

Рис. 24. 'Лошадинная антилопа' (гну) Центральной Африки (Gordon taurinus Burcbell) На верхнем снимке небольшая часть стада 'на марше на лугах в кратере потухшею вулкана Нгоронгоро. Точки вдали - другое большое стадо. На нижнем - стадо, сфотографированное с испугавшего их самолета (M. Johnson? 1936)
По мнению В. И. Громовой (1952), род гиппарион, по-видимому, возник в Северной Америке из среднемиоценового мерик гиппуса; в Старом Свете в среднем миоцене до сего времени но обнаружено рода, который мог бы считаться предком гиппариона; последний возник, вероятно, в самом верхнем миоцене и быстро расселился в Старый Свет. Черты строения самых ранних гиппарионов Старого Света, по мнению В. И. Громовой (1952), являются показателями влажной обстановки и облесенных ландшафтов, свойственных умеренным широтам. Поэтому естественно предположение о сравнительно северных путях миграции гиппариона из Северной Америки в Старый Свет. (С этим согласуется и тот факт, что одновременно в Америку из Старого Света не проникли типично теплолюбивые жирафы, антилопы, гиены, а из Северной Америки в Азию - верблюды, склонные к сухому климату.) Следует считать, что и в уточнении истории развития самих гиппарионов Старого Света и всей фауны гиппариона большую роль будет играть изучение третичных млекопитающих территории СССР, Китая, Индии.

Рис. 24. 'Лошадинная антилопа' (гну) Центральной Африки (Gordon taurinus Burcbell) На верхнем снимке небольшая часть стада 'на марше на лугах в кратере потухшею вулкана Нгоронгоро. Точки вдали - другое большое стадо. На нижнем - стадо, сфотографированное с испугавшего их самолета (M. Johnson? 1936)
Пока же наши сведения о фауне хотя в целом обильны, но в сущности очень неполны. В ее состав входили млекопитающие, птицы, пресмыкающиеся, земноводные, не говоря уже о множестве сухопутных и водных моллюсков, ракообразных, насекомых и других беспозвоночных. О них пока мы можем лишь догадываться. Выморочность захоронения костных остатков и неполнота геологической и палеонтологической летописи вообще всегда ставят препятствие палеонтологу в его стремлении восстановить возможно полнее жизнь прошлого...

Рис. 25. Современные копытные Центральной Африки На верхнем снимке (а) - 'чернопятая газель' импала (Aepyceros melampus Lichtenstein) на водопое. На нижнем снимке (б) - африканские буйволы на водопое (из 'Way of the wild', 1958)
В последнее время Куртэном (1952) сделана очень интересная попытка разделения гиппарионовых фаун на лесные, степные и смешанные, основанная на изучении газелей и некоторых других млекопитающих этой фауны.

Рис. 25. Современные копытные Центральной Африки На верхнем снимке (а) - 'чернопятая газель' импала (Aepyceros melampus Lichtenstein) на водопое. На нижнем снимке (б) - африканские буйволы на водопое (из 'Way of the wild', 1958)
Каким бы путем ни сложилась фауна гиппариона, "пришла" ли она из Азии в Африку, как считается многими, или на Африки (Необходимо принять во внимание относительно слабую изученность ископаемых фаун Африки сравнительно с Европой и Азией. Некоторые млекопитающие гиппарионовой фауны, например трубкозуб (Orycteropus), известный из европейских местонахождений, по-видимому, заведомо африканского происхождения) в Азию, или, наконец, формировалась одновременно (или почти одновременно) на широких пространствах Евразии и Африки, связанных в прошлом шире, чем теперь, - ее "африканский" общий облик всегда бросался в глаза, и невольно ставил и ставит вопрос о климате, в котором жила фауна. Этот вопрос решается в общем просто для местонахождений Южной Азии, Южной Европы и самой Африки, где теплый климат начинается с неогена. Не так ясно дело с Западно-Сибирской равниной и вообще с Сибирью, где находятся самые северные из обнаруженных местонахождений. Ископаемая флора (если не считать неопределимых остатков) не обнаружена ни в сибирских, ни в других местонахождениях гиппарионовой фауны, открытых позднее в Казахстане и Киргизии, и о растительности во времена павлодарской фауны пока приходится лишь строить догадки по флорам из других мест - то более древним, то более поздним, чем, например, фауна Павлодара на Иртыше.

Риc. 26. Стада жираф (верхний рисунок а) и слонов (нижний - б) в Центральной Африке (М. Johnson, 1937)
Можно принять во внимание имеющиеся данные о более древних, чем фауна гиппариона, неогеновых флорах Северной и Средней Азии и о более поздних (приблизительно верхнеплиоценовых). Имеются интересные сведения и о том, что многие палеогеновые и неогеновые теплолюбивые растения Северной Азии нашли себе - с наступлением похолодания в конце плиоцена и в четвертичном периоде - "убежища" на Дальнем Востоке, в Южном Китае, на атлантическом побережье Северной Америки (П. Н. Дорофеев, 1957). Некоторые современные растения этих территорий - остатки той теплолюбивой флоры, которая известна в ископаемом состоянии в неогеновых отложениях Западной Сибири и, вероятно, входили в состав ландшафта, частью которого была фауна гиппариона; многие из современных южнокитайских растений, очевидно, со временем будут открыты в Западной Сибири или Казахстане в ископаемом состоянии.

Гиппарионовая фауна. Рис. К. К. Флерова
Трудность точного восстановления ландшафта, в котором обитала сибирская фауна гиппариона, заключается и в большей геологической длительности существования некоторых растений, чем многих ископаемых млекопитающих. (Впрочем, и сама фауна гиппариона известна на большом промежутке времени, измерявшемся по крайней мере многими сотнями тысяч лет, вернее несколькими миллионами). К тому же и относительно геологического возраста ряда сибирских третичных флор у палеоботаников пока нет единого мнения. Все же во времена гиппарионовой фауны можно предполагать для обширной территории Западной Сибири, Казахстана (вероятно, и прилегающих участков Азии) довольно мягкий климат и вместе с тем известное разнообразие ландшафта, несмотря на меньшую, чем в настоящее время, расчлененность Азии горными хребтами, ибо многие современные хребты моложе гиппарионовой фауны, так как возникли ъ итоге более поздних горообразовательных процессов. Последние могли быть одной из основных причин климатических изменений и смены физико-географических условий, следовательно, и изменения растительности, что должно было в свою очередь неизбежно отразиться на животном мире, вызвать "отступание" гиппарионовой фауны на юг или по крайней мере сокращение огромного ареала его обитания в Азии. Растительность Западной Сибири и Северного Казахстана в эпоху гиппарионовой фауны должна была быть очень разнообразной и включать, судя по упомянутым выше более ранним и более поздним флорам, многие хвойные и широколиственные породы, но в значительной степени листопадные, а также многочисленные травянистые растения. В совокупности вся эта флора кормила огромные стада мелких и крупных копытных. В лесных массивах росли болотные кипарисы, метасеквойя с опадающей хвоей, открытая сначала в ископаемом виде в Японии, позднее как "живое ископаемое" в Китае, а недавно в ископаемом виде в Западной Сибири (на реках Оби и Тавде). Для неогеновых отложений Западной Сибири известно также несколько видов сосны. Из неогеновых отложений Казахстана, Киргизии (и Туркестана) известны тростник и многие листопадные деревья и кустарники: дуб, бук, ясень, вяз, ольха, различные виды тополя, крушина, лещина, граб, грецкий орех, клен, плакучая ива, черная бузина, кизильник, спирея, ликви-дамбр.
Вряд ли приходится сомневаться в наличии в "гиппарионовое время" всех этих и многих других пород, в том числе различных лиан, в лесах Западной Сибири и Северного Казахстана.
Данные о травянистой растительности пока совсем скудны; известны лишь не поддающиеся точному определению отпечатки злаков. Можно предполагать, что и травянистая растительность была богата (иначе на чем паслись бы газели, разнообразные антилопы и др.), да и древесные породы, несомненно, были гораздо разнообразнее. По мнению П. Н. Дорофеева (1957), западносибирские третичные флоры рисуют нам картину лесной растительности, умеренно теплолюбивой, свойственной умеренно теплым странам, скорее всего с муссонным или ему подобным климатом, при котором обычны сезонные чередования сухих континентальных и влажных морских ветров.
Листопадность большого числа широколиственных древесных пород (из хвойных теряла на зиму хвою метасеквойя) и, возмож, но, довольно прохладные зимние периоды могли вызывать значительные, на многие сотни километров, откочевки копытных гиппарионовой фауны на юг, а за ними двигались кормившиеся копытными активные хищники и падалеяды, в том числе круп ные хищные птицы - грифы и т. д.
С наступлением жаркого летнего времени и засух могли иметь место откочевки и в обратном направлении, далеко на север: путь с юга на север, вероятно, был открыт до самого моря - ни горных хребтов, ни рек широтного направления в то время в Западной Сибири, очевидно, не было; возможности перекочевок скорее всего регулировались климатическими условиями и характером лесов и растительности в целом.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'