
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Глава четвертая. Ископаемые позвоночные Монголии
О прекрасная пустыня, Любимая моя другиня... Безмолвная и непразднословная Безропотная, нестроптивая
(Старинный стих пустынников)
Азия, ты свет жизни...
(П. Б. Шелли)
Монгольская Народная Республика расположена довольно-высоко над уровнем моря (в среднем 700 м и выше) я покрыта в южной части почти безводными, выгорающими летом степями и глинистыми полупустынями.
Горные породы в этой "гобийской" части Монголии нередко почти лишены растительного и почвенного покрова, легко разрушаются ветрами и атмосферными осадками. В полупустынной обстановке Гоби при резко континентальном климате это приводит к развитию глубоких оврагов, прорезающих сетью обширные участки древней суши, которую море не покрывало с нижнемелового, а местами и с верхнеюрского времени. Овраги особенно развиты в глинисто-песчанистых отложениях, заполнявших обширные межгорные котловины в мезозое и кайнозое. Все эти особенности ландшафта и геологического строения Монголии выяснили в конце прошлого и начале настоящего века наши замечательные географы и геологи Н. М. Пржевальский, Г. П. Потанин, В. А. Обручев, Г. Е. Грум-Гржимайло, А. А. Чернов. В. А. Обручеву принадлежит и первая находка (в 1892 г.) ископаемого позвоночного - носорога в юго-восточной части МНР. В Китае "кости и зубы драконов", т. е. ископаемых млекопитающих, были известны с древности и применялись для приготовления лечебных порошков; до самого последнего времени даже был особый промысел - добывание ископаемых костей и зубов как "лекарственного сырья".
Во втором десятилетии нашего века А. А. Борисяк на основании находок, сделанных экспедицией Академии наук в Казахстане, установил новые для Азии и всего Старого Света фауны третичных млекопитающих. Работы А. А. Борисяка вызвали большой интерес, были переведены и опубликованы в Западной Европе и Америке и в значительной степени послужили американским ученым путеводной нитью в поисках ископаемых позвоночных в Монголии в 1921-1930 гг. - в осадочных континентальных отложениях, сходных с казахстанскими. В 1941 г. Комитет Наук Монгольской Народной Республики обратился в Академию Наук СССР с просьбой организовать изучение ископаемых позвоночных в Монголии, но война помешала начать работы, В 1945 г., после вторичного приглашения Комитета Наук МНР, Палеонтологический институт АН СССР составил план экспедиционных работ в Монголии. Большие раскопки в отдаленных, безлюдных и безводных районах Гоби при бездорожье представляли сложное техническое предприятие и требовали серьезной подготовки и опытного руководства. Начальником экспедиции был профессор И. А. Ефремов (Палеонтологический институт АН СССР). В 1946 и 1948 гг. в экспедиции принял участие и автор этой книги.
В 1946 г. мы работали в Гоби сентябрь и октябрь ж обнаружили крупные местонахождения динозавров и других ископаемых позвоночных, что послужило основанием для последующих: раскопок и поисков.
Затем экспедиция работала с середины марта до половины; октября 1948 г. Недалеко от Далан-Дзадагада - основной базы экспедиции (юг МНР) - нами был найден новый интересный динозавр, а юго-западнее хребта Нэмэгэту скелеты хищных динозавров, остатки крупных растительноядных динозавров, целое кладбище водных черепах, окаменелая древесина. В нескольких километрах от этого места обнаружено местонахождение раннетретичных копытных и других позвоночных. Экспедиция осмотрела обширные древние котловины, заполненные отложениями верхнемелового возраста с остатками динозавров, собрала в разных местах многочисленные остатки шгжнетретичных млекопитающих.
В 1949 г. работы продолжались с мая до октября. На этот раз экспедиция добыла пресмыкающихся мелового периода, древних копытных - дштоцерат, а также верхнетретичных млекопитающих - мастодонтов, носорогов и др. Была обследована западная часть МНР в районе больших озер (рис. 27). Начиная с 1946 г., экспедиция содействовала организации в Улан-Баторе отдела палеонтологии в музее Комитета Наук МНР. Подробное описание полевых работ имеется у А. К. Рождественского - "За динозаврами в Гоби" (1957) и И. А. Ефремова - "Дорога ветров" (1957).
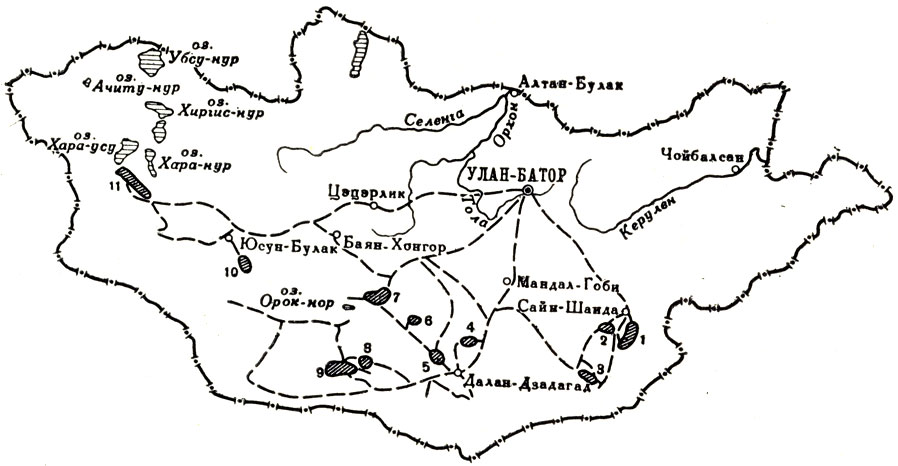
Рис. 27. Схематическая карта маршрутов и районов раскопок Монгольской палеонтологической экспедиция АН СССР в 1946-1949 гг. ---- маршруты; места со штриховкой - районы расценочных работ; 1 - Бэин-Ширэ; 2 - Хара-Хутул-Ула; 3 - Эргиль-Обо; 4 - Олгой-Улан-Цаб; 5 - Баин-Дзак; 6 - Оши-Нуру; 7 - Орок-Нор; 8 - Ширэгин-Гашун; 9 - Нэмэгэту; 10 - Бегер-Нур; 11 - Дзергенская котловина
Позднее мы узнали о некоторых новых находках. Вряд ли ными были обнаружены все местонахождения ископаемых позвоночных - пространства Гоби велики... Но нам удалось открыть крупные захоронения и собрать материал, представляющий и крупный научный интерес и большую музейную ценность. Экспедиция сделала не только палеозоологические сборы, но также интересные палеоботанические - отпечатки папоротников, окаменелую древесину магнолиевых, хвойных типа болотных кипарисов и других деревьев. В нижпемеловых отложениях сохранились вертикально стоящие ппи (до 2 м в поперечнике и более) целого леса, а в верхнемеловых - длинные стволы с сучками и годичными слоями, куски обгорелой древесины.
Наиболее крупные местонахождения мезозоя, около гор Нэ-лгэгэту, по данным экспедиции, образовались в области больших дельт, в их подводной части.
Местонахождения дельтового типа часто характеризуются хорошей сохранностью костного материала - здесь часты находки целых скелетов, так как нередко происходило захоронение трупов, приносившихся течением. Оно способствовало иногда скоплению трупов; поэтому на небольшом участке порой залегают несколько скелетов, например около горы Алтан-Ула. Другие местонахождения образовались, вероятно, в условиях более oспокойных вод - в озерах или старицах - и содержат менее полные, худшие по сохранности скелетные остатки.
Для кайнозоя (А. К. Рождественский, 1954, и И. А. Ефремов, 1950) можно отметить три типа местонахождений, образовавшихся: 1) в предгорных впадинах (Алтан-Тээли); здесь встречается огромное количество черепов и других различных частей скелета разных млекопитающих; 2) в дельтах рек или более кысокой части русел (Наран-Булак, Эргиль-Обо); эти местонахождения, заключающие обильные скопления отдельных костей и скелетов, как прашило, хорошей сохранности и 3) в озерах, oстарицах или больших болотах (Татал-Гол, Бэгэр-Нур), где значительное скопление костного материала образуют лишь мелкие животные, остатки крупных редки и встречаются в виде разрозненных частей скелета, реже - целых скелетов животных, увязших в трясине.

Академик А. А. Борисяк (1872-1944)
Сборы Монгольской Палеонтологической экспедиции Академии наук СССР происходят, по-видимому, из десяти "горизонтов жизни", начиная с нижнего мела и кончая плиоценом.
Многие пресмыкающиеся и млекопитающие, остатки которых oбыли нами собраны в Монголии, принадлежат новым видам и родам, иногда и семействам. Некоторые группы динозавров, как, например, игуанодоны, анкилозавры и некоторые млекопитающие, ранее не были встречены на территории МНР и вообще в Азии или были известны лишь по очень разрозненным остаткам.
Изучение строения скелета найденных позвоночных позволяет сделать некоторые предположения об образе жизни и условиях существования животных. Помогало судить о мезозойских и кайнозойских ландшафтах Центральной Азии также изучение самих захоронений, условий их образования, что проводилось экспедицией на каждом местонахождении.
Добытые данные побудили И. А. Ефремова (1948, 1950) предложить следующую общую схему истории древней центрально-азиатской суши и жизни на ней в мезозое и кайнозое.
Континентальные мезозойские отложения Монголии начинаются с мощной толщи, вероятно нижней юры, частично размытой и подвергшейся выветриванию на глубине до 60 м. Характер этого выветривания, по мнению И. А. Ефремова, указывает на влажный тропический режим территории в раннемеловую эпоху. Большая мощность следующих выше слоев пижнемелового возраста, очевидно, говорит о длительном опускании страны в эпоху нижнего мела. В этих отложениях найдены остатки костистых рыб, насекомых, низших ракообразных, двустворчатых пресноводных моллюсков, а также крупные стволы деревьев.
Во второй половине мезозоя на территории Монголии располагались огромные заболоченные низменности с окружавшей их обильной растительностью. Обстановка в условиях влажного и теплого климата, вероятно, была благоприятна для многочисленных пресмыкающихся, в том числе динозавров.
Самые древние мезозойские четвероногие позвоночные, обнаруженные на территории МНР, - нижнемеловые "птицетазовые динозавры" из орнитопод (птиценогие) - игуанодоны и пситтакозавры (рис. 28-31). Напомним, что в настоящее время общепринято считать "ящеротазовых динозавров" и "птицетазовых динозавров" двумя разными отрядами пресмыкающихся. Сам термин динозавры (с греческого-страшные, тяжеловесные ящеры), предложенный в прошлом веке Ричардом Оуэном на основе первых находок гигантских ящеров, оказался неудачен, так как позднее были открыты небольшие и мелкие динозавры, в том числе прыгавшие и величиной с тушканчика. Тем не менее термин динозавры, укоренившийся за многие десятилетия в палеонтологии, применяется до сих пор.
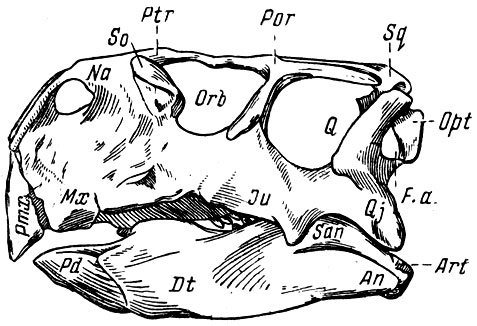
Рис. 28. Пситтакозавр (Psittacosaurus mongoliensis Osborn)
Череп пситтакозавра (рис. 28, 29) относительно размеров тела животного крупный, с высокой лицевой частью. Как обычно, у птицетазовых динозавров конец морды лишен зубов, но вооружен сильным роговым клювом, который при жизни животного еще более увеличивал его сходство с попугаем. Довольно большая височная впадина говорит о сильной височной мускулатуре, и хотя пситтакозавр - типичный представитель (птицетазовых растительноядных динозавров, вероятно, он мог своим мощным клювом не только добывать растительную пищу, но и наносить очень сильные укусы, а быть может, не пренебрегал и; мелкой живностью.
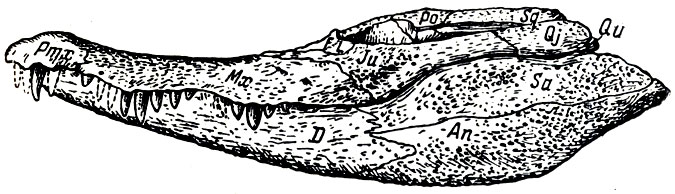
Рис. 29. Параллигатор (Paralligator gradilifrons Konjukova). Череп, вид сбоку, приблизительно 1/2 натуральной величины (Е. Д. Конжукова, 1954)
Окостеневшие сухожилия, сохранившиеся в крестцовой области позвоночного столба, несомненно гибкие и упругие при жиеши животного, в общем, как и строение таза, те же, что и у более поздних утконосых динозавров, но лапы пситтакозавра архаичнее (на задних сохранились все пять пальцев и т. д.).
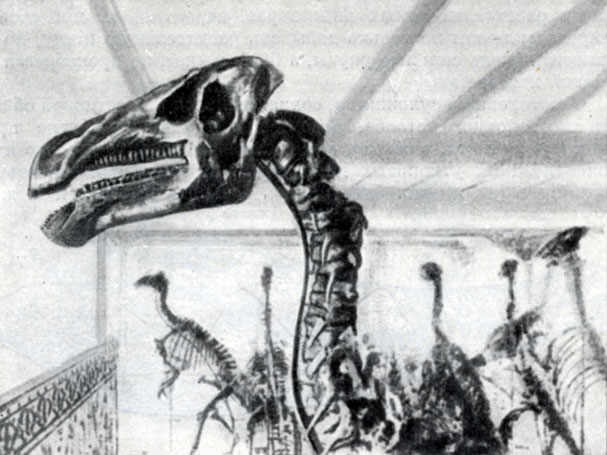
Рис. 30. Игуанодон (Iguanodon bernissartensis Boulenger) длиной около 10 м. Нижний мел Берниссара. Бельгия, Брюссель, Естественно-исторический музеей
Эти животные сочетают в себе признаки трех подотрядов птицетазовых динозавров - орнитопод, панцирных динозавров и рогатых; поэтому пситтакозавров нередко относили то к одному, то к другому отряду. Это типичные орнитоподы ("птиценогие"), передвигавшиеся на двух ногах; череп в общем того же типа, что и у примитивных рогатых динозавров, а зубы сходны с зубами панцирных. Такая смесь признаков объясняется, вероятно, тем, что три основные группы птицетазовых - орнитоподы, рогатые и панцирные - произошли от общих предков. Сами пситтакозавры вряд ли могут быть предками одной из трех групп. (По-видимому, они отделились от общего "ствола" птицетазовых динозавров в начало нижнего мела, когда началось становление и быстрое развитие основных ветвей этой группы.) Недавно остатки пситтакозавров обнаружены в Западной Сибири; они известны также из Северного Китая. Все описанные (виды пситтакозавров относятся к одному роду - пситтакозавр, названному так за сходство в строении лицевой части черепа с (попугаем (Psittacus). Не исключено, что Центральная Азия была вообще центром возникновения птицетазовых динозавров.

Рис. 31. Реконструкция внешнего вида игуанодона (W. Е. Swinton, 1958)
Название игуанодон указывает на сходство этих динозавров в строении зубов с крупными современными ящерицами игуанами. Челюсть европейских видов игуанодона несет 23 ряда зубов, а у монгольского вида их 27. Его лопатка той же величины и формы, что и у найденного в Бельгии. Вероятно, монгольский вид достигал размеров бельгийского (рис. 30, 31).
Китайскими палеонтологами описаны игуанодоны из северо-восточного Китая; этот род санпазавр, очень близкий к игуанодону, впервые обнаруженному в Западной Европе, а недавно и в Монгольской Народной Республике, где игуанодон представлен, по данным А. К. Рождественского, особым видом Iguanodon orientalis из Хамарин-Хурала.
Игуанодоны, как и пситтакозавры, были, по-видимому, приспособлены к обитанию во влажных, с богатой растительностью местах, которые со временем, возможно, стали убежищем для многих растительноядных динозавров, лишенных сильных защитных средств. Полукопытного характера конечные фаланги задних и передних конечностей у игуанодонов и пситтако-завров, вероятно, говорят о приспособлении этих животных к хождению по топким грунтам. Это отчасти подтверждается и отпечатками задних лап игуанодонов на мягком грунте. Отсутствие особых защитных средств у этих динозавров, кроме, предположительно, первого пальца передней конечности с его напоминающей кинжал длинной конической острой когтевой фалангой, также побуждает предполагать, что они должны бы обитать в местах, где легко прятаться от хищных динозавров, типично сухопутных по своему строению. Такими местами могли быть заросли вокруг болот, на побережьях озер и других водоемов, где игуанодоны могм находить корм и убежище.
В верхнем мелу пситтакоэавролв и игуанодонов сменили утконосые динозавры, более приспособленные к обитанию в водной среде или к полуводному образу жизни. Слои с поитта-козаврами и игуанодонами характеризуют нижние горизонты континентальных отложений мелового периода в Монголии (их, вероятно, можно сопоставить с верхней частью свиты Тен-дагуру в Африке и с формацией Моррисон в Северной Америке; в Европе им, по-видимому, соответствует Велд).
По данным И. А. Ефремова, в конце раннемелового временя на территории МНР произошли медленные поднятия земной коры с разломами и подводными излияниями главным образом базальтовых лав. Осадки этой эпохи в южных и северных районах различны по своему характеру; разнородны и местонахождения динозавров и других пресмыкающихся. К этому же времени И. А. Ефремов относит "внутридельтовые" дюнные пески в местонахождении Баин-Дзак, недалеко от Далан-Дзадагада в Южной Гоби. Здесь были найдены небольшие растительноядные динозавры-протоцератопсы с зачаточным рогом на носу и яйца динозавров, скорее всего самих протоцератопсов (рис. 32, 33). Протоцератопсы - предки типичных цератопсов, более поздних крупных рогатых динозавров из верхнемеловых отложений Азии и главным образом Северной Америки, где их череп достигал двух метров длины.
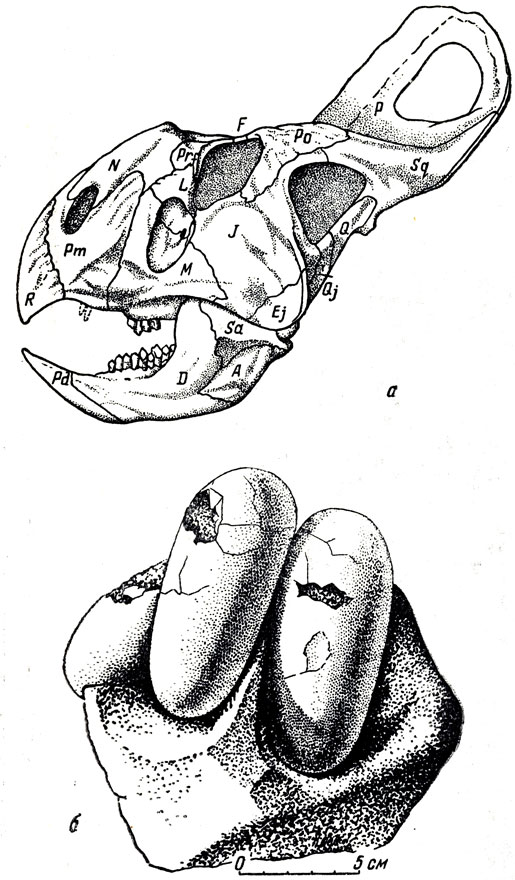
Рис. 32. Протоцератопс (Protoceratops andrewsi Brown et Schlajkier) a - череп; б - яйца динозавров (предположительно Protoceratops). Верхний мел, Баин-Дзак, Монголия. Музей Палеонтологического института АН СССР
Эволюция панцирных динозавров (анкилозавров), найденных в тех же местах, шла по пути слияния отдельных костных щитков, покрывавших в виде поясов тело животного сверху, и укрепления панциря. Особенно мощны такие бронированные полосы толщиной до 5 см в области крестца - наиболее высоко приподнятой части тела. Поверх пластин в коже развивались еще костные шипы паримидальной формы. На территории Монголии известны сравнительно ранние анкилозавры - оир-мозавры (Syrmosauridae), характеризующие приблизительно середину верхнего мела (соответствует формации Беллиривер Северной Америки, а по европейской шкале - "Каштану").

Рис. 33. Протоцератопс (Protoceratops andreWsi Brown et Schlajkier). Реконструкция внешнего вида (рис. Чарльза Найта). Естественно-исторический музей, Чикаго
Сирмойавр - своеобразный найденный нашей экспедицией новый птицетазовый динозавр (рис. 34), возможно, прямой предок поздних верхнемеловых анкилозавров. В отличий у сирмозавра (Е. А. Малеев, 1952) еще нет сплошного панциря, который развился позднее, в конце мела. Имеются лишь ряды изолированных друг от друга шипов (рис. 34), как у юрского сцелидозавра. Ряды расположены симметрично: по бокам шеи, спины и хвоста, а также по наружной стороне передних и задних оконечностей. Туловище сирмозавра - типичное для верхнемеловых анкилозавров - широкое, приземистое. Спинные позвонки длинные, суставные концы тел позвонков уплощенные, грудная клетка широкая - все это говорит о малой гибкости позвоночного столба. Шея короткая. У шейных позвонков задняя суставная поверхность тела не совсем параллельна передней и опущена ниже передней, это облегчало животному поднимать голову выше туловища. Крестец состоит лишь из трех (у более поздних - из многих) слитных позвонков. Передняя часть подвздошной кости очень длинная. Ноги короткие, передние конечности пятипалые, очень короткие сравнительно с задними трехпалыми. Мощный хвост с пучками окостеневших сухожилий (при жизни, очевидно, упругих, как у птиц в йогах) заканчивается толстой горизонтальной костной пластиной, вроде булавы. Сирмозавр напоминает этим некоторых ископаемых южноамериканских броненосцев. Вряд ли сирмозавр был проворен. Своим внешним видом он, вероятно, напоминал современных небольших тропических "жабовидных" ящериц-фринозом (рис. 35), покрытых шипами. Общая длина скелета сирмозавра - четыре с половиной метра.
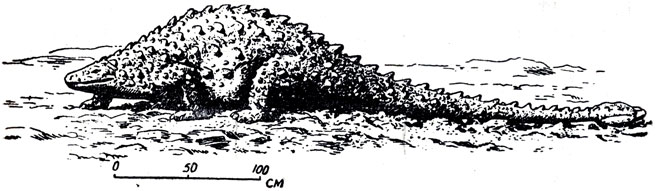
Рис. 34. Сирмозавр (byrmosaurus viminicaudatus Maleev). Реконструкция внешнего вида (рис. Н. А. Янышшова). Верхний мел, Баин-Дзак, Монголия
В сирмозавре обращает внимание примитивное, слабое развитие кожного защитного вооружения из отдельных изолированных шипов. Как указывалось выше, он напоминает этим более древних "панцирных" динозавров сцелидозавров (нижняя юра Англии). Черты архаичности сказываются и в малом числе крестцовых позвонков и в самостоятельности крестцовых ребер, не слитых с крестцовыми позвонками. (У типичных анкилозавров они слиты.) С другой стороны, "булава" хвоста из сросшихся 15-20 последних позвонков с концевой пластиной, строение плечевого пояса, сильная утолщенность таза около вертлужной впадины, вертикальная постановка подвздошных костей - все очень напоминает некоторых типичных анкилозавров.

Рис. 35. Жабовидная ящерица Северной Америки - фринозома (Phrinosoma cornutum). Длина около 15 см
Анкилозавры, скелеты которых были найдены в Восточной Гоби (Баин-Ширэ), отнесены Е. А. Малеевым к новому роду и виду - таларурус (рис. 36, 37) и, возможно, могут считаться более поздними, чем сирмозавр и протоцератопс. Таларурус ближе всего к типичному анкилозавру (род Ankylosaurus) и в средних и верхних горизонтов верхнего мела Соединенных Штатов или сколозавру (рис. 38). Череп таларуруса, судя по найденной задней части его, ниже и уже, чему у североамериканских анкилозавров; пластины, составляющие панцирь, ладьевидные, с "килем", сходны с пластинами североамериканского анкилозавра. По наружной поверхности щитов, по бокам хвоста и ног располагались симметрично полые толстостенные костные шипы. В отличие от других анкилозавров они сильно складчаты. Тяжелый панцирь таларуруса состоит из костных пластин, толщиной до пяти сантиметров, соединяющихся друг с другом при помощи слабо подвижных швов и образующих шейный, спинной и тазовый щиты; конечности массивные, с копытцами на концах пальцев; крестец длинный (четыре собственно крестцовых, четыре поясничных позвонка и один хвостовой); передние хвостовые позвонки короткие, (высокие и длинные; конечные хвостовые прочно, почти неподвижно, соединены в "ударное образование". Длина скелета 5-6 метров.
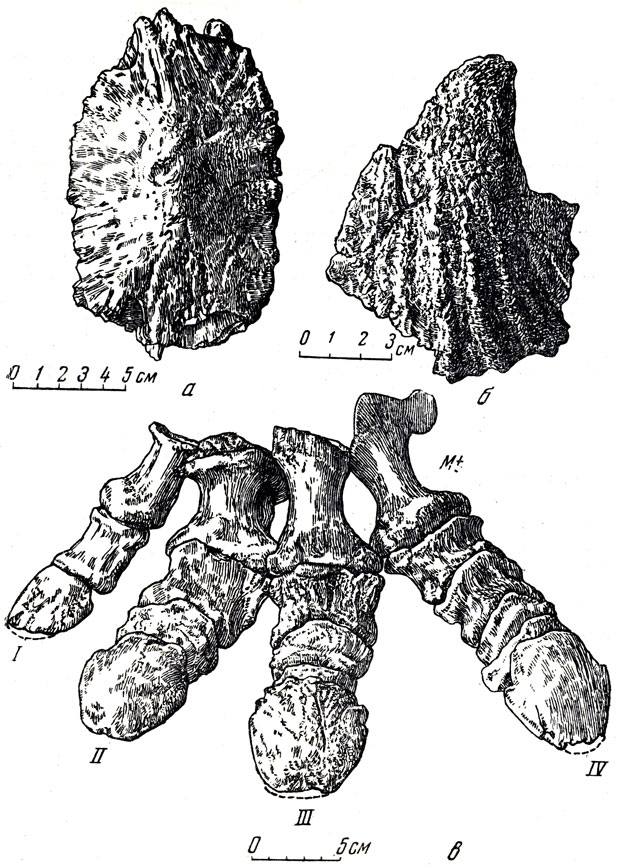
Рис. 36. Таларурус (Talarurus plicatospiaeus Maleev) а - пластина; б - шип панциря; в - стопа, вид сверху (5 фаланг в IV пальце, у рептилий он первично самый многофаланговый) Верхний мел, Баин-Шире, Монголия
В Монголии в это время еще жили и гигантские растительноядные динозавры типа диплодока или бронтозавра, но от них экспедицией собран сравнительно незначительный материал.
Для следующей, более молодой верхнемеловой фауны характерны гигантские утконосые динозавры - зауролофы и хищные; несколько скелетов тех и других было добыто в местонахождениях хребта Нэмэгэту в Южной Гоби. (Слои с костями этих динозавров, по-видимому, непосредственно продолжают слои Баин-Дзак и, возможно, могут быть сопоставлены с формацией эдмонтон Северной Америки, а в Европе - с Маастрихтом.)
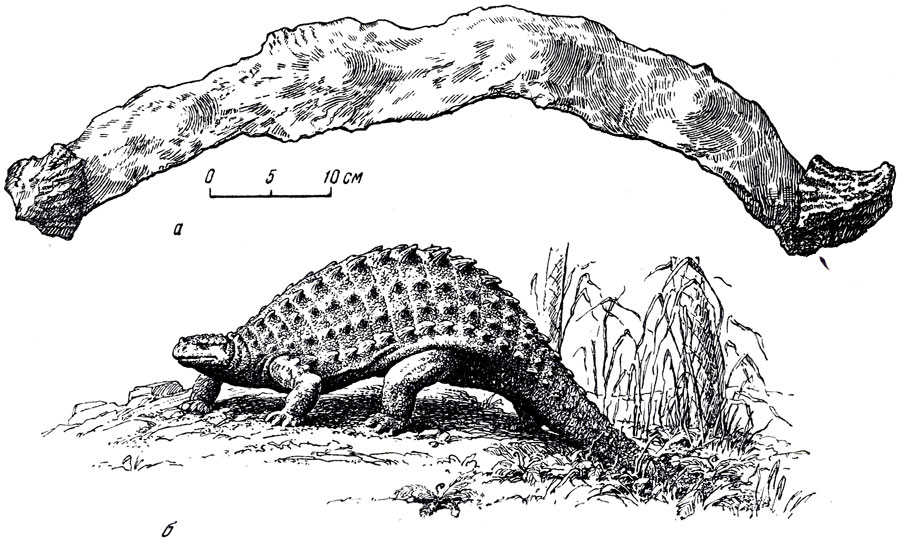
Рис. 37. Таларурус (Talarurus plicatospineus Maleev) a - часть спинного панциря сверху - дуга из нескольких прочно соединенных пластин с шипами на концах (см. рис. 36); б - реконструкция внешнего вида (рис. Н. А. Яньшинова). Верхний цел; Баин-Шире, Монголии
Большие захоронения остатков позвоночный начались, по данным И. А. Ефремова, с середины позднемеловой эпохи, что совпало, по его мнению, с уменьшением площади овер и поднятиями страны. Они ускоряли течения рек, впадавших в эти равнинные озера, и тем самым усиливали "врезание" рек в толщу озерных песков и песчаников со скелетами динозавров, крокодилов, черепах и других животных. Таково найденное экспедицией местонахождение около хребта Нэмэгэту - одно из самых больших в Старом Свете. Костенрсные слои залегают здесь именно в отложениях мощных древних русел, прорезавших в верхнемеловое время песчано-глинистые отложения больших пресноводных озер (рис. 40-41).

Рис. 38. Сколозавр (Scolosaurua cutleri Nopcsa). Около 1/35 натуральной величины. Верхний мел Канады (по А. С. Вундварду)
В нижних слоях этих русел, в песках и песчаниках, встречаются полные скелеты крупных динозавров, растительноядных и хищных; в верхних горизонтах - преимущественно черепахи (водные), крокодилы, остатки мелких хищных динозавров и отдельные части крупных, а также небольшие скопления пресноводных моллюсков, остатки пресноводных рыб. По данным И. А. Ефремова, широкие русла древних рек образовали, таким образом, костеносные слои и "выносы" динозавров, сравнительно удаленные от места гибели последних. Поперечник этих огромных русел составляет от двух до пяти километров.

Рис. 39. Обнажения глинисто-песчанистой толщи с ископаемыми остатками динозавров, черепах, аллигаторов и т. д. Верхний мел, Нэмэгэту, Южная Гоби, Монголия фото И. А. Ефремова)
Динозавры в местонахождениях Нэмэгэту разнообразны. Здесь найдены остатки гигантских "ящеротазовых" растительноядных (завропод), огромных хищных (карнозавров), целые скелеты растительноядных утконосых (гадрозавров).
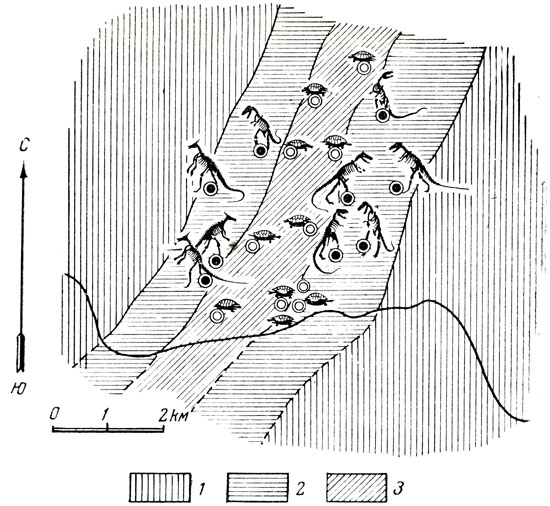
Рис. 40. Схематический план широкого (главного) косте-носного русла местонахождения Нэмэгэту, Южная Гоби, Монголия 1 - немые озерные песчаники и пески; 2 - пески и песчаники нижней зоны русла; 3 - конгломераты верхней и центральной зоны. Белые кружки (при черепахах) и черные (при динозаврах) даны для сопоставления со следующим рисунком
Утконосый динозавр зауролоф, по-видимому, был хорошо приспособлен к обитанию в прибрежной зоне бассейнов, что сказывается на строении черепа и конечностей (рис. 43, 45).
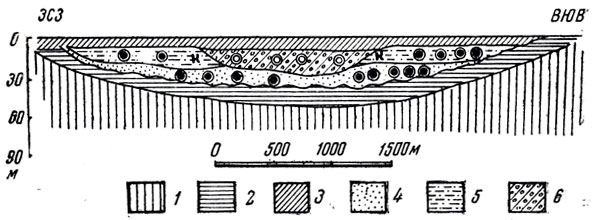
Рис. 41. Разрез главного костеносного русла местонахождения Нэмэгэту (по И. А. Ефремову, 1950. Рис. К. П. Мешкова). 1 - немые песчаники и пески озерной толщи; 2 - нижние красные-песчанистые глины; 3 - конгломераты; 4 - серые пески и песчаники с линзами 'шоколадных' известковистых глин; 5 - слоистые желтые пески; 6 - носослоистые гравийники и конгломераты; К - остатки крокодилов; остальные обозначения те же, что и на рис. 40
Склеротические кольца в орбитах, вероятно, говорят о частом пребывании головы под водой. Утиный по форме клюв был одет, как у гусей и уток, широким, сильным роговым чехлом и, как у них, вероятно, мог служить не только для срывания растительной пищи, но и для отцеживания воды. Зубная "батарея", состоявшая на каждой половине челюсти из 40 сменяющих друг друга и плотно соединенных рядов шечных зубов, по несколько сот в каждой челюсти, хорошо приспособлена для перерезания и измельчения растительной пищи, в том числе не только мягкой и сочной, но и жесткой. В связи с постоянным примешиванием минеральных частиц к пище, а, быть может, их содержанием в ней, и быстрым износом коронки развилась "функциональная гипсодонтность": каждый поперечный зубной ряд состоял из трех зубов, сидевших друг под другом (рис. 44).

Рис. 42. Остистые отростки динозавра зауролофа (Saurolophus angustirostris Rozhdestvensky). Алтан-Ула (фото Ю. А. Орлова, 1948)
Относительно слабый таз, вероятно, говорит о том, что зауролофы большую часть времени проводили в воде, а на суше вряд ли были очень подвижны. Передние лапы, судя по строению наружных пальцев, ревко отходящих в сторону, и фаланг, имеющих палочковидную форму, были снабжены плавательными перопонками, отпечатки и отливы которых хорошо сохранились у некоторых североамериканских верхнемеловых утконосых динозавров; зауролофы, вероятно, были приспособлены и к плаванию. Хвост, судя по скелету, высокий, также мог служить для плавания. Копытные фаланги в задней конечности широкие; если учесть огромный вес животного и нагрузку, которую, по-видимому, испытывала задняя лапа, это было важно для массивного животного при ходьбе, особенно по влажному субстрату, вероятно, и по дну водоемов (рис. 45).

Рис. 43. Зауролоф (Saurolophus angustirostris Rozhdestvensky). Скелет в Музее Палеонтологического института АН СССР. Верхний мел,, Нэмэгэту, Южная Гоби, Монголия
От североамериканского зауролофа монгольский отличается более узким черепом с длинным (загадочным) затылочным шпилем, направленным наэад более полого, иными пропорциями плеча и предплечья, более низким и широким тазом. Кроме нескольких скелетов зауролофа, из которых один монтирован (рис. 43), имеется серия черепов разного размера и возраста, отдельных костей и т. д. Утконосые динозавры - своего рода "копытные" среди пресмыкающихся по строению ног, а до известной степени и по характеру приспособленности к перетирающей работе щечных зубов. Но при всем бросающемся в глаза сходстве зубных "батарей" щечной области следует отметить и существенную разницу: коренные зубы копытных млекопитающих обладают неровной, хорошо перетирающей поверхностью, а у зауролофа она -гладкая, но с острым режущим внешним краем (рис. 44).
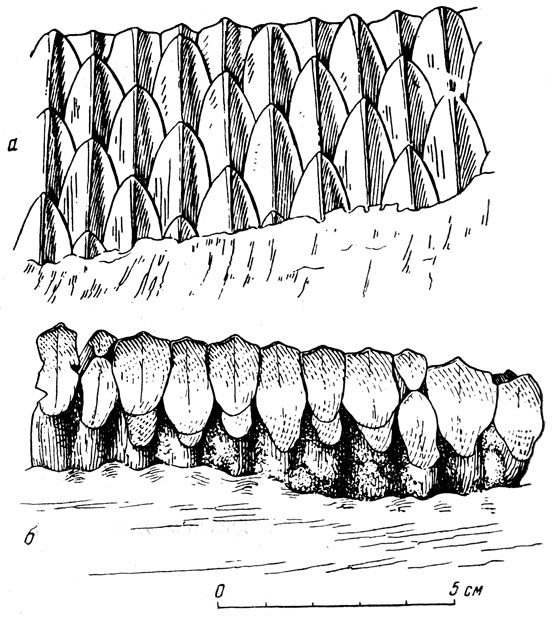
Рис. 44. Зауролоф (Saurolophus angustirostris Rozhdestvensky) Щечные зубы с язычной стороны (а) и жевательная поверхность, стертая косо наружу (б). Верхний мел, Нэмэгэту, Южная Гоби, Монголия
Западнее Нэмэгэту наша экспедиция обнаружила скопление нескольких скелетов зауролофов и отпечатки егожожи.
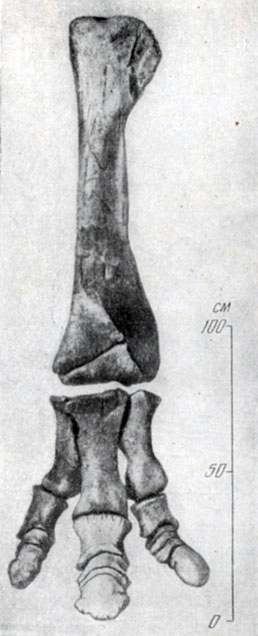
Рис. 45. Зауролоф (Saurolophus angustirtosris Rozhdestvensky). Левые большая берцовая кость и стопа. Верхний мел, Нэмэгэту, Южная Гоби, Монголия
Хищные динозавры, оставшиеся сухопутными, достигают в верхнем мелу Монголии очень крупных размеров. Это были, как известно, самые большие сухопутные хищники, когда-либо населявшие Землю. В Нэмэгэту найден тираннозавр, ранее известный род из верхнемеловых отложений Северной Америки; другой монгольский динозавр выделен в новый род - тарбозавр (рис. 46).

Рис. 46. Тарбозавр (Tarbosaurus bataar Maleev), очень близкий к тираннозавру (Tyrannosaurus), хищный динозавр. Музей Палеонтологического института АН СССР. Верхний мел, Нэмэгэту, Южная Гоби, Монголия
Коснемся некоторых особенностей строения динозавров и предполагаемою их образа жизни (по данным И. А. Ефремова и др.), и ирежде всего двуногого способа (хождения утконосых и всех хищных (он, как известно, первичен у всех древних динозавров; часть вернулась впоследствии к передвижению при помощи четырех ног). Причина двуыогости, возможно, лежит в приспособлении к жизни среди обильной и высокой растительности. Главный орган чувств динозавров, вообще пресмыкающихся (и птиц) - зрение (не обоняние, столь важное для млекопитающих). Высоко поднятая при двуногом передвижении голова была удобна для ориентировки среди высокой растительности и хищному динозавру, и прятавшемуся от него растительноядному.

Зауролофы в таксодиевом лесу. Рис. К. К. Флерова
Многие пресмыкающиеся, особенно мелкие, живут в сухой среде, но их непосредственные предки - земноводные - связаны с водой. Первичной обстановкой жизни пресмыкающихся, очевидно, были обширные прибрежные низины с густой растительностью.
Когда растения расселились на более высокие участки материков, расширились и первичные области обитания пресмыкающихся.
Однако выход из затененных растительностью областей гта открытые пространства, нередко под жаркое солнце, был для пресмыкающихся не прост. У рептилий нет надежной внешней термоизоляции, типа оперения птиц или волосяного покрова млекопитающих, нет и постоянной температуры крови, что свойственно птицам и млекопитающим. Все это означает для пресмыкающихся опасность перегрева в жаркое время. Вероятно, поэтому предки динозавров, приспособляясь к возвышенным и мало затененным частям материков, образовали ряд, по-видимому, ночных животных, а многие поселились в прибрежной зоне около рек и oisep (добавочная терморегуляция, быть может, могла происходить рефлекторно и при помощи теменного глаза). Млекопитающие, с их более высоким обменом веществ, не перешли (за очень редкими исключениями) к двуногому передвижению, которое оказалось выгодно для менее активного организма динозавра. Двуногие млекопитающие, как более активные, приспособлены, за исключением человека, к прыганию, тогда как пресмыкающиеся ходят, бегают или ползают.
Растительноядные, а за ними и хищные динозавры, вероятно, в основном обитали на влажных участках с густой растительностью. Разнообразие этого рода ландшафтов в мезооое содействовало и развитию разнообразных динозавров. По-видимому, в связи с этим обстоятельством (может быть, и с несовершенством терморегуляции) большинство крупных и гигантских растительноядных динозавров - лолуводные, прибрежные жители обширных бассейнов с богатой растительностью по берегам. Вне обильной растительности трудно представить существование гигантских растительноядных животных, хотя и нет отчетливых данных о том, какими именно растениями они питались. Разнообразие в строении зубов, видимо, говорит о различном растительном корме. Некоторые, вероятно, питались сочной водной растительностью, другие предпочитали листву и молодые побеги кустов и деревьев. Зубы некоторых уткопосых динозавров указывают на приспособленность к питанию и более жесткой растительной пищей. Небольшие зубы, по несколько сот в каждой челюсти, составляющие сплошные ряды, поразительно напоминают ряды коренных зубов копытных. Это и были "мезозойские копытные", так как их лапы оканчивались копытами,- но рептилии.
Хищные динозавры были наземными сухопутными животными. Характерно, что у водных растительноядных не обнаружено кожных защитных образований; они очень разнообразны у сухопутных растительноядных, нередко причудливы по форме - в виде костных рошв, шипов , вертикально стоящих на спине пластин или сплошного панциря, необходимых для защиты от огромных двуногих хищных, которым было удобно нападать на растительноядных сверху.
В местонахождении Нэмэгэту интересно преобладание именно огромных хищных динозавров, ранее неизвестных в Центральной Азии, в том числе гигантских, типа тираннозавра, вероятно, погибавших при наводнениях. Этот хищный ящер, возможно, был менее опасен для растительноядных вроде дип-людока или утжшосых, чем кажется на первый взгляд; для них, по-видимому, обычной средой обитания и спасения могла быть вода. У хищных динозавров задние ноги высокие, тазобедренные суставы сближены, что облегчало быстрый бег и насти-гание добычи. Расположение глазных владин в выступающих наружу углax черепа и направленность орбит вперед содействовали бинокулярному стереоскопическому зрению, столь важному для хищника. Длинные кинжаловидные зубы хищных диназавров сплюснуты с боков, задний и передний края зубов пильчатые; зубы сменялись на протяжении всей жизни. Пальцы высоких задних ног (и укороченных передних) были вооружены длинными когтями и совсем не похожи на копыта утконосых и других птицетазовых динозавров. По строению таза и скелета ног некоторые хищные динозавры имели вид как бы гигантских бегающих птиц с головой хищника (рис. 46). И след их лапы, как у гигантской птицы (рис. 47). Среди добытых нами хищных динозавров имеются и небольшие, типа орнитомимуса.
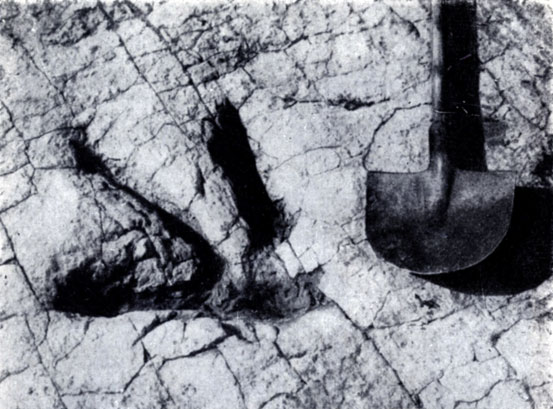
Рис. 47. Отпечаток задней лапы крупного хищного динозавра. Фото сотрудника Государственного музея МНР Намнандортка. Для масштаба - рядом лопата. Верхний мел Монголии
Очень многочисленны водные черепахи, найденные в этих отложениях; некоторые, возможно, указывают на недавнюю близость моря, так как по своему строению близки к морским; одна из них, судя по найденным остаткам, гигантская, очень близка к архелон верхнего мела Северной Америки, длина которой достигала 4-5 м (рис. 48), а длина черепа - 1 м. О близости моря говорят, быть может, и найденные здесь двустворчатые моллюски - новые виды "морского" облика, но, очевидно, приспособившиеся к жизни в пресной воде.
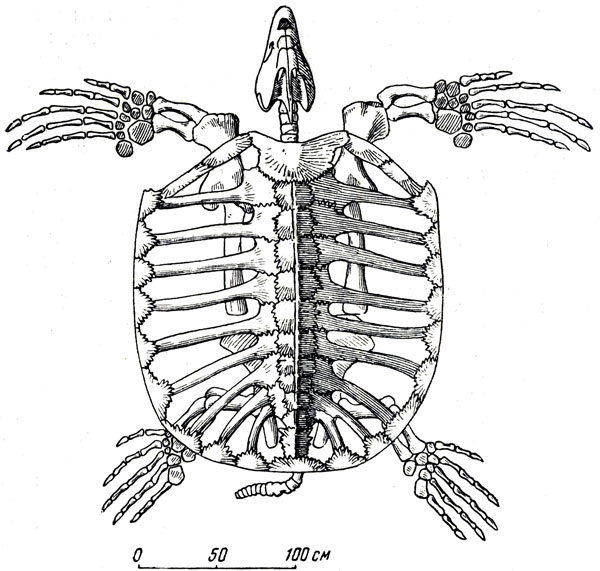
Рис. 48. Архелон (Archelon ishyros Wiel.), гигантская морская черепаха из верхнего мела США
Крокодилы, костистые рыбы, двустворчатые и, брюхоногие моллюски и низшие ракообразные дополняют пока имеющийся список верхнемеловой фауны обширной котловины Нэмэгэту. Аллигаторообразные крокодилы, входившие в состав этой фауны, выделены Е. Д. Конжуковой (1954) в особое семейство параллигаторов, близкое к наиболее широко распространенному в мезозое Западной Европы, Северной и Южной Америки семейству гониофолисов (из вымершего в меловое время подотряда мезозухий). Быть может, по мнению Конжуковой, параллигаторы (рис. 29) - прямые предки современных аллигаторов, широко распространенных в Азии и Северной Америке.

Тираннозавр. Рис К. К. Флерова
Ракообразные - эстерии (рис. 49) собраны в отложениях береговой полосы озер вместе с остатками растений. Хвойные деревья, типа болотного кипариса (таксодиума), современных субтропиков Северной Америки представлены окаменевшими стволами, нередко огромными, с прекрасно видной годичной слоистостью древесины. Хорошо сохранились длинные стволы магнолиевых.
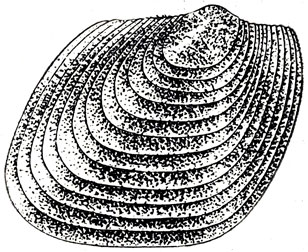
Рис. 49. Ромбограпта (Rhombograpta rhombica Novojilov), раковинчатое пресноводное ракообразное Верхний мел, Нэмэгэту, Южная Гоби, Монголия. Увеличено
Отложения с фауной конца мелового периода экспедиция обнаружила в Восточной Гоби. Характер осадков говорит о том, что при поднятии страны наступил сильный размыв континентальных отложений. Затем, вероятно, установился более сухой климат. На это указывают "пустынные многогранники" древнейших пород в костеносных слоях, обточенные песком и ветром, и найденные здесь панцирные динозавры - таларурусы, которые в отличие от бронтозавров и утконосых - гадрозавров, приспособлены к обитанию на более сухих местах. К этому периоду относятся красные озерные глины со скелетами крупных панцирных динозавров, вынесенных водой в эти озера в виде целых трупов.
Позднее по этому же ископаемому бассейну распространяются слои с многочисленными малоповрежденными костями разных динозавров; очевидно, место их обитания находилось недалеко от места гибели. Встречены кости с ясными следами зубов, что говорит о гибели животных на суше, где их трупы объедали хищные динозавры. И здесь погребено много водных черепах, как и вообще в Монголии в отложениях суши мелового периода.
Начавшиеся в конце верхнего мела поднятия должны были привести к значительному осушению Центральной Азии (рис. 50), обеднению и во всяком случае сильному изменению растительного покрова, лишению разнообразных групп меловых пресмыкающихся их естественных зон обитания. Это все могло быть одной из причин вымирания динозавров, а частично и других групп.
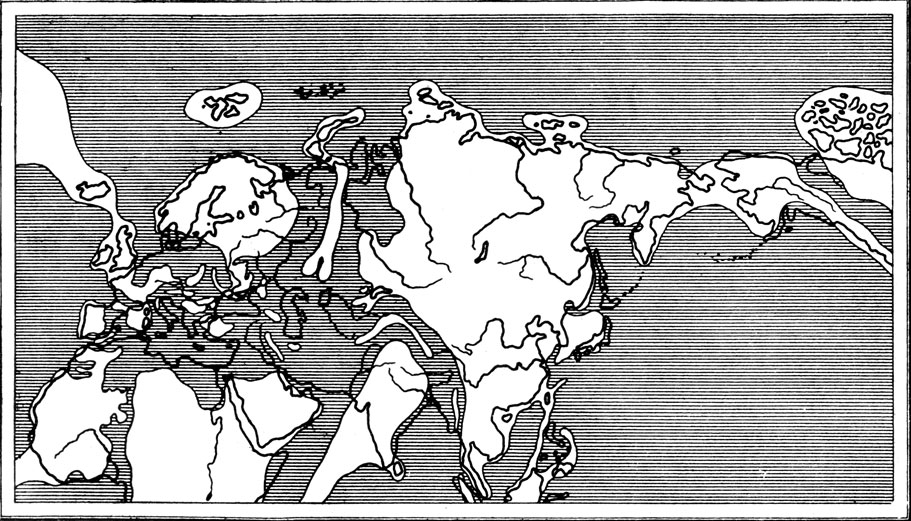
Рис. 50. Схематическая карта палеогеографии верхнемелового времени (по Термье, 1952)
Экспедиция не обнаружила в Монголии постепенного перехода отложений мелового периода к палеогеновым. Наши наблюдения говорят о континентальном режиме и о размыве верхнемеловых отложений с динозаврами в самом начале палеогенового периода в палеоцене, как и у нас в Средней Азии. Палеоценовые отложения, поздние, залегают с резким эрозионным несогласием на верхнем мелу.
Наиболее древняя фауна кайнозойских млекопитающих Монголии относится к верхнему палеоцену (местонахождение Гашато, близ Баин-Дзака в Южной Гоби). Отсюда известны архаические копытные, хищники и насекомоядные. Существование меловых млекопитающих, описанных из Баин-Дзака американскими палеонтологами, спорно. По-видимому, это палеоценовые млекопитающие, так как черепа добыты американскими палеонтологами из конкреций, залегавших на размытой поверхности верхнемеловых отложений и, вероятно, ошибочно приняты за меловые. Наблюдения Монгольской палеонтологической экспедиции показали, что конкреции с "меловыми млекопитающими" (при непосредственном прослеживании разреза) соответствуют слоям Гашато, содержащим типичных палеоцежхвых млекопитающих - продиноцерасов и др.
Млекопитающие, родственные фауне Гашато, были обнаружены в Нижнеэоценовом урочище Наран-Булак, около Нэмэгэту.
Нижнеэоценовые отложения в Гоби, по данным экспедиции, залегают в узких речных руслах, главным образом, широтного направления. Они врезаны в отложения мелового периода и идут, по данным Ефремова, поперек русел мелового (времени. К нижнему эоцену относится глинисто-песчанистая толща в Южной Гоби, где экспедиция добыла остатки небольших новых пантодонтов - древнейших копытных, еще пятипалых, без типичных копыт, почти когтистых, возможно всеядных. Это археоламбда - древнейший пантодонт Азии (рис. 51).
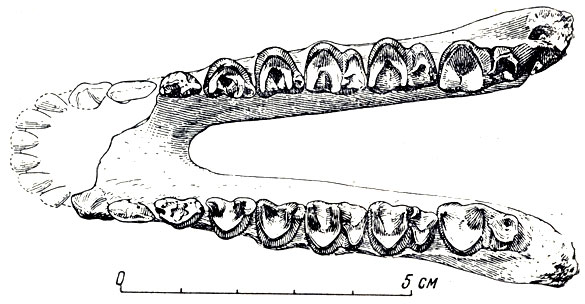
Рис. 51. Археоламбда (Archaeolambda planicanina Flerov), архаическое копытное из амблииод. Нижняя челюсть. Эоцен Наран-Булака, Монголия (К. К. Флеров, 1954)
По данным К. К. Флерова (1952), этот новый пантодонт должен быть выделен в особое семейство археоламбд с полной зубной системой, с зубами почти одинаковой высоты, непосредственно прилегающими друг к другу. Сходство зубов пантодонт с некоторыми рукокрылыми насекомоядного образа жизни указывает, по мнению Флерова, на невозможность отождествления лунчатых V-образных складок на зубах пантодонт ни морфологически, ни функционально с лунчатыми зубами растительноядных копытных; у пантодонт могли быть только давящережущие движения снизу вверх, и незначительо передне-задние (конвергентное сходство с копытными?).
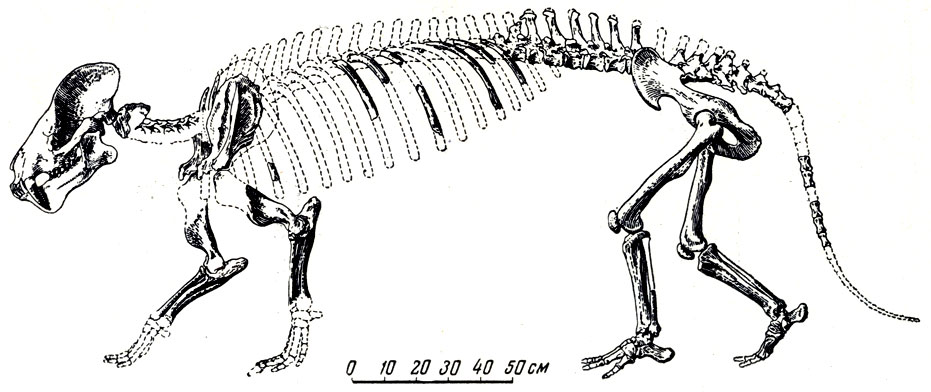
Рис. 52. Монголотерий (Mongolotherium plantigradum Flerov), архаический безрогий деноцерат. Эоцен Наран-Булака, Монголия (К. К. Флеров, 1954)
Археоламбда архаичнее североамериканской среднепалеоценовой пантоламбды, несмотря на свой несколько более поздний геологический возраст.
К. К. Флеров предполагает, что мы встречаемся с случаем, когда форма, продолжавшая существовать в месте своего первичного распространения в мало изменившихся условиях древ-неазиатского континента, сохранила много примитивных черт. В то же время американские пантодонты, ушедшие далеко в новые резко отличные ландшафты, претерпели значительные изменения. Возможно, что колыбелью пантодонт была часть палеазиатского материка, длительное время остававшаяся сушей.
Еще своеобразнее (найденные здесь же другие архаические "пятипалые копытные, новые диноцераты - монголотерии (рис. 5, 53).
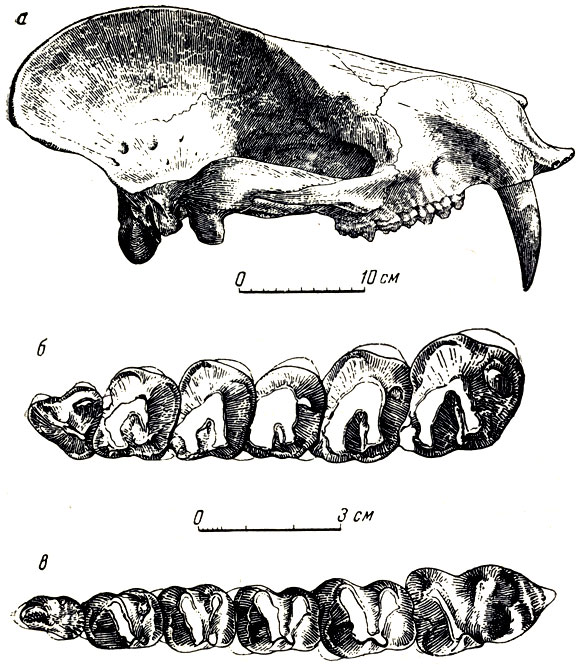
Рис. 53. Монголотерий (Mongolotherium plantigradum Flerov) а - череп; б - правые верхние и в - левые нижние лоданокоренные и коренные зубы. Эоцен Наран-Булака, Монголия (К. К. Флеров, 1954)
Большая коллекция черепов и костей монголотерия позволила установить следующее (К. К. Флеров, 1952): головной мозг монголотерия был незначительных размеров; череп без рогов, очень напоминающий череп первичных хищников-креодонтов с очень сильным сагиттальным и затылочным гребнями, вследствие чего верх затылка нависает над его нижней частью; скуловые дуги широкие; клыки саблеобразные, особенно большие у самцов, ложатся в "полуножны" нижней челюсти; челюсть несет в области массивного симфиза хорошо развитые выступающие вниз лопасти для предохранения клыков; туловище длинное, задние короткие конечности стопоходящие, таз длинный, узкий, выпрямленный; длинный мощный хвост составляет около трети длины всего позвоночника. Монголотерий - потомок палеоценового продиноцераса, но значительно прогрессивнее последнего. Американские и азиатские диноцераты уже с верхнего палеоцена развивались самостоятельно. Диноцераты, возможно, возникли в Азии, а потом расселились в Северную Америку, где, как известно, приняли вид странных огромных массивных животных, с туловищем и ногами слона, с тремя парами костных porols, с длинными копьевидными верхними клыками. Монголотерии безроги; другие азиатские диноцераты, гобиатерии, имели низкий череп и большой сводообразный вырост на носу. Гобиатерии^остигали величины носорога и, вероятно, вели полуводный образ жизни, подобно бегемотам, с чем связан, быть может, их причудливой формы очень низкий череп. В отличие от пантодонтов, это были, видимо, растительноядные животные, скорее всего питавшиеся листвой и молодыми побегами кустов и деревьев. Древнейшие копытные нижнего эоцена Монголии были, вероятно, по преимуществу обитателями лесов.
Здесь найдены также древние хищники, типа мезоникса Северной Америки, еще без настоящих "плотоядных" зубов, насекомоядный, черепахи, крупные рыбы типа "панцирных щук" лепидостеусов, а из растений - окаменелая древесина хвойных.
Более молодая, позднеэоценовая фауна собрана в Восточной Гоби в местонахождении Эргиль-Обо. В нее входят крупные, величиной почти со слона, ископаемые непарнопалые - броято-терии (от греческого бронтэ - гром), известные также под названием титанотериев и впервые открытые в середине прошлого века в Северной Америке. Они представлены в раннеолигоценовой фауне Эргиль-Обо крупным "протэмболотерием Ефремова" (рис. 54-56) с черепом около 75 см длины, для которого характерны направленные вверх массивные носовые кости с образованием огромного "тарана", суженного в средней части и расширенного у основания и на конце (греческое эмболэ - клин, таран). Боковые части носовых костей завернуты внутрь, к средней линии черепа.
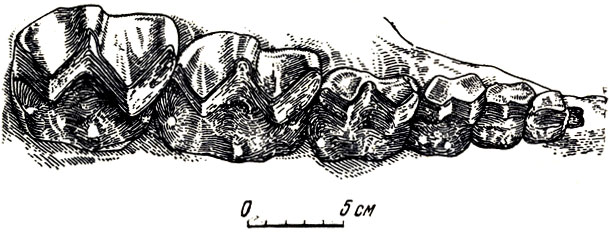
Рис. 54. Протемболотерий (Protembolotherium efremovi Janovskaja). Правый верхний зубной ряд Нижний олигоцен Эргиль-Обо, Монголия (Н. М. Яновская, 1954)
Низкие, но крупные щечные зубы бронтотериев могли быть приспособлены только для мягкой растительной пищи, низкие относительно слабые резцы и сходные с ними клыки могли быть пригодны для срывания листвы или мягкой растительности. Пищей эмболотериев, вероятно, была сочная прибрежная растительность рек, озер, топей, болот. Круто поднятые вверх носовые кости с их завернутыми внутрь внешними краями могли играть роль не только "оружия", но и служить костной опорой для удлинения носа с высоко поднятыми ноздрями; это могло быть приспособлением к свободному дыханию, когда морда "полупогружена" в воду при добывании из нее растений (рис. 55, 56). Из местонахождения Эргиль-Обо известны также мелкие примитивные носороги - ценолофусы.

Рис. 55. Протемболотерий (Protembolotherium efremovi Janovskaja) Реконструкция головы (рис. К. К. Флерова)
Следующая по времени раннеолигоценовая фауна обнаружена точно также в Эргиль-Обо и собрана из осадков олромной реки того времени, прорезавшей глинисто-песчанистые отложения более древних эоценовых озер. Характер осадков говорит, по мнению И. А. Ефремова, о том, что они отложились очень задолго до впадения реки в озеро. Эта фауна представлена в основном админодонтами, включающими несколько родов и видов: кадуркодон, гигантаминодон, гипсаминодон, аминодон. Аминодонты (рис. 57, 58) - крупные, с носорога ростом, массивные, неповоротливые непарнопалые очень своеобразного строения. Несмотря на свою непарнопалость в смысле систематического положения и явное, хотя и отдаленное, родство с носорогами, аминодонты были практически почти "парнопалы" по-развитию и способу хождения пальцев; в передних ногах давление распределялось почти равномерно на четыре пальца (II-V). Еще курьезнее у аминодонтов, по данным В. И. Громовой (1954), то, что опора была не на последние фаланги пальцев, (у носорогов копытного характера и сильные), а на расширенные нижние концы пястных и плюсневых костей (метакарпальных и метатарзальных), на крупные сесамовидные кости и на нижние поверхности первых и вторых фаланг. Была явно утеряна опорная функция конечных третьих фаланг (которым надлежало бы на первый взгляд нести копыта). Верхние и нижние клыки аминодонтов не перекрещены, как у большинства млекопитающих, а противостоят друг другу (рис. 57), сильно снашиваются и явно представляют приспособление для добывания корма, но не специальное "оружие боя". Взаимное расположение клыков, характер стирания коронок, редукция передних и массивность задних коренных зибов, ряд особенностей строении черепа, шеи и других частей скелета приводят В. И. Громиму к предположению о полуводном, "болотном" образе жизни этих странных родичей носорогов. Вероятно, аминодонты были обитателями низменных равнин по берегам больших рек, затоплявших эти равнины, где могли клыками захватывать и извлекать мясистые корневища и клубни водолюбивых растений, камышей, осок, аронниковых и др. (ом. рис. 58).

Энтелодопы. Рис. К. К. Флерова
Далее в состав этой раннеолигоценовой фауны входили примитивные таиирообразные эргилии, гигантские свинообразные - энтелодоны; своеобразные непарнопалые, пятипалые халикоте-рии схизотерии с когтями вместо копыт, древние хищники - креодонты (гиенодоны) и мелкие грызуны. Здесь же найдены еще не описанные остатки мелких и больших птиц, сухопутные черепахи, в том числе и крупные.

Рис. 56. Протемболотерий (Protembolotherium efremovi Janovskaja). Реконструкция внешнего вида (рис. К. К. Флерова по эскизу Н. М. Яровской, 1954)
Представители оюзднеэоценовой и раннеолигоценовой фауны Эргиль-Обо - бронтотерии, аминодонты, талирообразные и др. - были в основном жителями влажных, частью заболоченных лесов. Многие из них предки или предшественники среднеолигоценовых млекопитающих, широко известных на территории как Монголии, так и Казахстана.
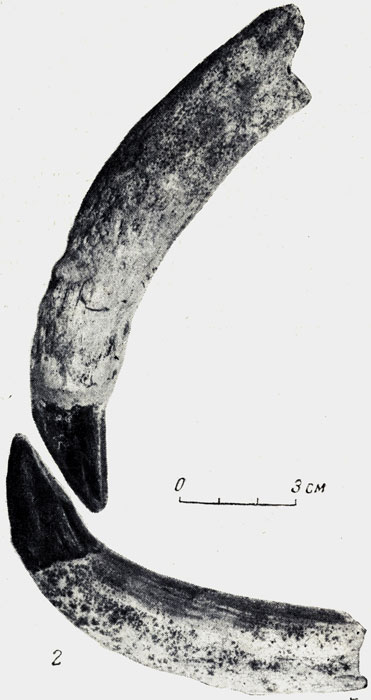
Рис. 57. Кадуркодон (Cadurcodon ardynensis Osborn), аминодонт 1 и 2 - верхний и нижний клыки самца. Уменьшено. Нижний олигоцен Эргиль-Обо, Монголия. (В. И. Громова, 1954)
Следующая по времени средноолигоценовая фауна, собранная экспедицией в Юго-Западной Гоби в местонахождении Тагал-Гол, носит почти такой же характер, как и одновозрастная казахстанская палеогеновая "индрикотериевая"( впервые установленная для Азии и описанная в ряде работ А. А. Борисяком (1916-1943).

Рис. 58. Кадуркодон (Gadurcodon ardynensis Osborn) Реконструкция К. К. Флерова (В. И. Громова, 1954)
Из непарнопалых в нее входят гигантские, почти жирафообразные безрогие носороги типа казахстанского индрикотерия, названного так по имени "Индрика - Зверя" древнерусских оказаний, и мелкие носороги - аллацеропсы. Из других непарнопалых - близкие к тапирам, а также халикотерии с большими когтями вместо копыт. Жвачные (миомерикосы, (рис. 59) в то время были еще мелкие, некоторые из них близки к предкам оленей и жираф, или же сами были их предками. Из насекомоядных найден близкий к ежам палеоскаптор. Особенно многочисленны ископаемые грызуны Монголии, размерами от суслика до бобра. Их около 20 видов: белкообразнЫе, дикообразоподобные, крупные и мелкие роющие - хомяки, слепыши и др.

Рис. 59. Мелкие жвачные миомериксы (Miomeryx). Средний олигоцен Азии . Рис. К. К. Флерова
Разнообразны хищники: мелкие, в том числе из куниц (общий род Азии и Америки), крупные креодонты до медведя величиной, часто древнего облика, гиенодоны.
Животный мир Казахстана и Монголии в то время был очень сходен, но в Казахстане найдено больше влаголюбивых животных (свиней, лофиомериксов, антракотериев).

Индриектории - Рис. К. К. Флерова
Видимо, в Монголии, как и в Казахстане, в то время на более сухих пространствах с разреженными лесами жили носороги, мелкие жвачные, многочисленные роющие грызуны и другие животные. На более влажных участках, где около больших водоемов росли широколиственные породы деревьев, а в заболоченных местах была влаголюбивая растительность, обитали жители болот и влажных лесов - таниры, аминодонты, некоторые парнокопытные, ежи, грызуны, напоминавшие белок, и др. В среднем олигоцене Монголии, по имеющимся материалам, преобладали все же млекопитающие - жители относительно сухих пространств.
По данным И. А. Ефремова, позднее, в верхнем олигоцене и нижнем миоцене, в Монголии озерные бассейны снова сокращаются, что связано отчасти с развитием горных хребтов. На западе Монголии при их появлении образуются предгорные наносные равнины с новыми озерами и реками, в которых постепенно накапливаются многочисленные остатки позднетретичных млекопитающих. В это время на территории Монголии живут хоботные (предположительно выходцы из Африки), уже проникшие в это время и в Северную Америку. Некоторые бегемотообразные носороги того времени, мастодонты и трехпалые лошади, представлены одними и теми же родами в Европе, Азии и Северной Америке. Нижнемиоценовая фауна известна из Улан-Тологоя, неподалеку от Татал-Гола Здесь встречены остатки древних мастодонтов - серридентинов, мелких трехпалых лошадей - анхитериев, полорогих жвачных - гобиоцерасов, более примитивных, чем в Европе и Африке. Возможно, что вся фауна формировалась именно в Центральной Азии. По образу жизни млекопитающие Улан-Тологоя были обитателями низин с сочной растительностью.
В конце миоцена и начале плиоцена, с развитием лесостепей и степей, в Азии широко расселилась фауна млекопитающих, для которых характерна трехпалая лошадь - гиппарион. Этзт фауну, кроме гиппариона, составляли хоботные, носороги, разнообразные жирафы, олени, антилопы, газели, кошки (мелкие и величиной с тигра), гиены, страусы и другие животные, теперь в общем близкие к типичным для саванн тропической Африки (см. гл. III).
В то время Сибирь и Китай еще не были разделены горами Алтая, Тянь-Шаня и других крупных хребтов. Огромное местонахождение гиппарионовой фауны, найденное экспедицией $ Западной Монголии, содержит, по-видимому, остатки тех же трехпалых лошадей, носорогов, жираф, крупных антилоп и газелей, которые бесчисленными стадами паслись на просторах нынешней Западной Сибири и Казахстана; Большое местонахождение этой фауны Алтан-Тээли в Западной Монголии является, возможно, результатом массовой гибели животных, вызванной эпизоотией или какой-то катастрофой, вроде наводнения. Здесь в хаотическом беспорядке нагромождены части скелетов, черепа и т. д. Эта фауна с жирафами, гиенами, страусами и другими теплолюбивыми животными рисует нам в общем ландшафт типа современных африканских лесостепей и саванн (см. гл. III).
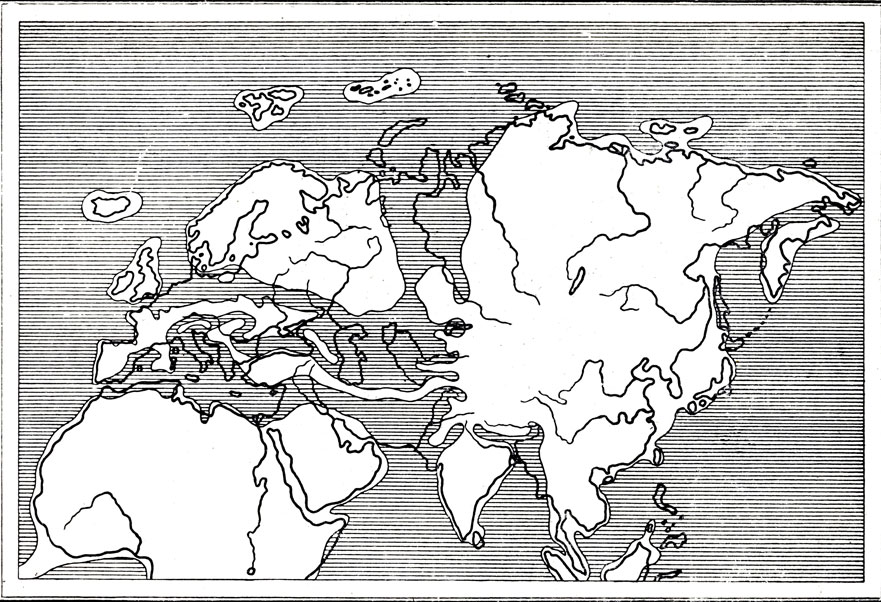
рис. 60. Схематическая палеографическая карта середины олигоцена (по Термье, 1952)
В настоящее время шет возможности проследить подробно историю развития всех упомянутых групп животных, так как мы пока не располагаем достаточно полными данными; более того, отдельные группы пресмыкающихся и млекопитающих, несомненно обитавшие на территории нынешней Монголии, пока еще здесь не открыты, таковы, например, пермские рептилии - несмотря на наличие пермских континентальных осадков; в мезозойских отложениях не встречены стегозавры, поздние цератопсы; не найдены многие архаические палеоценовые млекопитающие, примитивные среднеэоценовые носороги и креодонты и т. д. Все же можно судить" об истории позвоночных Центральной Азии, древней суши, где, по-видимому, на протяжении но крайней, мере полутора эр, а может быть и более, имелись благоприятные условия для возникновения и развития отдельных групп и для формирования целых фаунистических сообществ с последующим их расселением на другие части материка. Исследования экспедиции и изучение собранных материалов говорят о том, что климат в Монголии развивался в общем от влажного субтропического в сторону менее жаркого и более сухого, но еще теплого в конце неогена.
Таковы в самых общих чертах некоторые моменты истории жизни на Центрально-Азиатской суше в мезозое и кайнозое, как это представляется по, данным экспедиции Академии наук СССР (в отличие от некоторых более ранних данных американских исследователей).
Материалы экспедиции частично уже доступны для осмотра в Музее Палеонтологического института Академии наук СССР. Они послужат основой: для выставки, посвященной истории наземной жизни, Можно надеяться, что обработка материалов экспедиции поможет осветить историю многих групп животных и фаун в мезозое и кайнозое, и что итоги работы представят-интерес, выходящий за пределы изученной территории.
Со времени этой экспедиции прошло более десяти лет. Они внесли много нового в палеонтологию позвоночных Центральной Азии, прежде всего, работами Института палеонтологии позвоночных Китайской Народной Республики. Эти исследования, успешно развиваемые палеонтологами Китая, в частности на территории Автономной области Внутренняя Монголия КНР, куда продолжаются костеносные мезозойские и кайнозойские отложения МНР, не смогли быть включены в настоящую главу по своему объему, а также вследствие неопубликованности многих важных данных.
Нет сомнений в том, что эти работы внесут вскоре новый крупнейший вклад в палеонтологию Китая, всей Азии и в мировую науку.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'