
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Глава седьмая. Об изучении головного мозга ископаемых позвоночных
...Душа имеет свое местоположение преимущественно в маленькой железе, расположенной в центре мозга.
Р. Декарт ("О страстях души", 1649)
...Недавно еще существовал обычай взвешивать головной мозг людей, прославившихся при жизни умственной деятельностью, причем исходили из мысли, что весом мозга выражается... степень функционального развития гемисфер. Относительно попыток этого рода один исследователь не без основания заметил, что они имеют такое же значение, как если бы кто вздумал взвешивать целые города и судить по весу об их богатстве и благоустройстве.
И. Сеченов ("Физиологические очерки", ч. II, СПб.. 1898).
Предыдущие главы были посвящены нескольким фаунам ископаемых позвоночных СССР и Монголии разного геологического возраста.
В этой главе автор касается некоторых вопросов так называемой "палеоневрологии", т. е. изучения нервной системы ископаемых, их головного мозга.
Головной мозг не сохраняется в ископаемом состоянии; как и другие отделы нервной системы, он быстро подвергается разрушению после смерти животного. Правда, время от времени в литературе появлялись сообщения о находках "окаменелого мозга", в том числе и человека. В последнем случае они, естественно, вызывали сенсацию. Но пртг более подробной проверке было твердо установлено, что все, что когда-либо принималось за "окаменевший головной мозг", оказывалось чем угодно, но только не мозгом. Никакой минерализации мозга никогда не наблюдалось и не могло быть вследствие его состава и консистенции. Тем не менее изучение объема, внешней формы и характера рельефа больших полушарий, мозжечка и других отделов головного мозга, размеров отходящих от него нервов и т. п. на отливах (слепках) черепномозговой полости часто представляет значительный интерес. Для приготовления искусственных отливов, по размерам и форме заменяющих головной мозг, применяются воск, пластелип, гипс, каучук и т. д. Такой отлив довольно точно воспроизводит размеры и форму основных отделов головного мозга, а также рельеф его извилин и борозд, хотя и не во всех случаях отлив вполне соответствует по величине и характеру рельефа самому мозгу. Так, у рыб, земноводных и многих пресмыкающихся головной мозг часто отделен от костей крыши черепа значительными разращениями мозговых оболочек и сосудистых сплетений, а также и большими полостями для кровеносных резервуаров (кровеносными синусами). Все они иногда занимают значительное пространство, и тогда отлив полости мозговой коробки отличается размерами и пропорциями от головного мозга и крупное его. Наибольшее соответствие отлива самому мозгу наблюдается у птиц и особенно у млекопитающих, хотя и не у всех в одинаковой степени. У китообразных и многих крупных копытных мозг плотно лежит на основании черепа, но имеется некоторое пространство между мозгом и сводом черепа. При этом рельеф верхней стороны мозга не отражен в подробностях па внутренней поверхности свода черепа; отлив почти лишен борозд и извилин (рис. 102) и вдобавок несколько больше самого мозга. Наиболее точное представление о размерах и пропорциях отделов, о рельефе поверхности мозга можно получить по отливам мозговой коробки у многих насекомоядных, рукокрылых, хищников и др. В особенности отчетливое представление можно получить о головном мозге при рано наступающем зарастании некоторых швов черепа, когда оно происходит до окончания роста голодного мозга, например у (некоторых хищников и др. Рост мозга, продолжающийся некоторое время после сращения костей мозговой коробки, и оказываемое растущим мозгом давление на внутреннюю поверхность костей черепа приводят к резорбции (рассасыванию) кости, в которую мозг как бы втискивается своими извилинами. Вероятно, это явление имеет и некоторое приспособительное значение, так как этим путем достигается предохранение мозга от сотрясений при скачках, прыжках и т. п., что важно для хищников и многих копытных. В случае такого раннего зарастания швов у костей крыши черепа, рельеф ее внутренней поверхности, обращенной к мозгу, будет точно отвечать поверхности самого мозга. Отлив мозговой полости станет отражать общую внешнюю анатомию головного мозга и в больших подробностях его извилины, борозды, крупные, а часто и мелкие сосуды на его поверхности и т. д. У некоторых животных зарастание швов крыши черепа наступает относительно рано, задолго до окончания роста головного мозга, и он довольно долго "втискивается" в кости черепа. Стенка черепа (лобные, теменные, височные кости и чешуя затылочной) истончается давлением изнутри, но в это же время внешняя надкостница продолжает рост кости в толщину наложением новых слоев. Если этот процесс "наступания" головного мозга на кость и одновременно идущего наслаивания кости извне продолжителен, тогда можно даже заметить отражение некоторых извилин больших полушарий головного мозга, "червячка" мозжечка и т. д, на внешней поверхности костей черепа (мозг собаки, куниц и др.) (Иногда в патологических случаях при водянке головного мозга).
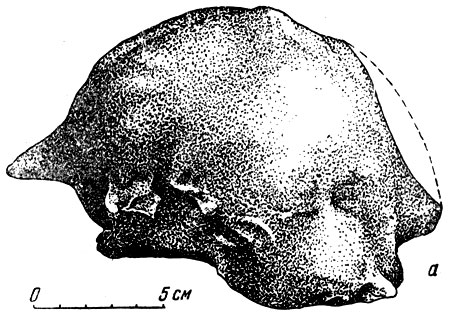
Рис. 102. Отливы головного мозга. а - петотерия (Cetotherium sp.)
Особенно точно воспроизводят форму и рельеф внешней поверхности головного мозга естественные отливы. Они получаются в черепах ископаемых животных в итоге заполнения мозговой коробки мелким песком, илистыми осадками и т. д. Тонкозернистые илистые пески дают очень точные отливы, подвергающиеся иногда окремнению, или прочной цементации солями кальция, окислами железа и т. д. (При недостаточной прочности естественный отлив можно пропитать каким-либо лаком.)
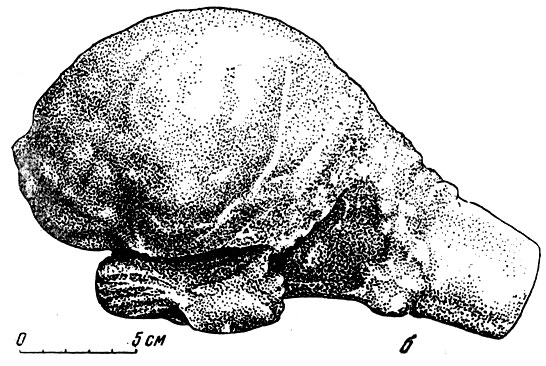
Рис. 102. Отливы головного мозга. б - эласмотерия (Elasmotherium sibiricum Fischer). Вид сбоку. Музей Палеонтологического института АН СССР, Уменьшено
Наиболее верные, "гальванопластически" точные отливы получаются в итоге заполнения мозговой коробки каким-либо осадком, выпавшим из раствора, например солями кальция и др. Иногда заполнение черепномозговой полости породой происходит до разрушения твердой мозговой оболочки и затем, спустя некоторое время, наступает замещение этой оболочки после ее разрушения каким-либо осадком, выпадающим из водного раствора. В таком случае, наряду с отливом самого головного мозга, наблюдается образование обособленного отлива плотно к нему прилегавшей и его одевавшей мозговой оболочки.
Кроме изготовления слепков можно пользоваться и помощью фотографии, особенно, если мы имеем дело с хрупкими ископаемыми костями, которые легко повредить изготовлением слепков. В таком случае с внутренней поверхности мозговой коробки, будет ли это искусственный распил черепа или неполный кусок мозговой коробки, найденный при раскопках, делается негатив; далее готовится прозрачный позитивный отпечаток, т. е. диапозитив, который применяется как негатив для приготовления отпечатка на фотографической бумаге. Этот "окончательный" отпечаток, естественно, будет иметь снова вид негатива: снимок, сделанный с внутренней, вогнутой стороны левой половины мозговой коробки, будет казаться выпуклым, принадлежащим правой стороне мозга, отпечатки сосудов и мозговых извилин на внутренней стороне костей черепа будут казаться выпуклым отливом сосудов и извилин правой стороны мозга; как на негативе, темное будет светлым, а светлое - темным.
Рассмотрим несколько отливов полости мозговой коробки, которые иногда для краткости будем называть головным мозгом. Слепок, избраженный на рис. 102, а, дает представление о головном: мозге цетотерия, ископаемого кита, остатки которого встречаются в отложениях обширного полузамкнутого верхнетретичного сарматского моря, простиравшегося от территории Румынии до района Аральского моря. На слепке в общем видны разные отделы мозга, например лобная и височная доли полушарий большого мозга. Местами (имеются слепки - сосудов, главным образом крупных венозных синусов, несущих кровь от мозга к внутренним яремным венам. Но сам рельеф поверхности большого мозга и мозжечка отражен на отливе очень слабо, почти незаметен; это затрудняет сравнение мозга цетотерия и его родичей современных китов. Отлив не дает возможности судить о развитии борозд и извилин; (поверхность представляется почти гладкой. Между тем для головного мозга современных китов характерно в общем именно сильное развитие извилин и борозд; при незначительных, ничтожных размерах головного мозга у крупных китов сравнительно с животным в целом сильная складчатость коры дает ей, как известно, возможность вме стить необходимое для высшей нервной деятельности число нервных элементов. Но отлив черепномозговой полости и у современного кита так же мало показателен, как и у цетотерия; об извилинах и бороздах головного мозга китов ископаемых и современных по отливу судить трудно.
Немногим лучше обстоит дело и для некоторых наиболее крупных копытных, например носорогов. На рис. 102, б изображен отлив черепномозговой полости эласмотерия, носорога, с небольшого слона ростом, жившего на территории СССР в четвертичном периоде. Этот слепок несколько выразительнее, чем у кита, но лишь в смысле общих пропорций больших полушарий, их долей и мозжечка, так как подробностей строения не видно. То же мы наблюдаем и на отливе полости мозговой коробки у слонов (рис. 103). Во всех этих и подобных случаях неясность относится лишь к верхней и отчасти к боковым поверхностям отлива головного мозга. Его нижняя сторона плотно прилегает к основанию черепа; поэтому все черты внешнего строения нижней стороны мозга, расположение отходящих от мозгя пар черепных нервов, их толщина, по которой можно догадываться о степени их развития - все это на отливах видно достаточно отчетливо. Примером может служить отлив черепномозговой полости индийского слона (рис. 105). Здесь хорошо видны округлый гипофиз, варолиев мост, тройничный и другие нервы. Обычно по отливу всегда можно получить отчетливое представление о развитии обонятельных луковиц н пучков волокон обонятельных нервов, а следовательно, до известной степени и о развитии обоняния у ископаемого млекопитающего.
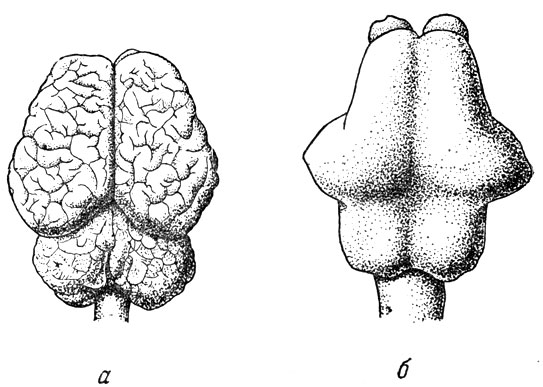
Рис. 103. Головной мозг индийского слона (Elephas iridicus) (а) и его отлив (б) (Т. Edinger, 1929)
Не у всех копытных, в том числе крупных, отлив мозга так мало выразителен с верхней стороны, как у эласмотерия или слона. В этом легко убедиться, если рассмотреть мозг гиппариона, ископаемой трехпалой лошади, населявшей неисчислимыми табунами территорию СССР, среднюю и южную Европу и Азию в верхнем миоцене и нижнем плиоцене. На отливе мозга гиппариона многие борозды и извилины достаточно отчетливы для сравнения с мозгом современной лошади. У нее также отлив довольно ясно передает рельеф полушарий, не говоря уже о границе большого мозга и мозжечка; их разделяет у лошади, как у хищников и некоторых других, поперечная относительно мозга костная пластина, тенториум ("костный намет"); он глубоко проникает между большим мозгом и мозжечком и поэтому на естественных отливах обычно отламывается от черепа и остается между большим мозгом и мозжечком.
Пожалуй, еще отчетливее картина у некоторых мелких копытных. Примером может служить небольшая газель, спутник гиппариона, остатки которой довольно обычны в местонахождениях этой фауны в Казахстане, на Украине и Кавказе. На отливе мозга (рис. 104) хорошо видны все основные извилины и борозды. Небезынтересно отметить, что уже в то время, в конце миоцена и нижнем плиоцене, т. е. несколько миллионов лет назад, головной мозг лошадей и антилоп по внешнему виду и относительным размерам достигал в сущности почти той же ступени развития, на которой мы видим его теперь, поскольку об этом для ископаемых животных мы в состоянии судить лишь по внешнему облику мозга.
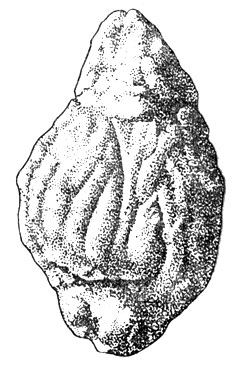
Рис. 104. Естественный отлив головного мозга ископаемой газели (Gazella deperdita Gaudry). Нижний плиоцен. Музей Палеонтологического института
Еще более четки отливы мозга многих хищников. Примером может служить перуниум, гигантская ископаемая куница, значительно (превосходившая размерами крупных росомах. Она входила в состав той же "фауны гиппариона", что и упомянутая газель (рис. 106). Для перуниума известен прекрасно сохранившийся естественный отлив полости черепа (внутреннее ядро). Мозговая коробка была вскрыта, порода (мелкий глинистый песок) пропитана для прочности лаком, после чего "головной мозг" был мною подвергнут сравнению с мозгом современных куниц (рис. 107). На отливе ясно видны все извилины и борозды, верхний продольный (сагиттальный) венозный синус и ветви поднимающейся по боковой стодоле полушария средней артерии мозговой оболочки.
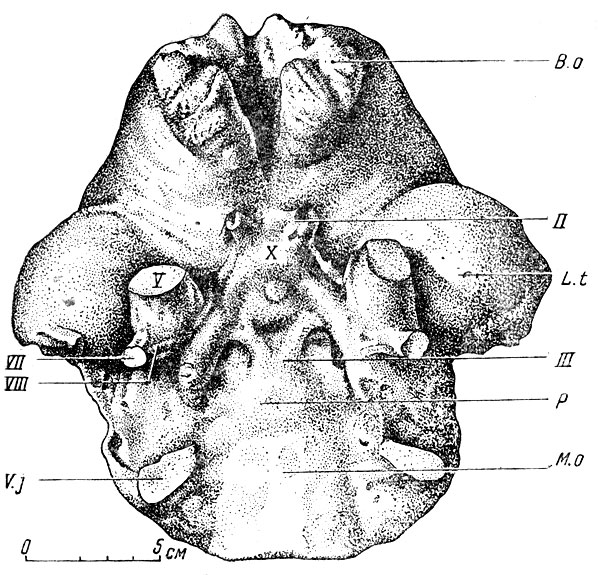
Рис. 105. Слон (Eiephas indicus). Отлив головного мозга, нижняя сторона II-VIII - черепные нервы; В. о.- обонятельные луковицы; L. t.- височная доля; Р - варолиев мост; M. о, -продолговатый мозг; V. j. - отлив яремного отверстия и его вены. Сильно уменьшено
Обращает на себя внимание очень сильное рдзвитие височном доли, как у медведей; она у перунмума делеко опускается вниз к основанию мозга и уже одним этим придает мозгу медведеобразный облик. Особенно велико сходство с мозгом белого медведя. Оно выражается для полушарий большого мозга в наличии трех борозд и четырех извилин в височно-затылочном отдела вместо двух борозд и трех извилин, встречающихся у куниц, в характере строения задневерхних углов полушарий (расчленения на извилины и борозды), в значительной высоте задней половины большого мозга, в сильном развитии затылочных извилин. От всех остальных куниц мозг перуниума отличается, кроме того, очень большим развитием сагиттальной (верхней) извилины полушарий большого мозга и некоторыми другими чертами. Но общий план строения мозга все же "куний"; и если расположить по возрастающей сложности извилин и борозд мозг различных членов семейства куниц, от мелких вроде ласки до самых крупных вроде росомахи, то конечным членом такого ряда будет мозг перуниума, медведеобразный не только по некоторым особенностям строения мозга, оно и черепа (рис. 108). Медведе-образность мозговой коробки черепа перуниума - в значительной степени итог сходных с медведем пропорций мозга. У млекопитающих с крупным головным мозгом, каковы все известные куницы, он, естественно, в сильной мере определяет форму мозговой коробки (и черепа в целом).
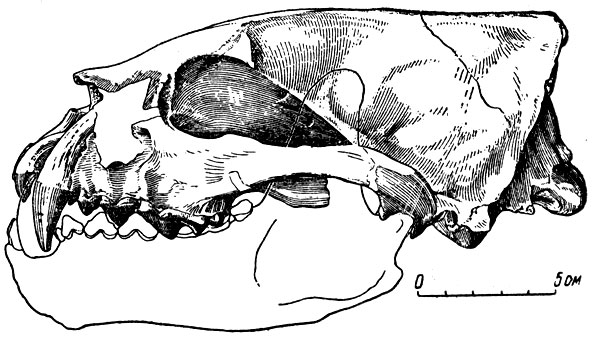
Рис. 106. Гигантская ископаемая куница перуниум (Perunium ursogulo Oriov). Череп (Ю. А. Орлов, 1948). Мэотис, село Гребеники Молдавской ССР
По строению головного мозга перуниум стоит особняком среди других куниц в еще большей степени, чем по строению черепа. Какие допустимы предположения о значении этих особенностей строения?
При невозможности знать внутреннее строение мозга перу ниума и при его слабой изученности даже у современных куниц а медведей (насколько известно, не представляющих собою обычного экспериментального материала физиологов) надлежит обратиться к другим млекопитающим и, в частности, к ближайшим родственникам куниц и медведей - собакам, головной мозг которых и локализация функций в его коре подверглись столь полному изучению в блестящих работах И. П. Павлова и его школы и многих других. Несмотря (на различия в строении извилин и борозд мозга, с одной стороны, куниц и медведей, с другой - собак, можно сказать следующее. Прежде всего, одна из особенностей большого мозга перуниума, сравнительно с другими куницами, - необычное развитие височной доли. Это должно было бы означать и какую-то особенность высшей нервной деятельности этой "психоакустической" области коры большого мозга; обязанность "области слуха" не только слышать звук, но к уметь дать ему должное толкование. У млекопитающих височная доля отличается замечательным разнообразием строения в смысле составляющих ее нервных элементов, что должно говорить о разнообразии специализации и дефференцировки слуха млекопитающих. Прогрессивное развитие височных долей и умение правильно толковать звук - одна из важнейших сторон связи животного с условиями его существования, со средой в самом широком смысле слова. Тем самым прогресс в трактовке звуков должен, вообще говоря, содействовать прогрессу лобных долей. Но у перуниума лобные доли развиты относительно слабо, без сколько-нибудь заметного прогресса в соответствии с развитием височных (видимо, то же наблюдается и у медведя).
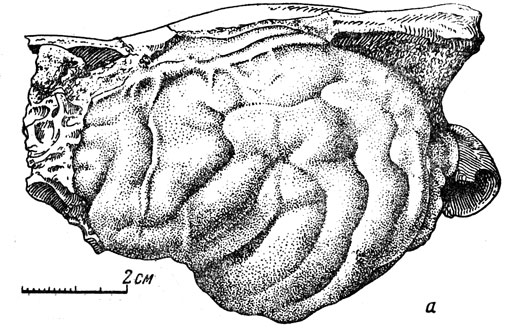
Рис. 107. Гигантская ископаемая куница перуниум (Perunium ursogulo Orlov). а - естественный отлив черепномозговой полости
Сильное развитие затылочной доли большого мозга, возможно, говорит о дифференцировке и какой-то специфике зрительной области мозга.
Для мозжечка перуниума при общем строении по типу ку ниц любопытно сильное развитие полушарий мозжечка. Это, вероятно, показатель ловкости и проворства животного в пользовании парными конечностями.
Пример перуниума указывает лишний раз на относительность положения о прогрессивном нарастании извилистости коры большого мозга с увеличением в ходе исторического развития размеров животного. Если бы это положение было непреложной истиной, то у перуниума, огромного сравнительно с барсуком или куницей, можно было бы ожидать значительно большего развития извилин и борозд, чем у них. Между тем у маленькой сравнительно с перупиумом куницы на боковой поверхности полушарий большого мозга имеются все основные борозды и извилины, что и у перулиума, а лобная доля у перуниума даже отстала в развитии сравнительно с барсуком.
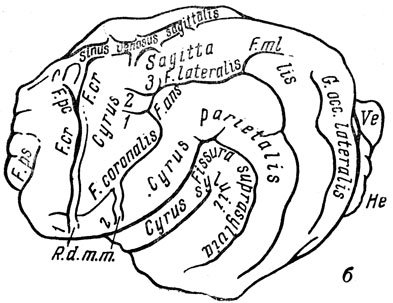
Рис. 107. Гигантская ископаемая куница перуниум (Perunium ursogulo Orlov). б - схема отлива. Уменьшено. Мэотис, село Гребешши Молдавской ССР
Категорические выводы о значении всех этих пропорций, или вернее "диспропорций" в развитии разных отделов мозга трудны и, вероятно, были бы легче при большой изученности мозга куниц и медведей. Но, быть может, мы имеем в перуииуме случай, когда увеличение теменной, затылочной (зрение) и особенно височной (слух) долей, другими словами, увеличение экрана или области проекции восприятия из внешнего мира с их обработкой в психосензорпых центрах коры мозга, не сопровождалось почему-то "соответственно" прогрессивным развитием лобных долей; это могло бы быть при сильном изменении внешних условий - оно налицо в среднем и верхнем плиоцене - одной из причин вымирания перуниума и его близких родственников.
В настоящее время, особенно при разработанности на основе учения И. П. Павлова представлений о головном мозге, его работе и о размещении функций на поверхности коры - изучение отливов мозга испокаемых млекопитающих представляет несомненный интерес. Но и изучение головного мозга низших ископаемых позвоночных, несмотря на меньшую точность по сравнению с млекопитающими отливов их черепномозговой полости в смысле соответствия самому головному мозгу, заслуживает внимания, так как позволяет проводить сопоставление с современными позвоночными и наметить некоторые черты эволюции мозга для разных групп позвоночных. К тому же головной мозг - единственный отдел "мягких частей", форма и объем которого доходят до нас из далекого прошлого с поразительной яркостью и с той же точностью, что и скелет, тогда как о внутренностях, сосудах, мускулатуре мы почти всегда вынуждены догадываться, хотя часто вполне обоснованно.

Рис. 108. Куница (Perunium ursogulo Orlov) Реконструкции внешнего вида (рис. К. К. Флерова)
Ограниченность данных, получаемых при изучении окаменевшего внутреннего ядра (или искусственного отлива) черепно-мозговой полости, сравнительно с итогами изучения самого мозга очевидна. Но палеонтологическому материалу свойственна особая, историческая ценность документов далекого прошлого, и одно это побуждает относиться к внутренним ядрам мозговой коробки позвоночных с должным вниманием.
Это несомненно сознавал еще Ж. Кювье (1835), давший первое описание мозга ископаемого млекопитающего по естественному отливу мозговой коробки и в нескольких словах положивший начало "палеоневрологии". Позднее Марш (1886) на большом материале по североамериканским архаическим копытным диноцератам дал картину исторического развития мозга этой группы. Но по справедливому замечанию Т. Эдингер (1955), это была эпоха "антропоцентрических" представлений, и "история" головного мозга для Марша заключалась в сущности в постепенном увеличении объема мозга, особенно его больших полушарий.
В настоящее время - и при известной отраженности внутренней структуры головного мозга на его внешнем облике и при наличии огромных и разнообразных сравнительных материалов по мозгу современных животных, наконец, при накоплении больших материалов по мозгу (т. е. отливам) ископаемых животных - палеонтология мозга естественно предстает перед нами сравнительно с прошлым веком в ином виде. В особенности пополнились наши знания о головном мозге ископаемых млекопитающих, отлив черепномозговой полости которых, как известно, более точно соответствует но размерам и форме рельефа самому мозгу, чем у низших позвоночных (у рыб, земноводных и пресмыкающихся полость мозговой коробки черепа обычно больше самого мозга, иногда значительно, за счет разращения мозговых оболочек и крупных синусов - резервуаров крови) (см. выше). В настоящее время палеонтология позвоночных, необычайно расширив наши знания об органическом мире в целом, вносит существенный вклад в сравнительную анатомию головного мозга, нередко подтверждая своими историческими документальными данными некоторые предположения сравнительной анатомии и эмбриологии. Например, можно указать на недавнее обнаружение у девонской панцирной рыбы титаних-тис двух теменных глаз, о чем для древних челюстноротых сравнительная анатомия и эмбриология позвоночных могли лишь строить предположения (Т. Эдингер, 1955).
Изучение головного мозга ископаемых позвоночных в (ряде случаев оказалось существенным для установления их систематического положения, при этом систематических категорий самого разного масштаба. Достаточно упомянуть классические работы Э. Стенше о древнейших бесчелюстных и их головном мозге, или выявившееся недавно сходство во всем существенном головного мозга ископаемого млекопитающего десмостилус, загадочного долгие годы, с мозгом современных сиреневых (дюго ней и ламантинов). Введение характеристики головного мозга ископаемых позвоночных в систему их крупных таксономических категорий и систематику имеет не меньше основания, чем обычная характеристика по скелету, хотя практически это пока по разным причинам редко делается даже для современных по звоночпых, не говоря 06 ископаемых...
Весьма существенно изучение головного моета разных по геологическому возрасту представителей одной и той же группы позвоночных. Эти сведения дают дополнительное освещение эволюции группы, уточняют ее филогению и наши представления об эволюционном уровне тех или иных представителей. Так, изучение головного мозга археоптерикса показало, что его мозгу присущи черты строения, свойственные пресмыкающимся в гораздо большей степени (12 признаков), чем характерные для типичных птиц (4 признака). С другой стороны, летучие ящеры-птерозавры выработали мозг, построенный по типу птичьего, задолго до настоящих птиц (Т. Эдингер, 1955).
Сказанное выше не предполагает переоценки скромных возможностей изучения по слепкам головного мозга ископаемых позвоночных. Следует соблюдать осторожность и иметь в виду возможность ошибок при сопоставлении мозга по одним отливам. Для головного мозга современных млекопитающих известны случаи вариаций борозд и извилин не только в пределах одного и того же вида, но и одного и того же индивида для правого и левого полушарий его большого мозга. Таким образом, необходимо стремиться учитывать индивидуальную изменчивость. Нужна осторожность и в гомологизации извилин и борозд у разных форм, особенно, если нет достаточно данных об их родственных взаимоотношениях, например, по скелету. Надо, наконец, учитывать явления конвергенции в строении, интересным примером чего для головного мозга может служить мозг летучих ящеров и птиц.
Однако осторожность не должна снижать актуальности изучения головного мозга ископаемых животных по слепкам. Палеонтология дала нам ряд интересных примеров, па первый взгляд, быть может, несколько неожиданных, когда на раннем этапе эволюции головной мозг вымершей группы достигал сравнительно высокого уровня развития, не меньшего, чем у доживших до более позднего времени родичей ископаемых животных, или даже, чем у доживших до современности. И тем не менее эти, казалось бы, "счастливые" первые обладатели высоко развитого мозга вымерли, не оставив никакого потомства. Примеры такого рода можно найти среди копытных, хищников и др. Таким образом, при всей общепонятной важности головного мозга для животных в их эволюции, борьбе за существование и т. д. - не в одном головном мозге дело... Это лишний раз говорит о необходимости всегда стремиться к изучению организма как целого во всех его взаимосвязях с условиями существования - насколько это позволяет данный ятац науки. Но из этого же вытекает и необходимость возможно детального изучения всех доступных частей ископаемого организма, в том числе и головного мозга, в меру возможностей представляемых остатка ми органической жизни минувших эпох и сравнительными данными для современных животных. И если до последнего времени палеонтология, в частности систематика, ископаемых млекопитающих оставалась в значительной степени палеонтологией их зубов и конечностей, то теперь, при разработанности учения oо работе головного мозга на основе данных И. П. Павлова и других настало время прилагать все усилия к введению в палеонтологию и сведений о головном мозге.
Вопросы эволюции мозга настолько существенны для пани мания эволюции позвоночных вообще, что даже незначительные данные могут многое уяснить палеонтологу; а в будущем, при oболее углубленном изучении мозга современных позвоночных и при усовершенствовании методов изучения мозга ископаемых, палеоневрология, без сомнения, станет играть большую и плодотворную роль в познании жизети прошлого и ее истории.
Указатель русских и латинских названий
Буква "А"
Аллацеpoпс - Allacerops turgaicus (Boriss.)
Альцпцсфалус - Alcicephalus
Амблиптерус - Amblypterus
Амблистома - Amblystoma
Аминодон - Amynodon mongoliensis Osborn
Аминодопты - Amynodontidae
Анкилозавр - Ankylosaurus
Анна петри (тероцефал) - Anna petri Amalitzky
Анпиды - Annidae Антодон - Anthodon rossicus Hartmann-Weinberg
Аптракозиды - Antracosidae
Анхитерий - Anhitherium aurelianense (Cuvier)
Архелоп - Archelon isliyros Wiel
Археоламбда - Archaeolambda planicanina Flerov
Ахтиария - Achtiaria
Буква "Б"
Барсук паратаксидэа - Parataxidea
Барсук североамериканский, современный - Taxidea
Батрахозавры - Batrachosauria
Бептозух - Benthosuchus
Бритоподиды - Brithopodidae Efremov
Буква "В"
Венгоковия - Venjukovia invisa Efremov
Буква "Г"
Гадрозавры - Hadrosauria
Газель (из Пикерми) - Gasella deperdita Gaudry
Гигантаминодон - Gigantamynodon cessator Gromova
Гиена - Hyaena (Crocuta) variabi lis Zdansky
Гиена-крокута - Crocuta eximiа Roth, et Wagner
Гиенодоп - Ilyaenodon compressus Filh., Hyaenodon aymardi Filh., Hyaenodon ambiguus Schl.
Гиппарион (род) - Hipparion
Гиппарион длинноногий - Hipparion longipes Gromova
Гиппарион малый - Hipparion elegans Gromova
Гиппарион североамериканский - Hipparion gratum
Гипсаминодон - Hypsamynodon progressus Gromova
Гобиоцерас - Gobiocerus mongoli cus Sokolov
Гониофолисы - Goniopholidae
Буква "Д"
Двиния - Dvinia suschkini (Amalitzky) Permocynodon
Двинозавр - Dvinosaurus primus Amalitzky
Десмостилус - Desmostylus
Диноцераты-монголотерии - Mongolotherium plantigradum Flerov
Дипловертебронтиды - Diplover-tebrontidae
Дицинодонт - Dicynodon traut-scholdi Amalitzky
Долиозапр - Doliosaurus yanshino vi Orlov
Буква "Ж"
Жирафы собственно - Giraffinae Завроподы - Sauropoda
Буква "З"
Зауроктон - Sauroctonus progressus Ilartmann - Weinberg
Зауролоф - Saurolophus angustirostris Rozhdestvensky
Буква "И"
Игуанодон - Iguanodon
Игуанодон бельгийский - Iguanodon bernissartensis Boulenger
Игуанодон монгольский - Iguanodon orientalis Rozhdestvensky
Иктитерий - Ictitherium hipparionum (Gervais)
Иностранцевия - Inostrancevia alexandri Amalitzky
Буква "К"
Кадуркодон - Cadurcodon ardynensis (Osborn)
Карпозавры - Carnosauria
Котлассия - Kotlassia prima Amalitzky
Креодонты-гиенодонты - Hyaenodon dubius Filh.
Куница - Martes palaeosinensis (Mustella palaeosinensis Zdansky)
Куница, лесная современная - Martes martes
Буква "Л"
Лаптанозух - Lantanosuchus qua-leni Efremov
Лепидостеусы, "панцирные щуки" - Lepidosteus
Буква "М"
Мастодонт Борсона - Mastodon borsoni
Махайрод - Machairodus irtyschensis
Мезозухии - Mesosuchia
Мерикгиппус - Merychippus
Мерикотерий - Merycotherium sibiricum
Миомериксы - Myomerix
Мунтжаки - Muntjacus
Буква "Н"
Никтеролетер - Nycteroleter ineptus Efremov
Никтифрурет - Nyctifruretus acudens Efremov
Буква "О"
Окапи - Okapia johnstoni
Олепь павлодарской фауны - Cervavitus orlovi Flerov
"Оленьки" трагулиды - Tragulidae
Орасиус - Orasius Wagner
Орнитоподы - Ornithopoda
Буква "П"
Палеоскаптор - Palaeoscaptor
Палеотраги - Palaeotraginae
Пантодонты - Pantodontia-Amblypoda
Парабрадизавр - Parabradysaurus udmurticus Efremov
Паракамелюс - Paracamelus
Параллигаторы - Paralligator gradillifrons Konjukova
Парейазавры - Pareiasauridae
Парейязухус - Pareiasuchus vjatkensis Hartmann-Weinberg
Пермоцинодон - Permocynodon
Перупиум - Perunium ursogulo Orlov
Плезиогуло - Plesiogulo brachygnalhus Zdansky
Попугай - Psittacus
Православлевия - Pravoslavlevia parva Vjuschkov
Продиноцерасы - Prodinoceras martyr Matt., Gr. et Simps.
Протемболотерий - Protembolotherium efremovi Janovskaja
Протоцератопс - Protoceratops andrewsi Brown et Schlajkier
Пситтакозавр - Psittacosaurus mongoliensis Osborn
Буква "С"
Саблезуб - Machairodus ishimicus
Самотерий - Samotherium
Санпазавр - Sanpasaurus
Сеймуриаморфы - Seymouriamor-pha
Семантор - Semantor macrurus Orlov
Семанториды - Semantoridae Orlov
Серридептины - Serridentinus to-logojensis Beliajeva
Сиватерии - Sivaiherinae
Сиодон - Syodon efremovi Orlov
Сиреневые (дюгони, ламантины) - Sirenia
Сирмозавр - Syrmosaurus vimini-caudatus Maleev
Сколозавр - Scolosaurus cutleri Nopcsa
Скутозавр - Scutosaurus karpin-skii Hartmann-Weinberg
Страус херсонский - Slrutio chersonensis Brandt
Сушкиния, сокол - Suchkinia
Схизотерий - Schizotherium avitum Matth. et Gr.
Сцелидозавр - Scelydosaurus
Буква "Т"
Таларурус - Talarurus plicatospineus Maleev
Тарбозавр - Tarbosaurus hntaar Maleev
(Тираннозавр - Tirannosaurns rex Osborn)
Титапофопеус - Titanophoneus potens Efremov
Tparouepoc - Tragoceros a malleus Gaudry
Трпониксы - Trionychidae
Трифозух - Triphosuchus pauci dens Konjukova
Трубкозуб - Orycteropus
Буква "У"
Улемозавр - Ulemosaurus svijagensis Rjabinin
Буква "Ф"
Фринозома - Phrinosoma cnrnutum
Фтинозухиды - Phtinosuchidae
Буква "Х"
Херсоттотерий - Chersonotherium eminens
Хилотерий - Chilotherium
Хонанотерий - Honanotherium Bohlin
Хтопозавр - Chtonosaurus veloei dens Vjuschkov
Буква "Ц"
Цополофус - Caenolophus promissus Matthew et Granger
Цепвавитус - Cervavitus orlovi Flerov
Цетотерий - Cetotherium
Буква "Ш"
Шталекерия - Stalekeria potens v. Huene
Буква "Э"
Эласмотерий - Elasmolherium sibiricum Fish.
Элафодус - Elaphodus Элурогпат - Aelurognathua
Энозух - Enosuchus brcvicep Efremov
Эптрлодон - Entelodon
Эргилия - Ergilia pachypterna Grоmova
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'