
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Глава шестая. Верхнепермские пресмыкающиеся Поволжья и Северной Двины
"Мир прошедший, покорный мощному голосу науки, поднимается из могилы свидетельствовать о переворотах, сопровождавших развитие поверхности земного шара; почва, на которой мы живем, эта надгробная доска жизни миновавшей, становится как бы прозрачною; каменные склепы раскрылись; внутренности скал не спасли хранимого ими. Мало того, что полуистлевшие, полуокаменелые остовы обрастают снова плотью, палеонтология стремится раскрыть закон соотношения между геологическими эпохами и полным органическим населением их. Тогда всё некогда животе воскреснет в человеческом разумении, всё исторгнется от печальной участи бесследного забвения, и то, чего кость истлела, чего феноменальное бытие совершенно изгладилось, восстановится в светлой обители науки, в этой области успокоения и увековечения временного".
Герцен А. "Письма об изучении природы". Полн. собр. соч., т. IV, стр. 165.
Пермский период - один из наиболее интересных в истории пресмыкающихся и в истории наземной жизни вообще.
Аридизация климата на значительных участках континентов, особенно в верхней перми, и дифференцировка суши сравнительно с предшествовавшим каменноугольным периодом были несомненно весьма важными факторами в истории развития и в характере распространения пермской флоры. За флорой следовала в своем (распространении и в своих изменениях и наземная фауна с ее сложными цепями питания и возраставшими в своем разнообразии "нишами жизни".
Что касается самих пресмыкающихся, то из предшествующей каменноугольной эпохи до нас дошли сравнительно немногочисленные остатки этого когда-то обширнейшего класса позвоночных. Пермский же период оставил, как известно, богатейшее наследие. Пресмыкающие достигли в это время, особенно в позднепермскую эпоху, замечательного разнообразия в своих приспособлениях к окружающей среде и в направлениях своей эволюции до млекопитающеподобных включительно. Эта эпоха была, видимо, и временем зарождения бесчисленных пресмыкающихся мезозоя.
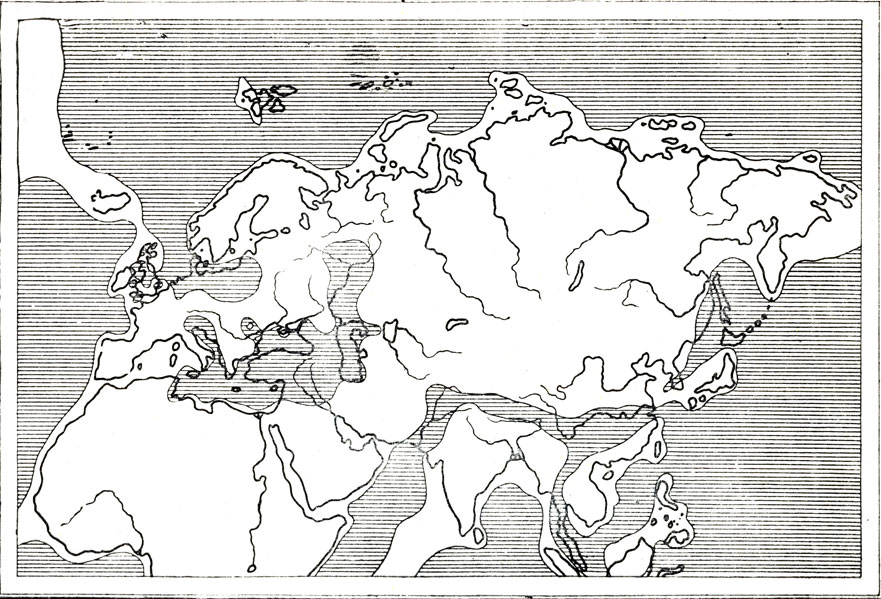
Рис. 72. Схема палеографии верхней перми для части восточного полушария (по Темье, 1952)
И сами палеонтологические материалы из пермских отложений разных (Материков, и результаты изучения этих материалов представляют собою в настоящее время итог огромного труда многих поколений ученых разных стран, богатейшее общее достояние единой мировой науки: по своему существу палеонтология, эта "зоология и ботаника угасших форм жизни", как ее называл академик А. П. Павлов, еще более международна, чем зоология современности. Но по специфике палеонтологии, из-за неполноты геологической летописи, по палеозоогеографическому, палеоклиматическому и т. д. разнообразию разных стран, вклад каждой страны в это общее дело, естественно, очень различен. Он часто различен по своему основному исходному материалу в виде остатков животных, но не редко и по выводам на основе изучения этих документов далекого прошлого, а также местонахождений ископаемых остатков.
Не имея возможности подробно останавливаться на истории изучения пермских пресмыкающихся СССР, отметим лишь, что первые упоминания о находках в России относятся еще к XVIII в., изучение же наших пермских рептилий началось в первой половине XIX в. и связано с именами С. Куторги (1838, 1842, 1844), Г. Фишера (1840-1847), Э. Эйхвальда (1861). На рубеже XIX и XX вв. (1895-1903) были поставлены крупные раскопки В. П. Амалицкого (1860-1917) в верхнепермских отложениях р. Северной Двины; итог раскопок - огромные палеонтологические сборы верхнепермских пресмыкающихся, а также земноводных. Эти работы впервые установили близость найденной фауны к южноафриканской из "формации Карру" и послужили научной и организационной основой для последую щего систематического изучения палеозойских пресмыкающихся и земноводных. Это стало реальным только после Октябрьской революции, когда огромное плановое развитие всей жизни Советского Союза и его науки, в том числе естествознания, обеспечило возможность быстрого роста и палеонтологии позвоночных. Он связан с именами наших выдающихся ученых - академика П. П. Сушкина (1868-1928) и в особенности основателя и директора Палеонтологического института Академии наук СССР А. А. Борисяка (1872-1944). Непосредственным планированием и выполнением палеонтологических экспедиций для сборов ископаемых палеозойских и мезозойских пресмыкаюшихся и земноводных, их изучением - лично и силами сотрудников - мы почти целиком обязаны И. А, Ефремову; в его работах (1954 и др.) содержится интересный очерк истории исследования низших четвероногих нашей территории. Настоящий очерк имеет целью дать краткий обзор итогов изучения некоторых пермских пресмыкающихся СССР за последнее полстолетие. При этом имеются в виду пресмыкающиеся верхней перми: именно верхнепермские отложения СССР дали наиболее разнообразный интересный материал. Изучение наших нижнепермских пресмыкающихся находится пока еще в начальной стадии. Обзор всего обнаруженного и изученного за последние годы в СССР вышел бы далеко за пределы настоящего очерка. Имеется в виду рассмотрение лишь двух фаун, точнее "фаунистических комплексов" пресмыкающихся. Одна из них происходит из континентальных отложений, соответствующих верхней части казанского яруса морских отложений и в общем по своему характеру и стратиграфическому положению зоне Tapinocephalus серии Бофорт Южной Африки (И. А. Ефремов, 1937-1954). Другая происходит из верхнепермских отложений, относимых к татарскому ярусу (соответствует зоне Cistecephalus Южной Африки) - из слоев, по-видимому, отделенных от содержащих фауну казанского времени перерывом в осадкообразовании.

Профессор В. П. Aмалицкий (1860-1917)
Остановимся на верхнепермской фауне пресмыкающихся и рассмотрим фауну села Ишее'ва, около г. Тетюши по среднему течению Волги.
Высокий правый берег Волги около г. Тетюши сложен верхнепермскими песками и глинами с прослоями известняков. Местами все они смяты в причудливые складки, привлекающие внимание пассажиров волжских пароходов, но в основном забегают горизонтально. Западнее, километрах в тридцати, верхнепермские отложения, относимые к казанскому ярусу, выступают в глубоких оврагах, впадающих в живописную долину р. Свияги, которая течет меридионально, как и Волга, но с юга на север.
Здесь, в верхнепермских песках, летом 1929 г. студент-геолог Носов нашел скелет ископаемого позвоночного. Место находки - верховье каменного Оврага, выходящего в речку Сухую Улему - пересыхающий летом правый приток Свияги, около села Ишеева. Скелет не удалось добыть - хрупкие кости рассыпались. К счастью, находка была сфотографирована местным краеведом Князевым, и этот снимок, опубликованный позднее в журнале "Природа", позволил установить принадлежность скелета древнему хищному ящеру (Б. А. Штылько и А. Н. Рябинин, 1932). В 1930-1931 гг. здесь вел раскопки центральный геологоразведочный институт (ныне ВСЕГЕИ, Ленинград), а в 1934, 1935 и 1939 гг. Палеонтологический институт Академии наук СССР (рис. 73). Эти работы обнаружили остатки интереснейших верхнепермских пресмыкающихся, а также земноводных и рыб. Уже первые предварительные описания И. А. Ефремова (1937-1940) были так интересны, что быстро вошли в мировую науку, а скелеты и черепа, украшающие музей Палеонтологического института Академии наук СССР в Москве, могут считаться гордостью этого учреждения.

Академик П. П. Сушкин (1868-1928)
Раскопки около Ишеева дали главным образом древних терапсид - пресмыкающихся из числа обладавших одной височной впадиной и дугой ("подкласс" синапсиды). Для древних тераисид - титанозухий и тапиноцефалов - характерны высокая, как бы вдавленная затылочная сторона черепа, большое отверстие непарного теменного глаза на возвышении теменных костей (рис. 74, 75, 79, 80); нёбо имеет древний тип строения ("первичное").

Рис. 73. Раскопки в Каменном овраге около Ишеева
Для удобства изложения начнем рассмотрение этой фауны с титанозухий, хищных терапсид семейства бритоподид (Brithopodidae Efremov, 1954). Это пресмыкающиеся, напоминающие южноафриканских титанозухий, но более примитивные. Как и улемозавр (см. ниже), они еще сохранили многие черты(рис. 74-77) своих предков, сфенакодонтных пеликозавров, известных из нижнепермских красноцветных толщ Северной Америки.
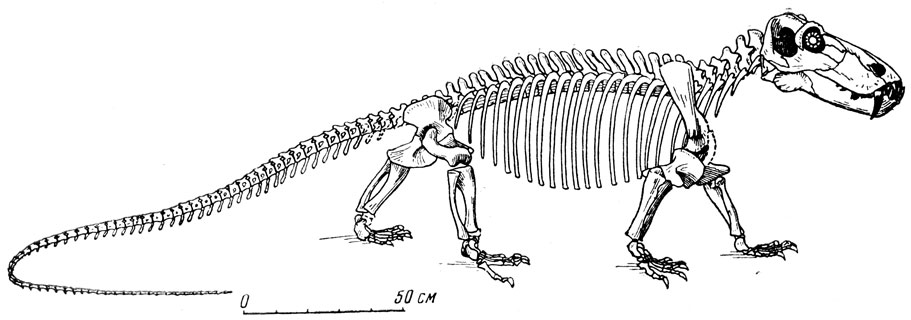
Рис. 74. Титанофонеус (Titanophoneus potens Efremov) Скелет. Верхняя пермъ, Татарская АССР Музей Палеонтологического института АН СССР
Наиболее полно изученный титанозух ишеевской фауны - титанофонеус (греческое - "титанический убийца"). В Музее Палеонтологического института хранится почти полный скелет этого пресмыкающегося длиною в три метра (рис. 74). Череп (рис. 75, а) высокий, задняя часть черепа, очень характерная для титанозухий, с отверстием теменного глаза, лежащем на заднем краю черепа на высоком бугре, образованном теменными костями. Нерв теменного глаза был при этом довольно длинным. Затылочная сторона черепа высокая и широкая, что говорит о сильной шейной мускулатуре. Большой срединный вертикальный затылочный гребень указывает на сильную срединную затылочную связку, что отчасти связано с тяжелым черепом (она облегчает своей упругостью приведение головы из опущен ного положения в нормальное). Большая височная впадина и сильное развитие гребней на окружающих ее костях, например на теменных, указывают на мощность височной мускулатуры и, вероятно, на стремительное и сильное захлопывание пасти. Глазные впадины титанозухий направлены косо в стороны и довольно сильно вперед, что для хищника важно при высмат ривании добычи. Глазное яблоко снабжено кольцом плоских, плотно сочлененных друг с другом косточек, которые увеличи вали устойчивость склеры (белочной оболочки глаз). На переднем конце черепа, сжатого с боков, нижний край морды поднимается вперед и вверх, что придает черепу некоторую бульдогообразность; это унаследовано от пижнепермских пеликозавров и, вероятно, связано со способом хватания добычи. Острые резцы направлены косо вперед, особенно верхние, очень плотно входящие в промежутки между нижними (как и нижние между верхними) (рис. 75, а). Точность в смыкании челюстей, без которой животное при стремительном хватании добычи могло бы сломать себе зубы, обеспечена строением челюстного сустава (глубокий двойной блок вверху и двойная впадина в нижней челюсти), возможно, и сильным развитием поперечных отростков крыловидных костей, вероятно, с упором их изнутри в нижнюю челюсть при закрывании рта (рис. 76). Боковые движения нижней челюсти были при этом совершенно исключены. Клыки изогнуты назад и имеют большие корни, глубоко сидящие в челюстях. Задний край клыка снабжен острым режущим гребнем, который мелко зазубрен, как и гребни, отходящие от вершин всех других зубов к их основаниям (такая зазубренность характерна для зубов многих хищных животных, например крупных акул, многих древних пресмыкающихся, ископаемых кошек - "саблезубов" и др.). Послеклыковье, щечные зубы - режущие, но редкие, небольшие, частью отогнуты назад (рис. 75, б; 76). В верхней челюсти для вершин нижних клыков имеются особые углубления.
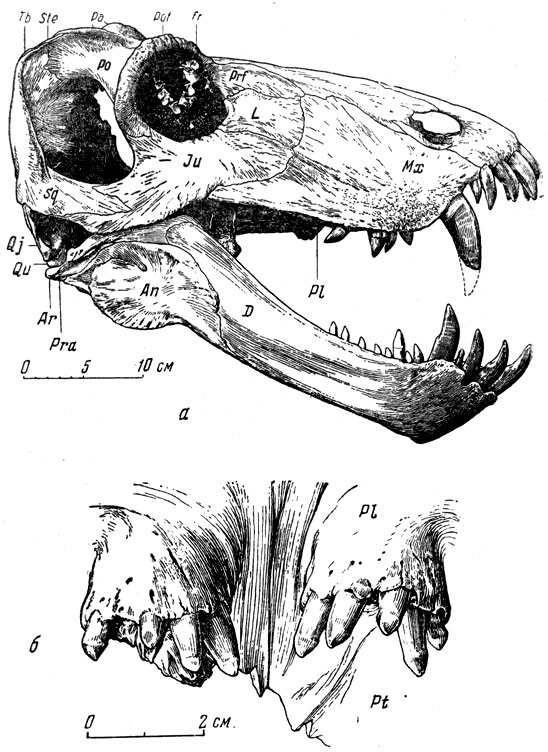
Рис. 75. Титанофонеус (Titanophoneus potens Efremov) a - череп с нижней челюстью, вид сбоку; б - бугры нёбных костей с зубами, вид снизу и спереди. Верхняя пермь, Татарская АССР (Ю. А. Орлов, 1958)
Своеобразно строение небных когтей - каждая снабжена бугром, из которого торчит вниз около десятка небольших цилиндрических зубов с вершиной в виде тупого конуса (рис. 75, б, 76). Массивные поперечные ветви крыловидных костей, примыкающих сзади к небным, внешними утолщенными концами опускаются далеко вниз. На их нижней стороне у титанофонеуса также по два-три зуба, сходных с небными, но посаженных довольно странно - почти горизонтально, вершинами в стороны ветвей нижней челюсти.
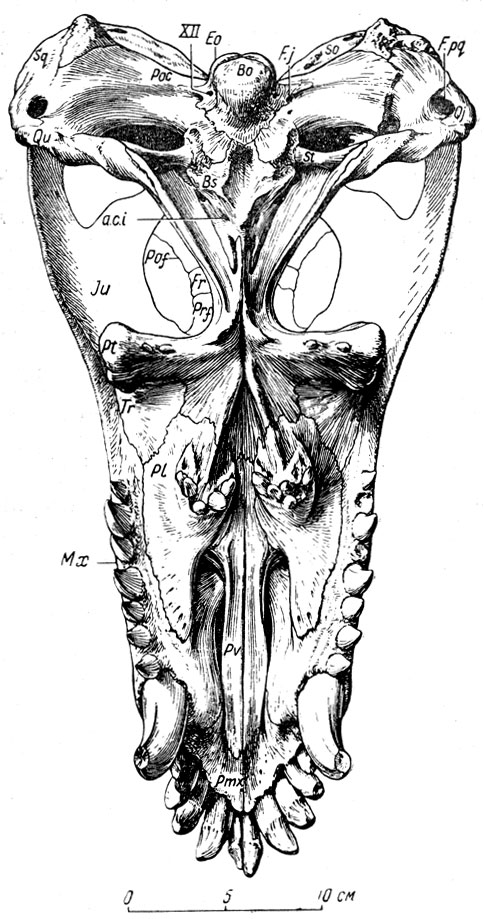
Рис. 76. Титанофонеус (Titanophoneus potens Efremov) Череп, вид снизу. Верхняя пермь, Татарская АССР
Затылочный суставнюй бугор отогнут вниз, очевидно, голова обычно была наклонена концом морды довольно сильно вниз.
Неясно восприятие звуков титанофоиеусом. Столбик уха (stapes, "стремечко") имеет вид небольшой распорки в основании черепа между квадратной костью и мозговой коробкой, где он упирается в овальное окно. Это положение столбика как будто бы исключает его подвижность и приспособленность для передачи звуковых колебаний через овальное окно во внутреннее ухо, хотя по местоположению такой столбик равнозначен стремени среднего уха млекопитающих, птиц и пресмыкающихся, имеющих барабанную перепонку и воспринимающих звук. Как воспринимали звук титанофонеус и другие пресмыкающиеся с таким строением - неясно, но вряд ли они были глухие. Впрочем, змеи по-своему "слышат", хотя лишены барабанной перепонки, и "столбик уха" у них прежде всего - механическая конструктивная часть черепа, а не слуховая {см. ниже).
Позвоночный столб титапофонеуса сравнительно с черепом стройный и легкий. В нем около сотни двояковогнутых позвонков с тонким отверстием для хорды. Шейный позвонок, следующий за атлантом (эпистрофой), снабжен массивным остистым отростком. Это, вероятно, также связано с большим весом тяжелой головы, что требовало сильной затылочной связки и мускулатуры. Тела позвонков на переднем и заднем концах с округлыми внешними краями, что указывает на гибкость животного и его подвижность. Три крестцовых позвонка соединены неподвижно с тазом крестцовыми ребрами. Хвост длинный, в нем около 60 позвонков из всех ста. Подвздошная кость по строению промежуточна между таковой древнейших пресмыкающихся, где она вытянута назад, и подвздошной более поздних млекопитающеподобных рептилий, у которых верхний отдел кости вытянут вперед, как у млекопитающих.
Передние и задние конечности титанофонеуса одинаковой длины, сильные, стройные, хорошо подвижные; плечо с предплечьем, бедро и голень очень напоминают крокодильи; при этом лапы относительно роста животного очень широкие, главным образом за счет пястных и плюсневых костей, увеличивающихся от первого пальца к пятому (рис. 77). Большая величина лап особенно заметна, если сравнить титанофонеуса с крокодилом, даже вдвое или втрое большим, чем титанофонеус, У титанофонеуса цри наличии перепонки между пальцами - это "почти ласты", скорее всего предназначенные для плавания, а также передвижения по влажному и вязкому, илистому побережью рек и озер. Крокодилы обычно плывут вперед, работая высоким хвостом, сжатым с боков наподобие весла, а также боковыми изгибами тела. Длинным хвостом титанофонеус, вероятно, мог наносить сильные удары, как это делают круп ные ящерицы и крокодилы, но для плавания хвост не был приспособлен. Титанофонеусу могли давать быстроту в воде его расширенные на концах передние и задние лапы, а также боковые изгибы тела. Число фаланг в пальцах на первый взгляд то же, что и у многих млекопитающеподобных пресмыкающихся и млекопитающих, т. е. та же "фаланшвая формула" - 2. 3. 3, 3. 3, от первого пальца к пятому. Но вторая фаланга четвертого пальца передней лапы срослась из двух: из них одна, сочленяющаяся с первой основной фалангой пальца, со-всем короткая, а следующая, сочлененная с когтевой, сохранила длину, близкую к нормальной (рис. 77). Очевидно, четвертый палец близких предков титанофонеуса еще "недавно" обладал четырьмя самостоятельными фалангами.
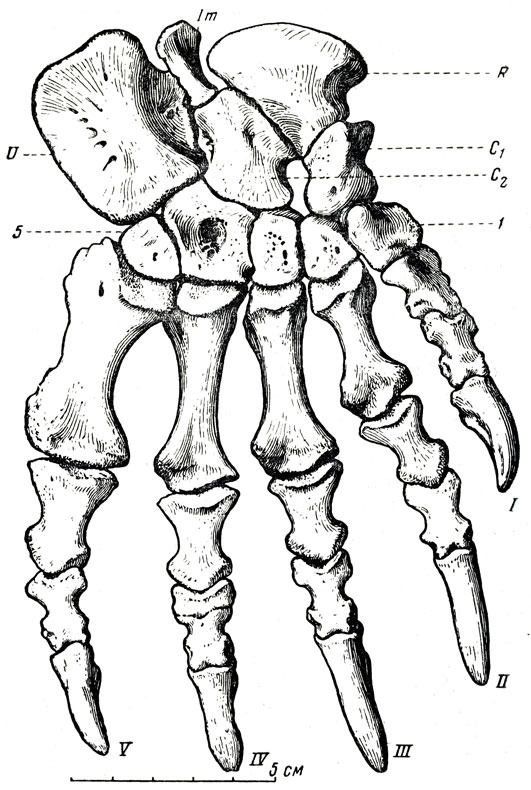
Рис. 77. Титанофонеус (Titanophoneus potens Efremov). Правая кисть: U - ulnare; Imintermedium; R - radiale; C1C2 - carpalia. Верхняя пермь, Татарская АССР (Ю. А. Орлов, 1958)
Титанофонеус явно хищник, но вряд ли часто охотился на своих крупных сородичей, растительноядных тапиноцефалов, улемозавров (рис. 80-82) и вообще на такую крупную добычу. Верхние резцы титанофонеуса, отчасти и нижние, направлены косо вперед; верхний клык, хотя изогнут назад, но не так велик, чтобы мог считаться предназначенным для разрывания на части взрослых улемозавров, величиной с самого титанофонеуса, или крупнее его. Сравнительно слабые послеклыковые зубы, хотя и режущие, но редкие, мало удобны для измельчения мышц, связок и грубой кожи крупных пресмыкающихся, а скорее всего приспособлены для удержания добычи. Из всех зубов наиболее мощные - клыки и цредклыковые зубы, видимо, по преимуществу, хватательные, хотя гребни на краях верхних резцов, заходящие за края нижних, делали резцы одновременно и режущими. Небольшой уступ на внутренней, язычной стороне резца, в который упиралась вершина зуба противоположной челюсти - "пятка", делал резцы отчасти измельчающими пищу, однако у титанофонеуса лишь в слабой степени. Наиболее совершенны в этом отношении резцы улемозавров (рис. 81) (И. А. Ефремов, 1940).

Титанофонеусы и улемозавры. Рис. К. К. Флерова
Небные зубы (рис. 75, б), торчащие вершиной вниз из бугров небных костей, при отсутствии противопоставленных им внизу, пе приспособлены для жевания. Вероятно, мы имеем дело с каким-то своеобразным раздавливающим, а частично и удерживающим добычу приспособлением:. Для раздавливания добычи могли служить отчасти и поперечные массивные отростки птеригоидов.
Если принять во внимание и "крокодилье" строение плеча с предплечьем, бедра с голенью и широкие лапы титанофонеуса, то можно прийти к предположению о приспособленности животного к полуводному образу жизни и питанию рыбой с плотной чешуей (остатки рыб в этом местонахождении довольно часты), моллюсками с тонкой раковиной, мелкими земноводными и пресмыкающимися, а также другой некрупной добычей. Титанофонеус мог ее ловить и высматривая с берега, что было удобно при обычно наклонном положении головы, мордой книзу (рис. 74), и в воде, так как он, вероятно, был хорошим пловцом. Разумеется, титапофонеусы, собравшись вместе, были небезопасны и для улемозавров (см. ниже), кормившихся растительностью на побережье верхнепермских озер и рек, и могли стащить в воду и разорвать зазевавшегося улемозавра, но по своему образу жизни вряд ли были повседневными врагами крупных улемозавров, вообще крупной добычи.
Другой, близкий к титанфонеусу, но более грозный титанозух из раскопок в Каменном Овраге - хищный долиозавр (греческое долиозавр - коварный ящер) (рис. 78), принятый вначале за большого старого самца титанофонеуса. Известно, что многие крупные низшие (позвоночные (крокодилы, черепахи, щуки и др.) очень долговечны, а главное обладают длительным ростом уже во взрослом, половозрелом состоянии. Поэтому разница между Долиозавром и титанофонеусом в размерах сначала не казалась сама по себе существенной. Позднее выяснились отличия в строении черепа, зубов и т. д., и "старый самец титанофонеуса" оказался новым очень крупным хищным титанозухом.
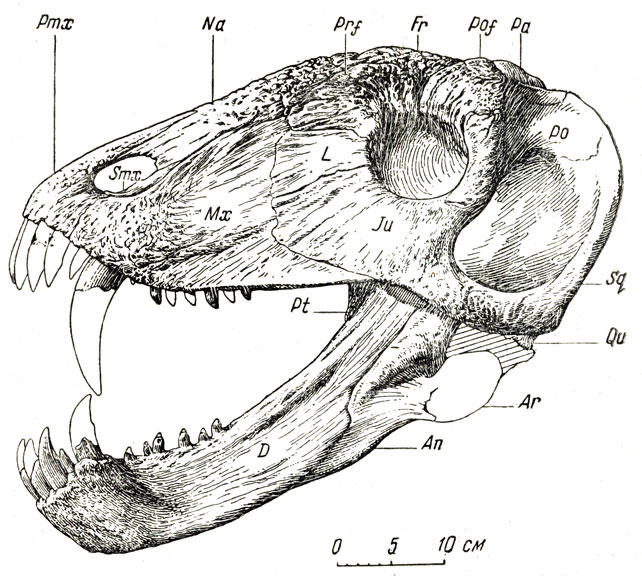
Рис. 78. Долиозавр (Doliosaurus yanshinovi Orlov). Череп, вид сбоку. Верхняя пермь, Татарская АССР (Ю. А. Орлов, 1958)
Тяжелый, высокий череп долиозавра достигает в длину более полуметра. Многие кости верхней стороны черепа очень утолщены (особенно лобные, окружающие гладницу, и теменные) ; снаружи они сильно бугристы с бороздами кровеносных сосудов и отдаленно напоминают бугристость костей, несущих рога, например у носорогов из млекопитающих. Вероятно, у долиозавра это указывает на очень сильное ороговение кожи защитного характера. Морда, сжатая с боков, высокая; глазничные впадины, окруженные массивными костями, невелики. Резцы и особенно клыки крупные, в основном приспособленные для хватания добычи при стремительном захлопывании пасти, па которое указывают большие височные впадины, вмещавшие челюстную мускулатуру. Послеклыковые зубы слегка копьевидны, малы, не образуют сомкнутого ряда, немногочисленны и скорее предназначены для удержания добычи, чем для ее измельчения. Небные бугры невелики, с мелкими тупоконическими зубами; позади небных бугров расположено по два-три таких же зуба на птеригоидах; зубы на поперечных отростках птеригоидов мелкие. Помимо впадин, для нижних клыков (впереди верхних), что характерно для многих хищных титанозухов и родственных им пресмыкающихся, на небе долиозавра имеются ямки и для нескольких послеклыковых, "щечных" зубоов. Их вершины должны были входить в эти небные ямки довольно точно во избежание поломки, а объединенность ямок в одном общем желобообразном углублении, быть может, указывает на возможность небольших, но только передне-задних движений - нижней челюсти; об этом же говорит и несколько уплощенная форма челюстного сустава. При захлопывании пасти нижние послеклыковые зубы ложились внутрь от верхних, не соприкасаясь с ними. Это подтверждает предположение о приспособленности щечных зубов для удержания добычи, а не ее измельчения.
Не совсем понятна необычайная (для размеров животного) массивность лобной и теменной частей черепа. Частично это можно было бы толковать как итог приспособления к большой механической нагрузке со стороны переднего конца морды при хватании и удержании крупной добычи. Однако это не объясняет странного бугристого строения внешней поверхности костей верхней стороны черепа. (Существует предположение, что большой толщины кости крышки черепа служили и защитой от перегрева в жаркое время, но это допущение не особенно убедительно). Остальной скелет в общем похож на скелет титанофонеуса, но крупнее и гораздо массивнее. Долиозавр был явно хищником, но характер его пищи не совсем ясен. Более массивный и, видимо, менее подвижный, чем титанофонеус, долиозавр вряд ли был особенно проворным, активным охотником за рыбой и другими водными животными, которых, правда, мог и подстерегать. Описываемая фауна была богата, кроме всего прочего, небольшими мелкими земноводными и пресмыкающимися; для них в то время долиозавр, вероятно, был поистине "царем зверей". Впрочем, известно, что хищные пресмыкающиеся, даже крупные, например огромные удавы и крокодилы, с их низкой, переменной температурой крови гораздо менее прожорливы, чем это может показаться на первый взгляд.
Небольшой хищный титанозух сиодон (рис. 79) с черепом всего 20 см длины, казалось бы, очень походит на титанофонеуса. Но теменной глаз у сиодона относительно крупнее; его отверстие лежит на тонкостенном бугре, относительно более высоком, чем у титанофонеуса. Верхние клыки сиодона изогнуты назад сильнее, в поперечнике округлые, без режущего заднего края, без острой колющей вершины. Послеклыковых зубов много, они низкие, тупоконические, менее сжатые с боков, стоят прямо, часто, в отличие от зубов титанофонеуса, хорошо приспособлены для измельчения пищи. Небные бугры с зубами, как и поперечные отростки птеригоидов, относительно сильнее, чем у титанофонеуса. Но особенно резко отличны все послеклыко-вые челюстные зубы. Вдобавок задние из них иногда стоят в нижней челюсти не только друг за другом, но и рядом, попарно. Такая "пара" однобугорчатых зубов - своего рода заменитель, хотя и не очень совершенный, одного, но бугорчатого зуба некоторых пресмыкающихся (более поздних, триасовых), а также млекопитающих. По-видимому, сиодон мог быть всеядным животным, хотя в основном, вероятно, охотился за разными мелкими животными. Большие глазницы наводят на предположение о значительной величине глаз и о сумеречном или ночном образе жизни этого животного.
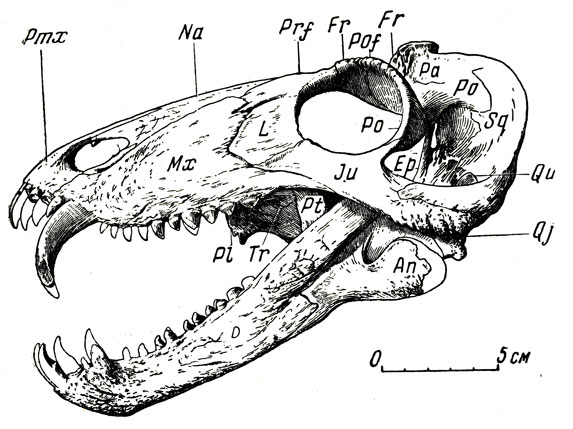
Рис. 79. Сиодон (Syodon efremovi Orlov). Череп, вид сбоку. Верхняя пермь, Татарская АССР (Ю. А. Орлов, 1958)
Добыты остатки более дюжины особей сиодона, но, к сожалению, ни одного цельного скелета; видимо, этому животному принадлежат многочисленные кости конечностей и позвонки небольшого титанозуха ишеевокой фауны. По костям конечностей и позвонкам сиодон походил на титанофонеуса, но был много меньше его (около полутора метров в длину). Лопатка у титанофонеуса и у сиодона изогнута очень слабо; между тем ее кривизна при близком (расположении лопатки к грудной клетке в известной степени отражает форму клетки: вполне возможно приписать титанофонеусу и сиодону сжатое с боков туловище, а не уплощенное сверху вниз, как обычно у крокодилов.
Крупное растительноядное пресмыкающееся этой фауны - тапиноцефал улемозавр (рис. 80) (Наиболее полный материал был первоначально изучен А. Н. Рябининым (1938). Подробное описание черепа с рисунками А. П. Быстрова дал И. А. Ефремов (1940). Название животное получило по речке Улеме, где был найден череп). Череп улемозавра имеет очень высокую, наклонную назад затылочную сторону; верхний край глазницы так сильно утолщен, что сбоку кажется лобным рогом; лобные и все окологлазничные кости бугристы на внешней поверхности и так массивны, что при рассматривании черепа сверху совсем не видно глазных впадин: они прикрыты "нахмуренным" лбом и задними краями глазниц. При длине всего черепа около 40 см с небольшим толщина теменных костей не менее 7 см (очевидно, еще больше была длина нерва теменного глаза). Утолщение теменных, скуловой, всех околглазничттых o костей и костей височной области сужает височную впадину, вмещавшую височный мускул (особенно в ее нижней части"), в относительно узкую щель.
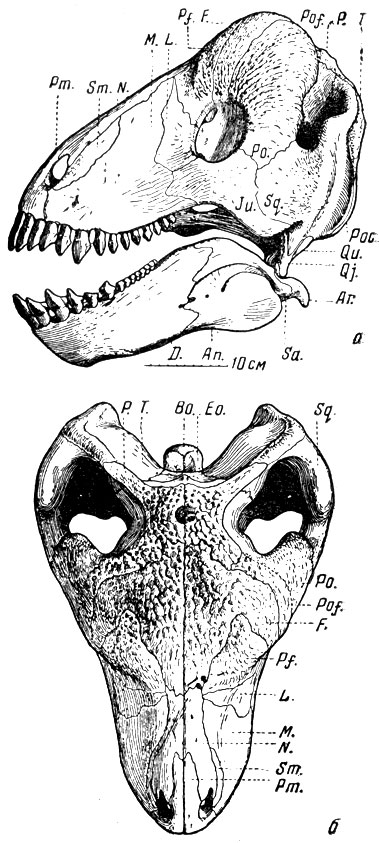
Рис. 80. Улемозавр (Ulemosaurus svijagensis Rjabinin). а - череп, вид сбоку; б - вид сверху. Верхняя пермь. Татарская АССР. (И. А. Ефремов, 1940)
Затылок пропорционально выше, чем у титанозухий, поперечные отростки крыловидных костей развиты слабее, не доходят до ветвей нижней челюсти и не ограничивают ее движений; челюстной сустав допускает некоторую свободу боковых движений, что обычно характерно для растительноядных (и всеядных) позвоночных. Скуловая дуга, морда и нижняя челюсть короче, выше и массивнее, чем у хищных дейноцефалов. Небосходно с нёбом титанозухий, но улемозавр лишен бугров с зубами на нёбных костях и без зубов на крыловидных. Столбик уха, "стремечко", как и у титанофонеуса, имеет вид распорки и в основании черепа между квадратной костью, сочлененной с нижней челюстью, и мозговой коробкой. Столбик закрывает своим расширенным концом овальное окно, ведущее в лабиринт внутреннего уха; столбик еще массивнее, чем у титанофонеуса, и на первый взгляда еще менее может быть заподозрен в способности проводить звук; в общем, восприятие звука здесь столь же неясно, как и у титанофонеуса. У некоторых современных пресмыкающихся, по своему образу жизни "прижатых" к земле (например, змей и некоторых ящериц), звуковые колебания, по-видимому, передаются внутреннему уху через кости черепа; однако" такой образ жизни трудно предположить для титанофонеуса, а особенно для улемозавра, голова которого, судя по строению плечевого пояса, передних ног и позвоночного столба, была обычно довольно высоко поднята над землей (рис. 82).
Отогнутый вниз затылочный сочленовный бугор черепа, нависший над ним затылок и форма скошенных на переднем и заднем концах шейных позвонко в указывают на наклонное положение головы улемозавра (концом морды обычно вниз).
Зубы улемозавра, около 15 на каждой стороне вверху и внизу, необычного для пресмыкающихся вида, но типичного для растительноядных тапиноцефалов. Самые интересные - передние, "резцы" (рис. 81). Они много крупнее остальных, в связи с этим передние концы межчелюстных и нижнечелюстных костей очень массивны. Далее назад зубы постепенно уменьшаются (рис. 80, а); исключение составляют снабженные режущими краями клыки, хотя они почти утратили клыкообразность. В основании коронки резцов верхних и нижних, на их язычной стороне, очень сильно развит выступ, так называемая "пятка"; в ее углубление входит вершина резца - антагониста противоположной челюсти. Эмаль зубов по краям коронок острая, зубчатая. При смыкании челюстей вследствие частичного вклинивания нижних зубов в промежутки между верхними (и наоборот) резцы одновременно перерезают, раздавливают и перетирают пищу (рис. 81, в), чему и должен был соответствовать характер работы челюстной мускулатуры. Таким образом, эти зубы - резцы по местоположению - одновременно выполняют и работу коренных зубов, будучи хорошо приспособлены для измельчения пищи, вероятно, каких-то мясистых и сочных растений. Послеклыковые зубы мелкие с отрыми режущими передними и задними краями (как у относительно слабого клыка) и скорее всего удобны для перекусывания растений.
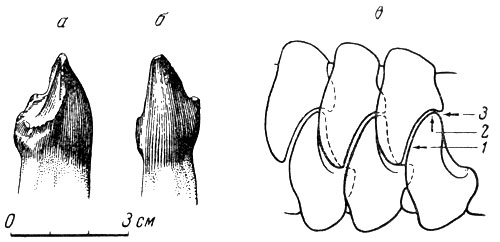
Рис. 81. Улемозавр (Ulemosaurus svijagensis Rjabinin). а - нижний третий правый резец улемозавра, вид сбоку; б - спереди; в - схема режущей, перетирающей и раздавливающей работы резцов улемозавра. Верхняя пермь, Татарская АССР (M. А. Ефремов, 1940)
Для тапицефалов (рис. 82) - растительноядных пресмыкающихся, к которым относятся улемозавры, характерны очень oсильные высокие передние ноги, массивный передний пояс (лопатка, коракоиды, ключицы, межключица). Спина своеобразно поката назад, на лапах последние фаланги в виде мелких копытец; сильные передние конечности возможно, служили и для добывания пищи и, видимо, (вообще выполняли наибольшую работу, задние относительно слабые. Строение конечностей, плечевого пояса и таза указывает на возможность (высоко приподнятого над землей положения животного, подобно млекопитающим (как это, впрочем, иногда наблюдается и у передвигающегося по суше крокодила). Для улемозавра такое приподнятое над землей положение было скорее всего обычно. Вряд ли были особенно проворны улемозавр и его ближайшие родичи, другие тапиноцефалы, более древние - дейтерозавры из верхнепермских отложений Южного Приуралья и тем более - поздние и громоздкие южноафриканские мосхопс, мормозавр и др. Но все тапиноцефалы были, очевидно, хорошо приспособлены к (растительной пище и, вероятно, поэтому распространились так широко. Это первые, наиболее древние известные крупные растительноядные пресмыкающиеся, вообще первые крупные растительноядные наземные позвоночные, притом "копытные", но пошедшие в приспособлении к растительной пище несколько необычным путем. Для измельчения растений у тапиноцефалов служили резцы, а не послеклыковые щечные зубы, как это было позднее, в мезозое у динозавров, в кайнозое - у млекопитающих. Любопытно, что задние щечные зубы улемозавра сохранили режущий тип зубов своих предков, пожалуй, даже усилил® его сомкнутым расположением зубов.
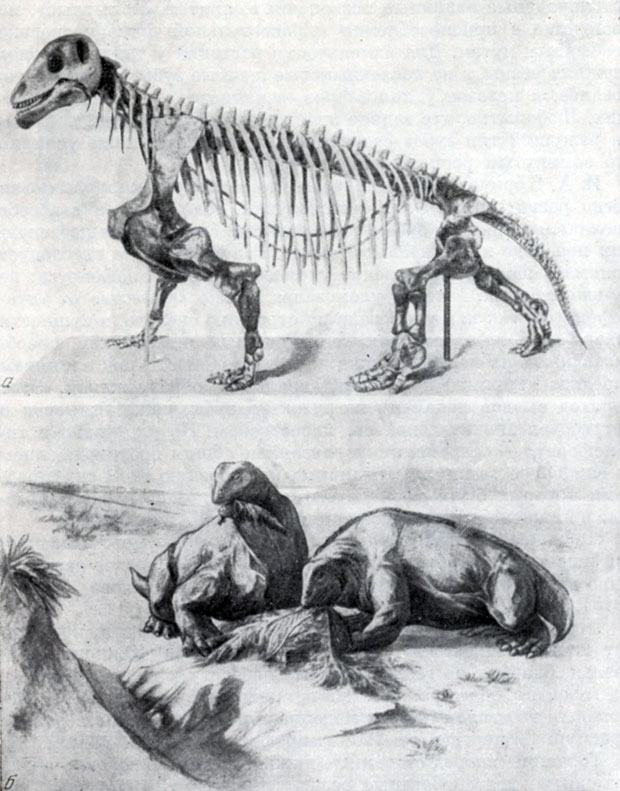
Рис. 82. Мосхопс (Moschops capensis Broom), близкий к улемозавру растительноядный дейноцефал из верхнепермских отложений формации Карроо Южной Африки а - скелет; б - реконструкция внешнего вида (W. Gregory, 1926)
И. А. Ефремов (1940) показал, что резцы улемозавров могли легко развиться из зубов титанозухий, если представить себе увеличение "пятки" зуба на его язычной стороне при постепенном переходе к растительной пище. Разумеется, это не следует понимать как происхождение улемозавра от титанофонеуса: не только зубы, но и череп улемозавра можно "вывести" от титанофонеуса, так как улемозавр по строению скелета, в сущности говоря, есть лишь видоизменение хищных титанозухий, преобразованных на первый взгляд до неузнаваемости под влиянием перехода хищника к растительноядному образу жизни. Этот переход вызвал перемену нагрузок на зубы, челюсти, череп и потребовал значительной его перестройки. Но улемозавр и титанофонеус - современники, связанные общим происхождением от еще более древних и архаичных по строению терапсид, предками которых были сфенакодонтные пеликозавры, разнообразные в нижнепермских отложениях Северной Америки.
Не менее замечательно и другое загадочное пресмыкающееся ишеевской фауны - венюковия (рис. 83). Родовое название было дано В. П. Амалицким в честь П. Н. Венюкова (1858-1915), профессора Петроградского горного института.
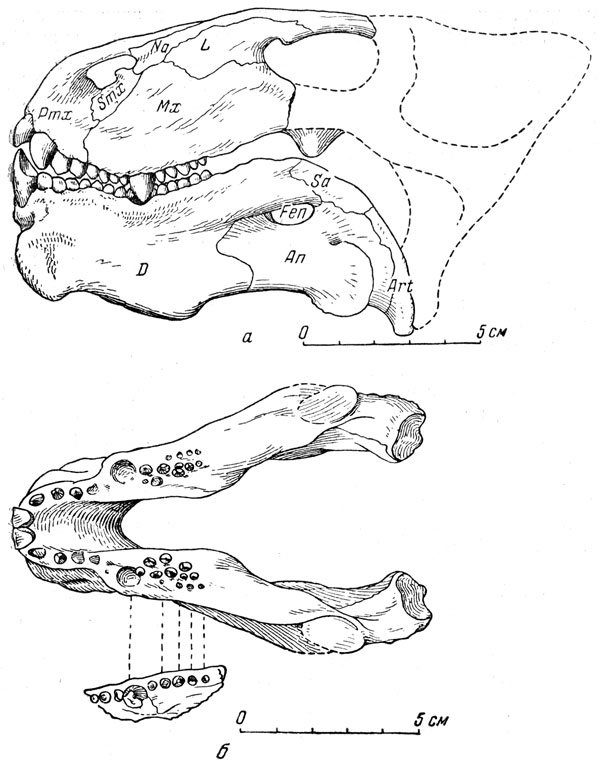
Рис. 83. Вешоковпя (Venjukovia invisa Efremov). а - череттГгвид сбоку (И. А. Ефремов, 1940); б - нижняя челюсть с куском левой верхней челюстп, вид сверху (Ватсон, 1948). Задние концы правой и левой ветвей нижней челюсти должны быть раздвинуты тире, судя по другим образцам (сжаты деформацией после захоронения). Верхняя пермь, Татарская АССР
Первая находка, четыре обломка нижней челюсти, происходит из верхнепермских медистых песчаников Южного Приуралья, близких по геологическому возрасту к пескам Ишеева, но, вероятно, более ранних. Эти остатки были собраны П. Н. Венюковым в отвалах старых медеплавильных рудников в районе Оренбурга и переданы в 1908 г. профессору В. П. Амалицкому.
Торчащие вперед сточенные нижние резцы венюковии очень похожи на долотообразные резцы грызунов. Это сходство, несомненно, и было причиной ошибки В. П. Амалицкого (1922),. который отнес животное к млекопитающим. Истинная природа венюковии была выяснена И. А. Ефремовым лишь после раскопок около Ишеева в 1935 г., когда стали попадаться такие же обломки нижних челюстей, а затем нашелся и череп.
Венюковия из описываемой фауны поражает массивностью конца морды, особенно нижней челюсти (рис. 83). Межчелюстные кости необычайно развиты и утолщены; на нёбе они доходят до нёбных, а снаружи черепа почти до уровня глазниц. Сильно утолщены и верхнечелюстные кости; а высота нижней челюсти та же, что самого черепа впереди глазниц. Особенно развита передняя часть нижней челюсти; ее передний край oсильно утолщен, внизу справа и слева имеются массивные выступы, спереди придающие симфизу вид раздвоенного подбо родка, опускающиеся значительно ниже симфиза. Пожалуй, самое замечательное у этого пресмыкающегося - его зубы, хотя сами по себе они имеют простое строение.
Самые крупные - резцы верхней челюсти, второй и третий, низкие, толстые; слегка вогнутые площадки стирания на корон ке сходятся под углом на вершине зуба. Далее назад следует после нескольких мелких конических зубов крупный, низкий, толстый клык со слабо стирающейся коронкой. Он входит при смыкании рта своей вершиной в особую ямку в нижней челюсти снаружи от нижних зубов (рис. 83). Из нескольких мелких конических зубов, следующих в верхней челюсти за клыком, самые последние стоят попарно рядом или почти рядом, а не друг за другом, как обычно. Еще отчетливее это необычное расположение наблюдается в заднем отделе нижнего зубного ряда (рис. 83, б) . На переднем конце нижней челюсти - два крупных долотообразных резца. И если передние зубы верхней и нижней челюсти сходны с резцами грызуна, то задние, расположенные в два ряда, по своему назначению, очевидно, соответствуют бугорчатым коренным зубам млекопитающего. Таким образом, венюковия своими зубами напоминает грызунов с их долотообразными резцами и с перетирающими растительную пищу коренными. Однако это сходство с грызунами относительное.
У венюковии не сохранилось заднего отдела черепа, но, судя по уцелевшей глазничной части, задняя половина черепа вряд ли была массивной. Во всяком случае лицевая часть сравнительно с глазничной очень мощная; особенно же непомерно массивна передняя половина нижней челюсти (рис. 83, а), а ее задняя половина и суставной отдел относительно слабые; поел еклыковые зубы мелкие. Если венюковия и обладала передними грызущими зубами, то ее послеклыковые зубы не годятся для перетирания твердой растительной пищи, зерен и т. п., и животное вряд ли питалось, как грызуны. Поэтому И. А. Ефремов (1940) предположил, что само питательное вещество было мягким, но извлекалось из каких-то твердых и крупных оболочек, которые не могли быть непосредственно раздроблены во рту, например раковин, и что венюковия могла питаться крупными растениями с твердой оболочкой вроде современных саговых пальм или кактусов. У некоторых современных грызунов имеются подбородочные выступы на нижней челюсти, как у венюковии. Эти выступы и небольшой гребень на черепе впереди глазницы побудили И. А. Ефремова предположить у венюковии большую силу челюстной мускулатуры.
Мы не знаем современных или ископаемых пресмыкающихся в точности с таким строением нижней челюсти, как у венюковии. Среди современных млекопитающих массивная лицевая часть черепа и нижняя челюсть имеются у моржа, питаю щегося главным образом двустворчатыми моллюсками. Простые столбообразные коренные зубы моржа относительно совсем не велики и служат для дробления раковин моллюсков; тяжеловесная нижняя челюсть моржа дробит раковину о верхние зубы, как молот о наковальню; глотает же морж только мягкое тело моллюска. Быть может, непонятная при небольших относительно слабых зубах тяжеловесность морды и нижней челюсти венюковии - такое же приспособление для дробления раковин, как и у моржа?
Интересны костные ямки на нижней челюсти для вершин верхних клыков венюковии. При смыкании челюстей вершина клыка входила в соответствующую ямку противолежащей челюсти, но разумеется, не упиралась непосредственно в кость: последняя должна была быть покрыта не только надкостницей, но и десной в целом. Кроме того, зубной ряд нижней челюсти при смыкании рта ложился внутрь от верхних зубов и упирался в массивные небные кости, а не в зубы верхней челюсти. Аналогичное явление имело место для нижних клыков титанофонеуса, долиозавра и сиодона, а также и для послеклыковых зубов долиозавра - на боковой части его небных костей, на границе с верхнечелюстными, имеются также ямки для нижнечелюстных щечных зубов (см. выше). Все это указывает на сильное ороговение покровного эпителия полости рта у упомянутых выше титанозухий, а у венюковии, возможно, с образованием прочного рогового чехла. У дицинодонтов, происшедших от венюковиеобразных предков и утративших в ходе исторического развития все зубы, кроме клыков, по-видимому, был роговой клюв, как у черепах. У дицинодонтов на это указывает и почти черепашья форма конца морды.
Сходство черепа венюковии с черепом титанозухий побудило И. А. Ефремова считать венюковии своеобразными дейноцефалами, сходными с дицинодонтами и другими аномодонтами (греческое - неправильно-, необычнозубые). Позднее Д. М. Ватсон (1942, 1948) указал на то, что сходство с хищными титано-зухиями и одновременно с аномодонтами объясняется прямым родством, промежуточным положением вепюковии между теми и другими. Во всяком случае венюковия подтверждает происхождение дициподонтов от титанозухий.
Крупные размеры и более высокое, по мнению И. А. Ефремова, стратиграфическое положение венюковии ишеевской фауны сравнительно с первыми находками в Приуралье побудило выделить ее в новый вид V. invisa Efremov (И. А. Ефремов, 1940). Отсутствие в сборах позвонков и конечностей, которые можно было бы с достоверностью приписать венюковии, делает это животное одной из наиболее интересных загадок пермской эпохи.
Другие пресмыкающиеся ишеевской фауны представлены пока скудным материалом. Сюда относятся остатки неизвестных мелких пресмыкающихся из наиболее древнего отряда котилозавров. Кроме того, найдены остатки тероцефалов (греческое - звероголовые) - хищных пресмыкающихся, относящихся к тому же подклассу синапсид. Некоторые мелкие поздние тероцефалы (триас Южной Африки) более других близки к вероятным предкам первых млекопитающих.
Лантанозух (греческое - скрытое, загадочное животное) - поистине загадочное животное ишеевской фауны, которое совмещает в себе некоторые черты строения древних земноводных и одновременно пресмыкающихся (рис. 84).
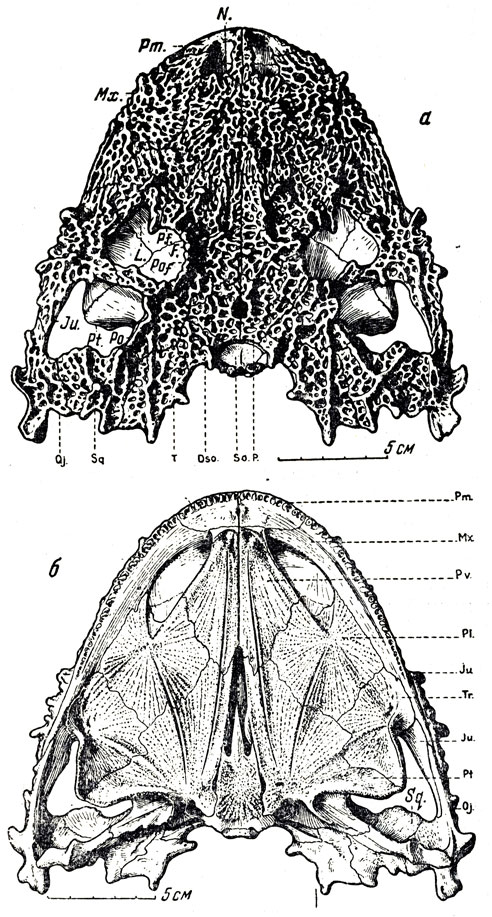
Рис. 84. Лантанозух (Lanthanosuchys qualeni Efremov). а - череп, вид сверху; б - снизу. Верхняя пермь, Татарская АССР. (И. А. Ефремов, 1946)
У лантанозуха широкий, плоский череп с несколько выпуклыми, направленными вверх и слегка в стороны глазницами; глаза при жизни животного, очевидно, довольно сильно выдавались, как и у многих современных земноводных; верхняя сторона черепа покрыта скульптурой из сети перекладин, повышавших прочность плоского черепа; внешне она напоминает ямчато-бугорчатую скульптуру черепа многих крокодилов. Самое любопытное - довольно большие височные впадины позади глазниц; эти впадины свойственны прогрессивным пресмыкающимся, но еще отсутствуют у типичных котилозавров, первых примитивных пресмыкающихся. В то же время нёбо лантанозуха в основном того же характера, что и у древнейших зем новодных: сошниковые, нёбные и крыловидные кости широкие, большие и образуют сплошную костную поверхность. Более того, все нёбо покрыто рядами мельчайших бугорков, напоминающих "шагрень" (кожные зубы) некоторых древних рыб; эти бугорки расположены у лантанояуха рядами, лучисто расходящимися от центров окостенения нёбных костей; нёбо лантанозуха этим напоминает даже рыб. Но крыловидные кости имеют отчетливые поперечные отростки, направленные вниз, как у многих пресмыкающихся. Верхнечелюстные и нпжнече-люстные зубы лантанозуха конические, простые, как вообще у лабиринтодонтов, но стенка зубов без характерной сложной лабиринтной складчатости, которая привела к самому названию "лабиринтодонты". Таким образом, челюстные зубы лантанозуха построены так же, как и у пресмыкающихся. Можно было бы составить длинный перечень особенностей строения лантанозуха, характерных для земноводных, и такой же для примитивных пресмыкающихся. Все это ставит лантанозуха как бы в промежуточное положение между теми и другими, но тем не менее отнюдь не в переходное от земноводных к пресмыкаюпцшся. Древнейшие пресмыкающиеся обнаружены в гораздо более древпих отложениях верхнекаменноугольного возраста и во всяком случае не моложе низов нижнепермских; к тому же первые пресмыкающиеся меньше лантанозуха. Он мог бы считаться лишь потомком действительно переходных от земноводных к пресмыкающимся животных, дожившим при благоприятных условиях до верхне-пермского времени. Вместе с несколько более поздней верхнелермской котлассией с Северной Двины, добытой раскопками В. П. Амалицкого (см. ниже рис. 100), и североамериканской нижнепермской сеймурией лантанозух выделен И. А. Ефремовым в особый подкласс "батрахозавров". Эта группа, одно время разнообразная по своим представителям и занимавшая промежуточное положение между земноводными и пресмыкающимися, вероятно, возникла еще в нижнекаменноугольное время при развитии из земноводных первых пресмыкающихся. Видимо, и образ жизни батрахозавров, возможно, и их размножение были разнообразны и представляли примеры перехода от характерного для земноводных откладывания икры в воду к откладыванию яиц с плотными оболочками на суше. Однако здесь мы вступаем пока в область догадок и неполноты геологической летописи, столь досадной, но хорошо знакомой каждому палеонтологу...
Лантанозух, вероятно, питался мелкой рыбой и другой мелкой добычей.
При знакомстве с земноводными ишеевской фауны, лабиринтодонтами, бросается в глаза различный тип строения их черепа и, особенно, его лицевой части. Широкомордые (энозух), с удлиненным рылом (трифовух), - все это хищные животные с многочисленными острыми, коническими, большей частью мелкими зубами. По-видимому, они охотились в основном за мелкой рыбой, водными насекомыми и их личинками, червями, прибрежными наземными моллюсками и другой по преимуществу мелкой добычей верхнепермских пресных вод и побережий. Зубы лабиринтодонтов, несмотря на различие в деталях, в общем все же довольно однообразны, хватающие и пронзающие добычу; впрочем, мелкую добычу лабиринтодонты с частым расположением зубов, возможно, могли и перекусить. Но добыча лабиринтодонтов при остроконечных зубах, вероятно, должна была быть лишена крепких твердых покровов вроде панциря крупных раков, толстой раковины моллюсков или прочной чешуи некоторых рыб. Тем резче при общем однообразии зубного "вооружения" этих земноводных бросается в глаза различный тип строения морды; это, вероятно, указывает на разные повадки хищника и на различный выбор добычи, быть может, на довольно разнообразный "стол" лабиринтодонтов, хотя низкий уровень развития головного мозга земноводных и не содействовал разборчивости в пище. Многочисленность земноводных и хищных пресмыкающихся, естественно, была возможна при общем изобилии пищи в виде рыбы, беспозвоночных, кормившего их планктона, водной и наземной растительности Однако о многом приходится пока лишь догадываться.
Наиболее понятны остатки некоторых рыб, найденных в Каменном Овраге вместе с земноводными и пресмыкающимися. Прежде всего могут быть упомянуты платисомусы, видимо, довольно обычные в верхнепермских пресных водах с очень высоким телом (рис. 85), с характерными вертикально вытянутыми, покрытыми тонкой эмалью чешуями. Весьма вероятны, кроме платисомусов, небольшие амблжгтерусы, широко распространенные в пермское время, а также другие костные рыбы. Однако многочисленные остатки рыб из Каменного Оврага очень разрознены, трудны для определения и не описаны.
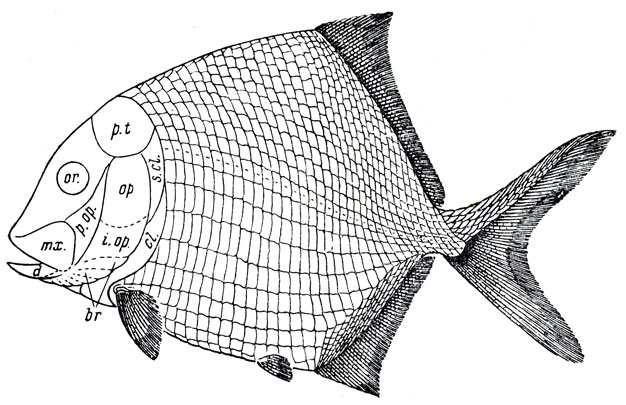
Рис. 85. Платисомус (Platysomus striatus Agassiz) (Traquair, 1879)
Интересны части обызвествленных, а поэтому сохранившихся черепов и нижней челюсти хрящевых (акуловых) рыб; их плавниковые шипы (ихтиодорулиты), а также минерализованный помет в виде "копролитов" с характерным для акуловых рыб отпечатком спирального клапана кишечника. Эти копролиты, сильно известковистые от непереваренных костных остатков, самых различных размеров и несомненно принадлежат мелким и крупным акуловым рыбам; многие содержат хорошо сохранившуюся чешую платисомусов и, вероятно, других костных рыб. Во всяком случае акуловые рыбы были здесь многочисленны и, возможно, довольно разнообразны. Это обстоятельство, вероятно, говорит о дельтовом характере глинисто-песчанистой толщи, в которой залегают остатки пресмыкающихся, земноводных и рыб ишеевской фауны, скорее всего о непосредственной близости моря и его приливной полосы.
Несмотря на все разнообразие верхнепермских позвоночных, собранных в Каменном Овраге, мы еще далеки от совершенного знания этой фауны. Лишь некоторые из найденных животных представлены достаточно полным костным материалом (черепа и скелеты); много остатков принадлежат каким-то неизвестным позвоночным. Кроме того, в этом захоронении в итоге сортирующей работы воды, отлагавшей костеносные пески, преобладают остатки сравнительно крупных животных, притом только позвоночных. Между тем здесь несомненно жили двустворчатые пресноводные моллюски, например типа мелких антракозид, широко распространенных в то время, и вообще разнообразные пресноводные или солоноватоводные беспозвоночные. В это время уже было известно много разных насекомых, найденных в приблизительно одновозрастных отложениях территории Советского Союза А. В. Мартыновым и др. (рис. 86). Наконец, из пермских же континентальных отложений Среднего Урала известны разнообразные растения (рис. 87). К ним очень близка должна была быть растительность, которая давала ишеевской фауне пищу и убежище от солнца или непогоды. Но условия накопления осадков в местонахождении Каменного Оврага явно не могли быть благоприятны для захоронения хрупких и мелких остатков антракозид, для образования отпечатков крыльев насекомых, листьев растений и т. п. Сильная река несла целые или полуразложившиеся трупы крупных пресмыкающихся вроде титанофонеуса или улемозавра; вода окатывала и разрушала, местами превращая почти в песок и костяную муку разрозненные кости и их обломки. В такой обстановке многое не могло уцелеть и захорониться. Изучение местонахождения в Каменном Овраге при раскопках, выполненное И. А. Ефремовым (1940, 1950), выяснило много интересного. Раскопки показали следующий разрез местонахождения (сверху вниз):
1. Лесная, почва (суглинок, щебенка, залегающие до 1,2 м
2. Известняк светло-серый, ноздреватый, сильно до 2,0
3. Красная сильно известковистая глина; до 2,0
4. Прослой рыхлого мергелистого песчаника; до 0,4
5. Костеносные пески, серые, красноватые с косой слоистостью не менее 30
Сильный приток воды в нижних частях костеносного пласта не дал возможности пройти его в глубину и установить полную вертикальную мощность пласта. Но И. А. Ефремов сделал во бремя раскопок следующие наблюдения над строением костеносного слоя и распределением в нем скелетов и других органических остатков.

Рис. 86. Ископаемые насекомые пермского периода а - соянорафидия (Sojanoraphidia rossica О. Martynova). Верхнепермские отложения р. Сояны Архангельской области. Ископаемая верблюдка, самка с яйцекладом. Голова - как у современных насекомых, но 'переднегрудь' еще не вытянута (О. М. Мартынова, 1962)
В верховье оврага, по его правому склону, выходят косослоистые красноватые или бурые пески, наиболее богатые костными остатками. Ниже по оврагу у его левого склона залегают преимущественно серые пески, содержащие лишь разрозненные и редкие обломки костей.

Рис. 86. Ископаемые насекомые пермского периода б - палеутиграмма (Paleutigramma tenuicornis Martynov) из вымершего отряда 'предпрямокрылых'. Артинский ярус среднего Урала. Увеличено (А. В. Мартынов, 1930)
Все скелеты и все наиболее ценные остатки залегали в красноватых песках и были тем лучшей сохранности, чем дальше находились от серых песков. Полный скелет титанофонеуса сохранил естественное взаимное расположение всех костей, хотя и был поврежден двумя параллельными сбросами, измельчившими часть позвоночника и плечевого пояса. Скелет долиозавра залегал в самой верхней части красных песков; скелет лежал позади черепа (по течению потока) наклонно вниз с беспорядочным нагромождением костей, а череп с нижней челюстью был повернут нижней стороной вверх; нижняя челюсть и громадные верхние клыки были срезаны по контакту с залегающими выше красными глинами.

Рис. 86. Ископаемые насекомые пермского периода в - палингениопсис (Palmgeniopsis praecox Martynov). Ископаемая поденка, отпечаток заднего крыла. Верхнепермсние отложения р. Сояны Архангельской области. Увеличено (А. В. Мартынов, 1931)
Остатки улемозавров добыты экспедицией Б. А. Штылько (1930-1931) на самом краю красноватых песков на границе с серыми. Здесь от скелетов сохранились наиболее тяжелые части, при этом сильно смещенные относительно друг друга. Таким образом, все найденные скелеты, начиная с первого погибшего, обнаруженного Носовым, залегали в полосе красноватых песков шириной в 8-10 м, простирающейся с запада на восток. И. А. Ефремов предполагает, что скелеты, погибших животных задерживались на песчаных отмелях вначале своей самой тяжелой частью - черепом, а потом уже поворачивались или разносились течением. Следует, (впрочем, иметь в виду, что туша каждого животного в целом (при захоронении трупов целиком) тяжелее головы; центр тяжести туши, например, улемозавра приходился позади плечевого пояса (или на его уровне), поэтому предположения И. А. Ефремова наиболее верны применительно к захоронению скелетов (именно скелетов) крупных титанозухий с их относительно тяжелым черепом и стройным и легким остальным скелетом.
В общем на основе имеющихся данных И. А. Ефремов предполагает следующую картину образования Ишеевского местонахождения.

Рис. 87. Верхнепермские растения Западного Приуралья а - пекоптерис (Pecopteris concinna Kutorga), папоротник
Захоронение трупов погибших животных, их скелетов, а также разрозненных костных остатков происходило в водном потоке речного типа, вероятно в низовьях реки, может быть в ее дельте. На это указывает отсутствие крупного осадочного материала тииа галечников, хорошая отсортировакность песков, их мелкозернистость (а также и наличие хороших остатков акуловых рыб, как известно типичных морских хищников, редко поджимающихся в реки выше их устий. - Ю. О.). Серые пески отлагались, видимо, в более глубокой части русла, с более быстрым течением, которое сносило остатки животных дальше; глубокая вода препятствовала осаждению безводной окиси железа. Это осаждение, окрасившее в красноватые и бурые тона костеносные красные пески, могло, наоборот, происходить с легкостью в мелких рукавах или тихих заводях, где заносимые течением остатки животных задерживались при спаде воды. Обогащение органическими веществами содействовало образованию здесь песков бурых тонов.

Рис. 87. Верхнепермские растения Западного Приуралья б - одонтоптерис ('неуроптерис'),(Odontopteris rossicaZalessky), папоротникообразное семенное растение. Уменьшено (М. Д. Залесский, 1927)
Вероятно, окончательное захоронение целых скелетов или туш происходило в межень, при быстром спаде воды; на это указывает и положение крупного скелета титанофонеуса на верхней наклонной плоскости серых песков. Около крупных костей часто встречается мелкая костяная брекчия, главным образом из остатков рыб.
Кости Ишеевского местонахождения минерализованы слабо, крайне хрупки и различны по своей сохранности. Большинство из них светлые, желтовато-серого цвета, вероятно, выбеленные солнцем до захоронения в пески, в которых приобрели местами красноватую или бурую окраску самих песков. (Хрупкость костей потребовала при препаровке длительного и повторного пропитывания их лаком после предварительной просушки). Все эти особенности местонахождения дейноцефаловой фауны в Каменном Овраге около села Ишеева привели И. А. Ефремова к следующим предположениям.
Прежде всего, местонахождение образовалось вблизи местообитания улемозавров и титанозухов. Те и другие, по-видимому, вели "приречное" существование, улемозавры вследствие обилия растительной пищи, титанозухи - богатого улова в воде и на суше.
Трупы улемозавров и титанозухов заносились в мелкие рукава и на отмели водой, но предварительное разложение в воде (до окончательного захоронения в пески) не могло быть длительным. На это указывает полнота некоторых скелетов. Следовательно, они не были принесены издалека. Условия захоронения ишеевских дейноцефалов, по мнению И. А. Ефремова, сходны с современными условиями захоронения крупных аллигаторов Северной Америки, гибнущих при огромных наводнениях в низовьях Миссисипи и после быстрого спада воды остающихся в мелких рукавах и протоках.
Большое число мелких, часто окатанных в почти круглую гальку, фрагментов костей в красных, а также и серых песках указывает, что эти пресмыкающиеся были распространены и далеко вверх по течению этой реки. Большое количество костной брекчии из остатков рыб говорит об обилии органической жизни IB реке. Кости других пресмыкающихся - тероцефа-лов, котилозавров и венюковий - скудны. Это указывает на то. что их остатки попадали сюда редко и в небольшом количестве возможно, вследствие обитания самих животных в более удаленных от реки местах.
Скелеты стегоцефалов, проводивших большую часть своей жизни непосредственно в воде, подвергались после гибели животных мацерации в воде и непосредственному захоронению без выветривания на воздухе. Вероятно, поэтому их кости в среднем лучшей сохранности, чем пресмыкающихся. Более полные остатки стегоцефалов следует искать выше по течению этой верхнепермской реки.
Акуловые рыбы, как известно, редко поднимаются высоко вверх по рекам; во всяком случае, очевидно, они жили где-то вблизи или здесь же, так как их хрящевые, но сильно обызвест-вленные черепа и челюсти хорошо сохранились, значит, подвергались быстрому захоронению. Остатки самих акул и множество их копролитов свидетельствуют, как указывалось, скорее всего о том, что местонахождение образовалось хотя и в водном потоке речного типа, но где-то в низовьях реки, может быть, в ее дельте. Об этом говорит и отсутствие в песках крупного осадочного материала типа галечников, хорошая отсортированность песков, их мелкозернистость.
Несмотря на всю неполноту наших сведений об ишеевской фауне, ее изучение внесло в науку много нового. Следует отметить промежуточное по геологическому возрасту и строению скелета, вероятно, и по "этапу эволюционного развития" положение ишеевских титанозухий и тапиноцефалов между несколько более древними их представителями с Южного Приуралья (еще более архаичными и более близкими к нижнепермским пресмыкающимся), с одной стороны, и более поздними и массивными южноафриканскими - с другой. Впрочем, разбор этого вопроса выходит за пределы настоящего очерка. Задача автора - ознакомить с некоторыми успехами палеонтологии в изучении позвоночных из верхнепермских континенталышх отложений СССР за последние годы, в частности, с титанозухи-ями и тапиноцефалами. Пожалуй, наиболее любопытная черта этих рептилий - это то, что они дали для времени, отстоящего от нас примерно на двести миллионов лет по современной шкале геологического летоисчисления, экологические типы растительноядного "копытного" (улемозавр), крупного хищника (долиозавр), почти крокодила (титанофонеус), небольшого всеядного хищника (сиодон), не то грызуна, не то моллюскоеда (венюковия) и вообще заняли самые различные "ниши" соответственно условиям жизни во всем их вечном разнообразии...
Общее сходство рассмотренных пресмыкающихся с южноафриканскими из зоны Tapinocephalus очевидно. Но все наиболее легко сопоставимые рептилии волжской фауны (а тем более другой, близкой, по, видимо, более ранней из медистых песчаников Приуралья, описанной И. А. Ефремовым, 1954) значительно менее специализированы, чем многие южноафриканские, массивные, с сильным пахиостозом. Это и было неоднократно (1940-1954) отмечено И. А. Ефремовым. Интересно, что некоторые примитивные черты, унаследованные еще от сфенакодонтных пеликозавров, встречаются и у южноафриканских форм титанозухов и тапиноцефалов вперемежку с чертами крайней специализации.
Рассмотрим вкратце другую позднепермскую фауну, Северо-Двинскую, соответствующую татарскому ярусу морских фаций, добытую в конце 90-х и в начале 900-х годов на правом берегу р. Малой Северной Двины вблизи г. Котласа раскопками В. П. Амалицкого, сыгравшими большую роль в успехах палеонтологии позвоночных не только у нас, но и за рубежом (рис. 88).

Рис. 88. Правый берег р. Малой Северной Двины у дер. Ефимовской около г. Котласа, место основных раскопок В. П. Амалицкого
В 90-х годах прошлого века В. П. Амалицкий (1860-1917) в отложениях неопределенного возраста, считавшихся до него "немыми", лишенными органических остатков, обнаружил остатки пластинчатожаберных моллюсков (антракозид) и примитивных "глоссоптерисообразных" папоротников верхнепермского возраста южного, "гондванского" типа. (Гондвана - название предполагаемого древнего материка, объединявшего Индостан, Африку, Южную Америку и Австралию, по живущему в центре Индостана племени гон дав). На этом основании им было высказано предположение о предстоящем открытии на пашей территории пермских позвоночных "южноафриканского" типа. Смелое предсказание, вначале вызвавшее недоверие, вскоре подтвердилось; В. П. Амалицким были обнаружены местонахождения, в том числе костеносные линзы на Северной Двине и Сухоне, приуроченные к речным руслам верхнепермского времени, по-видимому, в приустьевых участках этих рек.
Линзы залегают в красноватых и пестроцветных глинах и мергелях и содержат местами огромные скопления массивных конкреций прочного песчаника, цементированного главным образом карбонатами; внутри конкреций оказались отдельные кости, черепа и целые скелеты пресмыкающихся и земноводных прекрасной сохранности. Постановка огромных раскопок, доставка сборов первоначально в Варшаву, где в то время работал Амалицкий, их препаровка и первичная монтировка, изготовление большой серии огромных витрин, эвакуация всех материалов во время первой мировой войны в глубокий тыл, реэвакуация в Петроград и другие организационные мероприятия потребовали от Амалицкого опромной и напряженной работы, сломившей его здоровье; он скончался в 1917 г. и успел написать лишь предварительные сообщения об этой фауне, частично опубликованные уже после его смерти. Изучение завершается лишь в настоящее время, но в сущности и его пока нельзя будет считать окончательным - берега Северной Двины и ее притоков содержат, несомненно, еще много замечательных материалов, частично открытых совсем недавно, частично ожидающих своего открытия; да и две песчаные линзы, откуда добыт рассматриваемый ниже материал, раскопаны далеко не полностью...
Большая часть остатков в обширной коллекции этой фауны принадлежит парейазаврам из подкласса анапсид (типичные анапсиды лишены височного окна); они явно преобладают, и в этом отношении вся фауна с известным правом может называться "парейазавровой". Северодвинские парейазавры позднее были выделены А. П. Гартман-Вейнберг (1934) в особый род, получивший название скутозавра по наличию характерных костных щитков, покрывавших их туловище (скутум - щит). Это крупные животные, величиной с небольшого быка, с очень широким черепом (рис. 89-91), частью за счет разрастания квадратноскуловых костей, и с маленькой мозговой коробкой, небольшим, но отчетливым отверстием для теменного глаза; верхняя поверхность черепа сильно бугриста. Очень характерны верхние и нижние краевые зубы, острые, лопаточковидные зазубренные и, по-видимому, хорошо приспособленные для обкусывания каких-то растений, вероятно сочных, но не для жевания; кроме того, большое число мелких зубов расположено на нёбных и крыловидных костях, что придает нёбу характер "терки". В остальном скелете бросается в глаза массивность позвоночного столба с типичным для котилозавров - древних аыапсидных рептилий - очень аильным развитием нейральных дуг; далее массивные пояса конечностей и сами конечности с почти горизонтальной постановкой плеча и бедра; лапы снабжены приплюснутыми сверху вниз относительно короткими пальцами, с небольшими конечными фалангами, похожими на копытца. Хвост короткий. Раскопками на Северной Двине было добыто около десятка полных и почти полных скелетов, около тридцати отдельных черепов и тысячи разрозненных костей и обломков остального скелета. В настоящее время это "стадо", представляющее мировую научную ценность, украшает собою Музей Палеонтологического института Академии наук СССР, а вся "Северодвинская галерея" в целом представляет одну из главных достопримечательностей этого музея.
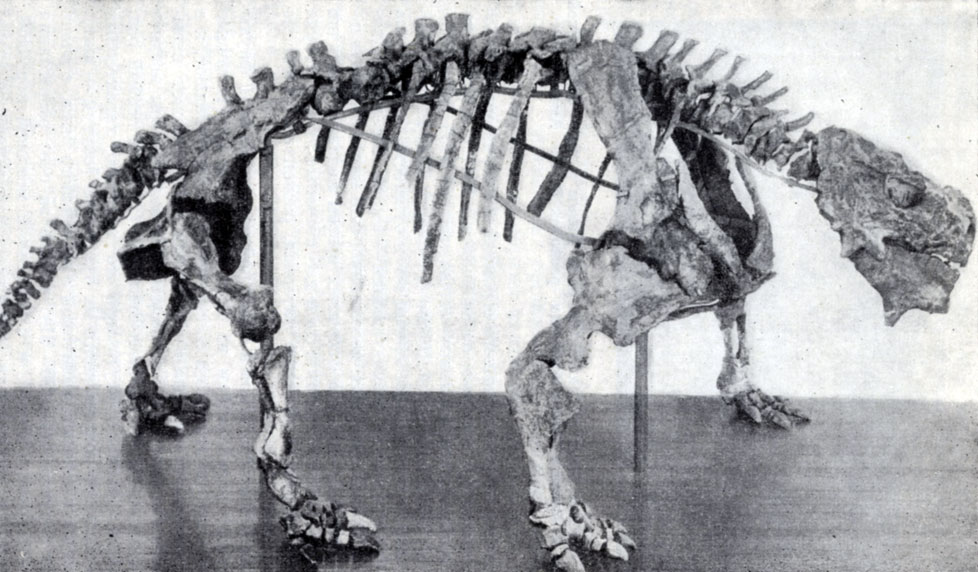
Рис. 89. Скутозавр (Scutosaurus karpinskii Hartmann-Weinberg), парейазавр из верхнепермских отложений (татарский ярус) Северной Двины. Раскопки В. П. Амалицкого. Скелет передан Академией наук СССР в дар Академии наук Китайской Народной Республики
Северодвинские парейазавры - скутозавры - в общих чертах строения сходны с южноафриканскими, но отличны в деталях: слиянием некоторых костей запястья и предплюсны, упомянутым выше сильным развитием кожных окостенений и некоторыми чертами строения мозговой коробки (внутри нее наблюдается костное образование вроде тенториума, отделяющего у млекопитающих большой мозг от мозжечка). В сборах с Северной Двины, кроме крупных скелетов, найден скелет маленького молодого парейазавра; с другой стороны, интересны отдельные кости огромных особей, раза в полтора превосходивших размерами обычных взрослых.
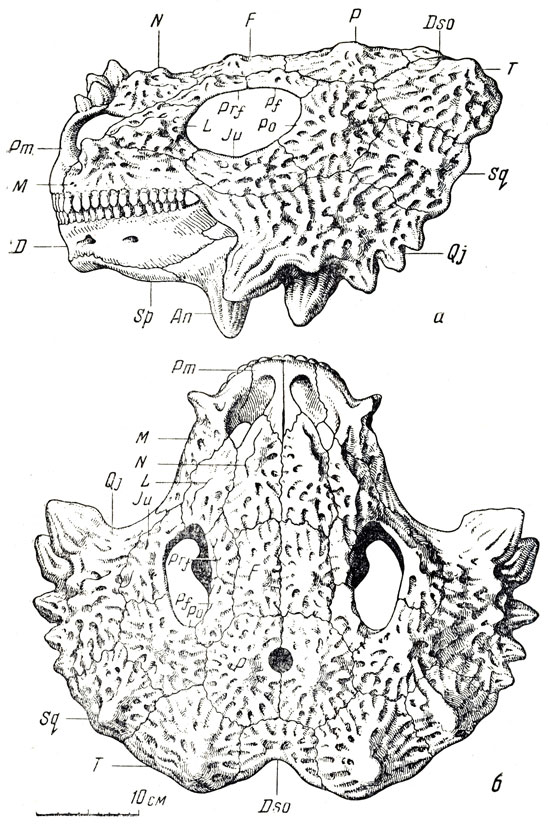
Рис. 90. Скутозавр (Scutosaurus karpinskii Hartvann-Weinberg). а - череп сбоку, б - сверзу (А. П. Быстров, 1957)
За последние десятилетия из верхнепермских отложений того же возраста на Волге и в Приуралье описаны и другие виды и роды скутозавров (антодон, парей-язухус).
Очень интересен более ранний и примитивный небольшой с мелкими не вполне текодонтными зубами парабрадизавр с р. Камы. Он происходит, правда, из более ранних отложений, относимых к казанскому ярусу. Однако их рассмотрение отвлекло бы нас в сторону от основной цели - дать общий обзор северодвинской фауны.
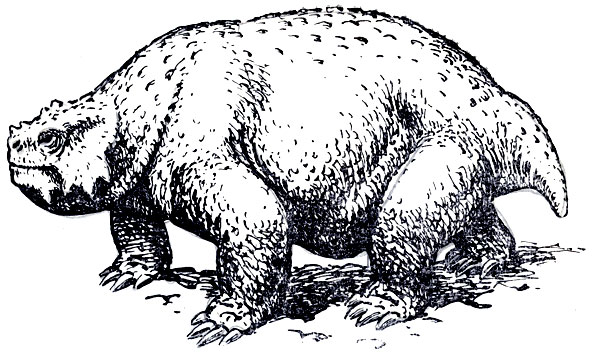
Рис. 91. Скутозавр Реконструкция внешнего вида (рис. К. К. Флерова)
Было бы трудно отчетливо характеризовать образ жизни парейазавров. В сущности, растительно-ядность - единственное, что представляется не вызывающим сомнения, хотя сами растения, служившие пищей, не могут быть названы с достоверностью. Массивность и форма плечевой кости, напоминающей плечевую кость крота и некоторых других роющих животных, невольно наводили на мысль о приспособлении к рытью и о роющем образе жизни. С другой стороны, об этом не говорят ни строение концевых фаланг пальцев, ни кисти. Архаическая постановка плечевой кости, унаследованная от древних предков, при тяжеловесности животного должна была вызывать огромную нагрузку на мускулатуру и скелет передней конечности, что уже само по себе могло стимулировать образование массивной плечевой кости и, вероятно, лишь чисто внешнее, конвергентное ее сходство с плечевой костью крота.
Л. Д. Бунстра (1932), посвятивший ряд работ строению скелета и реконструкции мускулатуры южноафриканских парейазавров, их эволюции и т. д., предполагает, что эти пресмыкающиеся по образу жшни напоминали бегемотов, казались в большинстве неуклюжими на суше, но, подобно бегемотам, могли быть хорошо приспособлены к водной среде.
В самой северодвинской фауне пока не обнаружено других котилозавров, хотя вряд ли можно сомневаться в их наличии в то время в ее составе. Замечательно интересные мелкие котилозавры из проколофонид, именно никтифрурет (рис. 92, 93) и никтеролетер (рис. 94) описаны И. А. Ефремовым (1940) из приблизительно одновозрастной верхнепермской фауны из красноцветных глин с р. Мезени. Это небольшие подвижные ящерицеобразные пресмыкающиеся длиной всего 30-40 см с очень глубокой, как у лабиринтодоптов, слуховой вырезкой в задней части крыши черепа, очень большим теменным глазом и огромными орбитами. Последнее наводит на предположение сумеречном или даже ночном образе жизни этих животных, видимо, питавшихся насекомыми, червями и вообще мелкой добычей. Эти или близкие проколофониды очень вероятны и для северодвинской фауны.
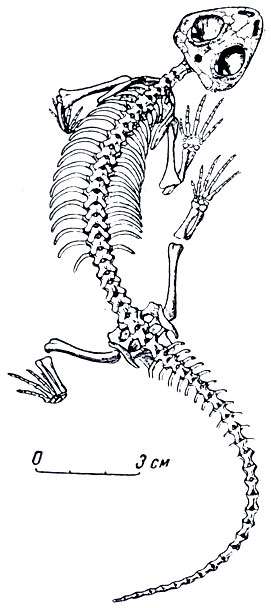
Рис. 92. Никтифрурет (Nyctiphruretus acudens Efremov). Верхнепермские отложения реки Мезени (И. А. Ефремов, 1940)
В южноафриканской "парейазавровой" фауне из формации Карру очень многочисленны и разнообразны дицинодонты. Там из десяти черепов позднепермских пресмыкающихся обычно чуть не девять принадлежат дицинодонтам. В северодвинской фауне также имеются дицинодопты, но с достоверностью пока установлен лишь один вид - дициподонт Траутшольда (Г. А. Траутшольд - проф. Петровско-Разумовской с.-х. Академии, 1817-1902); остатки дицинодонта (рис. 95) сравнительно с парейазаврами немногочисленны, хотя имеется несколько черепов хорошей сохранности, плечевой пояс и другие кости. Это типичный дицинодонт, снабженный только верхними клыками, крупными у самцов, с черепахообразной мордой, вероятно, при жизни с таким же, как у черепах, мощным роговым чехлом на ее конце и с необычайно широким для размеров черепа затылком, что говорит об очень сильной шейной мускулатуре. На материалах по дицинодоптам, добытых раскопками Амалицкого, выполнил интересные работы П. П. Сушкин (1922, 1923, 1920), но, тем пе менее, северодвинские дицинодонты еще не изучены полностью. Известны и разные точки зрения на их образ жизни. Очень большая поверхность затылочной стороны черепа и, очевидно, могучая шейная мускулатура натолкнули П. П. Сушкипа на мысль о приспособлении этих дицинодоптов к разрыванию па куски и поеданию трупов погибших животных, другими словами, к "падалеядному" образу жизни. С другой стороны, полная утрата зубов (за исключением клыков, достигающих у северодвинских дицинодонтов, главным образом у самцов, очень крупных размеров) как будто бы противоречит нашим обычным представлениям о падалеядпости, а почти несомненное наличие, как у черепах, мощного рогового чехла на конце морды разных аномодонтов наводит на мысль о растителыюядности дицинодонтов (О. Абель, 1927, и др.). Впрочем, мы знаем, что многие черепахи, озерные и морские, несмотря на беззубость, - очень сильные хищники, поедающие крупную рыбу, опасные и для других животных, включая человека.

Рис. 93. Никтифрурет (Nyctifmretus acudens Efremov) Реконструкция внешнего вида (рис. А. П. Быстрова)
Попутно можно упомянуть о том, что за последние годы у нас обнаружены, правда, в нижнетриасовых отложениях другие новые дицинодонты, в том числе гигантские с клыками до полуметра длиной, в общем приближавшиеся по размерам к достигшим гигантизма, но беззубым шталекериям из триаса Бразилии. По для пермских отложений наши материалы сравнительно с южноафриканскими пока как будто бы менее богаты дицинодонтами. Не исключена возможность и значительного разнообразия образа жизни у разных дицинодонтов - растительноядность, всеядность и плотоядность.
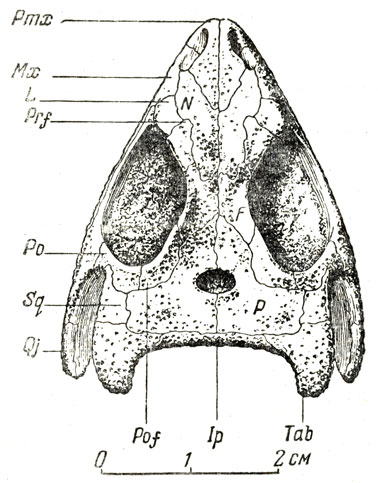
Рис. 94. Череп никтеролетсра (Nucteroleter ineptus Efreinov). Верхнепермские отложения р. Мезени (И. А. Ефремов, 1940)
Если образ жизни дицинодонтов и их пища не вполне ясны, то никаких сомнений не вызывают заведомо хищные пресмыкающиеся северодвинской фауны и прежде всего огромный горгонопс - инострапцевия (рис. 96-97) - род, названный так Амалицким в честь своего учителя, профессора Петербургского университета А. А. Ипостранцева (1843-1919).
При раскопках на Северной Двине найдены два почти полных скелета инострапцевии; не сохранились лапы и конец хвоста. (Из-за трудности препаровки передние и задние конечности одного из скелетов остались в смещенном состоянии вследствие чего этот скелет до сих пор остался с положением конечностей, свойственным млекопитающим, с вертикально поставленными ногами, что, разумеется, не соответствует их естественному прижизненному положению.)
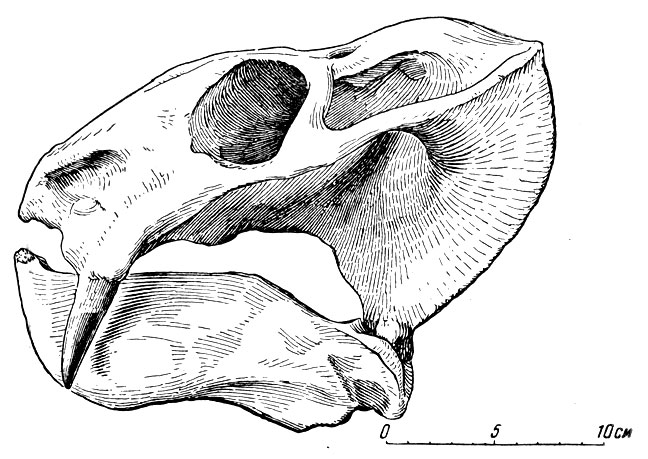
Рис. 95. Дициподопт (Dicynodon irautscholdi Amalitzky). Верхвевермские отложения Северной Двины. Раскопки В. П. Амалицкого
Для инострапцевии характерен мощный тяжеловесный череп при относительно легком и стройном остальном скелете, говорящем о подвижности и проворстве животного. Но прежде всего обращает на себя внимание череп (рис. 96), его высокая сжатая с боков лицевая часть, высокая (особенно в симфизе) нижняя челюсть, расположенные совсем рядом на конце морды наружные носовые отверстия, огромные свисающие далеко вниз кинжалообразные клыки и кинжалообразные же резцы. Верхние резцы длотно смыкаются с нижними, входя в промежутки между ними, и наоборот, а нижние клыки своими вершинами входят в ямы на нёбе. Послеклыковые щечные зубы, сравнительно слабые и немногочисленные, сохранились в верхней челюсти, но, видимо, не играли большой роли; нёбные и птеригоидные зубы совсем незначительны. Вся мощь этой страшной пасти явно сосредоточена в передней части черепа и зубов. Верхние клыки, пильчатые по заднему краю, снабжены огромными корнями и напоминают клыки саблезубых кошек; это сходство увеличивается и наличием желобообразных углублений на внешней стороне нижних челюстей; в эти желоба как в полуножны, ложатся длинные кинжаловидные клыки при закрытой пасти. По-видимому, размах пасти при хватании добычи должен был быть очень велик, чтобы хищник мог вонзить в добычу свои клыки, вообще схватить ее. Одновременно обращает на себя внимание незначительная величина височных впадин, лишь немного превосходящих размерами орбиту; это, вероятно, говорит о меньшем развитии и большей примитивности мускулатуры, закрывающей пасть, чем можно было бы ожидать для такого крупного хищника. У найденных экземпляров длина черепа достигает полуметра и более, а общая длина животного наверное была около четырех метров, из которых, очевидно, около половины приходилось на хвост. Но в сборах Амалнцкого с Северной Двины имеются отдельные кости особей, превосходивших в полтора, если не в два раза скелеты, выставленные в Музее Палеонтологического института Академии наук СССР.
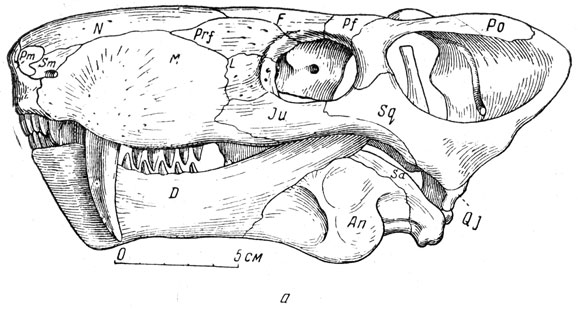
Рис. 96. Верхнепермские горгонопсы. а - череп зауроктона (Sauroctonus progressus Hartmann-Welnberg) из верхнепермских отложений правого берега Волги у села Ильинского около г. Тетюши (А. П. Быстрое. 1955)
Остатки иностранцевий, или близких им горгонопсов, известны из других одповозрастпых местонахождений верхнепермских отложений СССР.
Б. П. Выошков (1954) предполагает, что в северодвинской фауне было несколько видов ипостранцевий. Небольшой горгонопс выделен в отдельный род и вид православлевия (в честь проф. П. А. Православлева, 1873-1941).
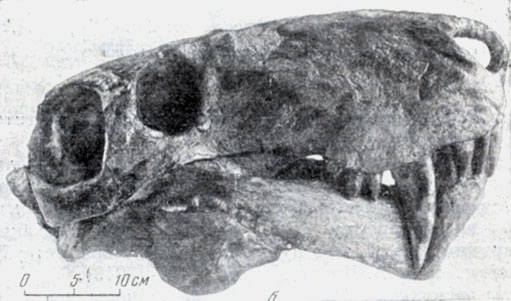
Рис. 96. Верхнепермские горгонопсы. б - череп шюстранцешщ ( Inostrancevia alexandii Amalitzky) из верхнепермских отложений Северной Двины. Раскопки В. П. Амалицкого
Из местонахождения этой же фауны, но в среднем течении Волги А. П. Быстровым описан (1955) небольшой новый горгонопс зауроктон, который строением черепа напоминает южноафриканского элурогната и сохраняет не особенно удачное видовое название "прогрессус" из-за предполагавшейся А. П. Гартман-Вейнберг (1938) близости к цинодоптам (они более млекопитающеподобны, чем горгонопсы). В более ранних верхнепермских отложениях И. А. Ефремовым (1954, 1955) установлены более примитивные, небольшие горгонопсы, фтипозухи, несущие черты строения, сближающие этих рептилий с архаическими хищными дейпоцефалами СССР, напоминающими еще нижнепермских сфенакодонтных пеликозавров Северной Америки (см. фтинозухид).

Рис. 97. Иностранцевия Реконструкция внешнего вида (рис. К. К. Флерова)
В составе северодвинской фауны был обнаружен небольшой, очень интересный цинодонт - двиния (рис. 98), названный так Амалицким по р. Северной Двине и первоначально описанный по неполной лицевой части черепа, впоследствии подробнее изученный П. П. Сушкиным по другому почти полному черепу под названием пермоцинодон, а недавно, вновь, Е. Д. Конжуковой (1949). Череп, длиною 10 см, очень напоминает череп примитивных млекопитающих своими общими пропорциями, за исключением, разумеется, мозговой коробки, совсем небольшой у двинии. Для двинии характерно отчетливо выраженное вторичное нёбо, образовавшееся за счет срастания отростков нёбных и верхнечелюстных костей. Сильное развитие венечного отростка нижней челюсти, очень крупные височные ямы и широкие скуловые дуги, вероятно, говорят о сильной и, возможно, хорошо дифференцированной жевательной мускулатуре. Лицевая часть черепа внешне очень напоминает таковую небольшого хищного млекопитающего, а бугорчатые передние и задние щечные зубы со сложно устроенной коронкой еще больше подчеркивают это сходство с млекопитающими. Наблюдается увеличение числа бугорков и усложнение коронки от передних щечных зубов к задним (за исключением последнего); это также говорит о сходстве с млекопитающими. Оно усиливается и некоторыми особенностями строения остального скелета; например большим объемом таза, направленностью подвздошной кости вперед, полушаровидной формой головки бедра, которая па две трети обращена своей сочленовной поверхностью медиально, общей формой вертлужпой впадины и т. д. Тем не менее строение мозговой коробки, наличие отверстия для теменного глаза и сложный состав нижней челюсти из обычного для рептилий (и птиц) "набора" нескольких костей явственно указывают на то, что мы имеем дело с пресмыкающимся, что, несмотря на всю свою "млекопитающеподобность", двиния стоит в стороне от прямого пути в направлении млекопитающих. На это указывает также и характер строения зубов, высоко специализированных и не могущих быть щредочными для зубов первых млекопитающих.
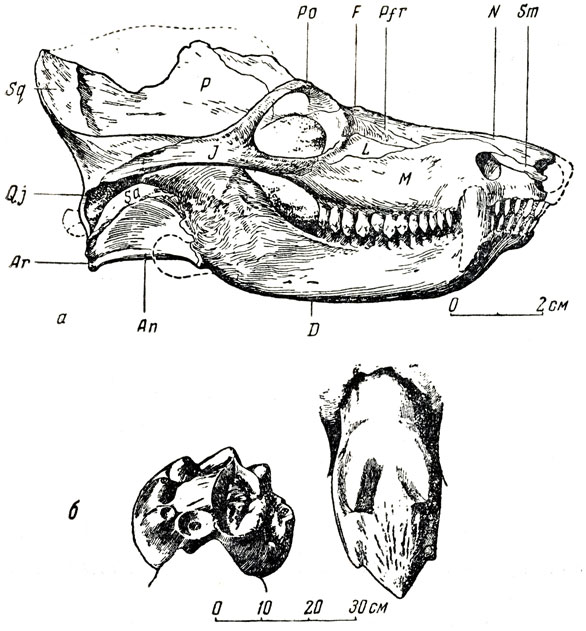
Рис. 98. Двиния (Dvinia suschkini Amalitzky). а - череп; б - задний щечный зуб (Е. Д. Кошкукова, 1949). Верхнепермские отложения Северной Двины. Раскопки
Среди синапсидных пресмыкающихся северодвинской фауны имеется тероцефал "анна петри", названный так Амалицким в честь Анны Петровны - жены и верной помощницы ученого в течение всей его научной деятельности. Анна петри (рис. 99) - тероцефал средних размеров с черепом до 25 см длины, с характерным для тероцефалов узким сагиттальным гребнем, простыми слегка загнутыми назад остроконическими зубами и задними "нёбными" отверстиями на уровне глазниц. Лицевая часть черепа широкая, весь череп низкий. Теменной канал отсутствует, но явно видны следы его зарастания. Очевидно, мы имеем дело с "отказом" животного с возрастом от теменного органа и с атрофией этого органа, во всяком случае его дистальной части, помещавшейся в теменном отверстии.
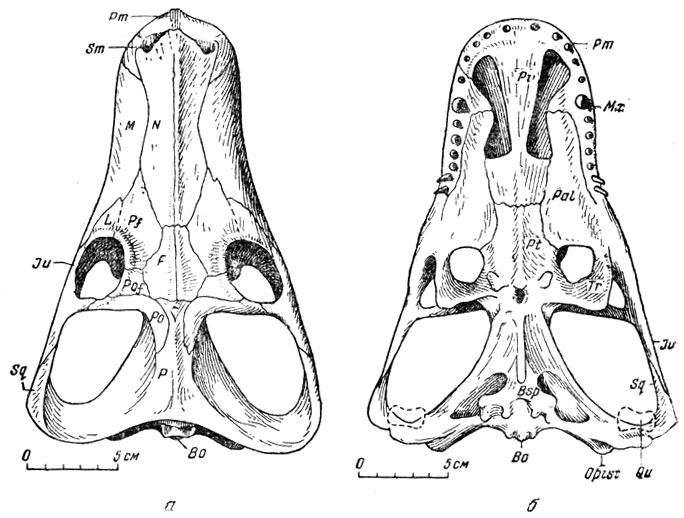
Рис. 99. Анна петрн (Anna petri Amalitzky). а - череп вид сверху; б - снизу Верхнепермские отлошешш Северной Двины. Раскопки В. П. Амалицкого
Орбиты анна петри имеют форму наклонно стоящих овалов в соответствии с укороченной и уплощенной формой черепа. Они обращены вверх, вперед и в стороны.
Верхнепермские отложения СССР доставили нам и других тероцефалов. Упомянем хтонозавра из одновозрастных с северодвинскими отложений Приуралья.
Хтонозавр характеризуется в отличие от анна петри суженным черепом, длинной и относительно слабой нижней челюстью; зубы поставлены редко и на значительном протяжении. Оба рода входят.в состав семейства Annidae, довольно обособленного от южноафриканских тероцефалов. Известны и другие, но пока разрозненные находки.
В северодвинской фауне имеется очень своеобразное животное, получившее от Амалицкого (1921) родовое название котлассия по г. Котласу, около которого велись раскопки этой фауны. Котлассия (рис. 100) была снабжена защитным покровом в виде скульптурированных щитков, располагавшихся несколькими рядами вдоль позвоночного столба на некотором его протяжении. Часть щитков несла на нижней стороне пластинчатые выросты, вероятно, внедрявшиеся прочно в нижележащую соединительную ткань кожи (рис. 100, б). Амалицкий отнес кот-лассию к сеймуридам, которые тогда считались анапсидными пресмыкающимися, котилозаврами. П. П. Сушкин (1925, 1927, 1928) на основе некоторых особенностей строения столбика уха ("стремечка"), квадратума и птеригоида котлассии и сеймурии пришел к заключению, что котлассия и сеймурия относятся к ископаемым земноводным, лабиринтодонтам, а не к пресмыкающимся. Ватсон (1914), Виллистон (1925), Броили (1927) стояли на позициях "котилозавровости", т. е. "рептилыюсти" сеймурии и котлассии. А. П. Быстров (1944) тщательно допрепарировал котласоию, подробно описал ее и пришел к выводу, что по строению черепа (в частности его слуховой вырезки, столбика уха), зубов, переднего и заднего поясов конечностей, котлассия скорее земноводное, а не пресмыкающееся.
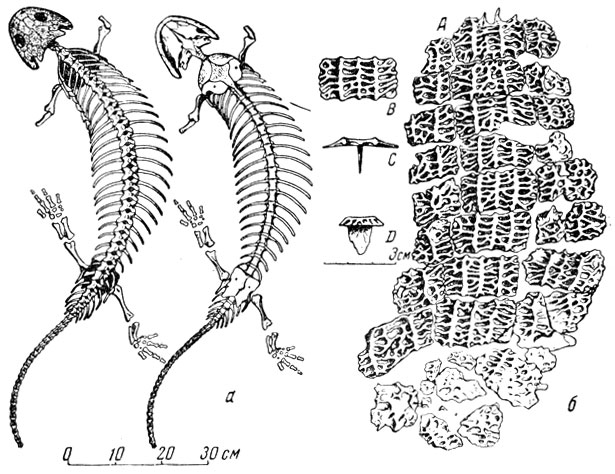
Рис. 100. Котлассия (Kotlassia prima Amalitzky) а - скелет с черепом, вид сверху и снизу; б - кожные Окостенения. Верхнепермские отложения Северной Двины (А. П. Быстров, 1944)
Действительно, не только череп котлассии во многом такой, как у лабиринтдонтов, но и зубы, и их складчатая стенка с тем же числом складок, как и на одинаковых по размерам зубах лабиринтодонта бентозуха из низов триаса; чередование тонкостепных молодых зубов и толстостенных старых, т. е. характер смены зубов, - все это, по мнению Быстрова, черты, типичные для лабиринтодонтов. "Котилозавровый" характер позвонков котлассии, по мнению Быстрова, говорит о реликтовом характере животного, вероятно, связанного происхождением со стегоцефалами, давшими начало котилозаврам.
Несколько позднее И. А. Ефремов (1946), изучив верхнепермского лантанозуха (рис. 87), объединяющего в себе подобно котлассии черты земноводных (лабиринтодонтов) и пресмыкающихся (котилозавров), предложил обе эти формы вместе с сеймуриями выделить в особый подкласс батрахозавров, промежуточный между амфибиями и рептилиями. Материал по этим формам в СССР значительно возрос за последние годы, хотя и носит пока (за исключением лантанозуха) очень разрозненный характер. В последнее время А. Ромер склоняется к тому, что лантанозух, быть может, относится к котилозаврам. Напомним, что Ромер (A. Romer, 1945, 1947, 1956) поместил сеймурии и котлассию вместе с дипловертеброятидами в особый отряд лабиринтодонтов под названием сеймуриаморф; И. А. Ефремов их считает частью особого подкласса батрахозавров. Необходимо иметь в виду, что все известные батрахозавры, несмотря на совмещение в себе одновременно признаков строения земноводных и пресмыкающихся, не могут считаться переходом от амфибий к рептилиям; уже по одному своему молодому геологическому возрасту это все лишь реликтовые формы.
Таковы вкратце пресмыкающиеся северодвинской фауны. В общих чертах это до такой степени "та же" фауна, что и в зоне Cistecephaius Южной Африки, что ее обнаружение Амалицким на севере Северного полушария, да еще в подтверждение смелого предсказания Амалицкого об ее предстоящем открытии, было крупным событием в палеонтологии позвоночных. Естественно встал вопрос о возникновении фауны, о путях ее расселения (из Африки в Северную Европу, или наоборот), о центре формирования и т. д. Подробный анализ имеющихся воззрений занял бы много времени и увел бы нас в область пока не разрешенных вопросов истории Земли. Отметим лишь, что некоторые накопившиеся за последние годы данные о меридиональном распространении области континентальных отложений (в частности угленосных толщ Ангарской суши), т. е. о меридиональном" на юг распространении гондванских фаций, ставят, по мнению И. А. Ефремова (1952) и некоторых других, под вопрос резкое разделение всей палеозойской суши на гигантские южную Гондвану и северную Лауразию. (Возникла даже мысль о "меридиональной" климатической зональности и возможности меридионального в верхнем палеозое положения экватора относительно плоскости эклиптики и т. п.). Несомненным остается широкое в то время распространение парейазавровой фауны.
В заключение обзора северодвинской фауны уместно отметить относительную бедность сборов лабиринтодоптами. Во всяком случае в составе этой фауны мы до сего времени имеем, если не считать котлассию, лишь одного достоверного лабиринтодонта - двинозавра (рис. 101).
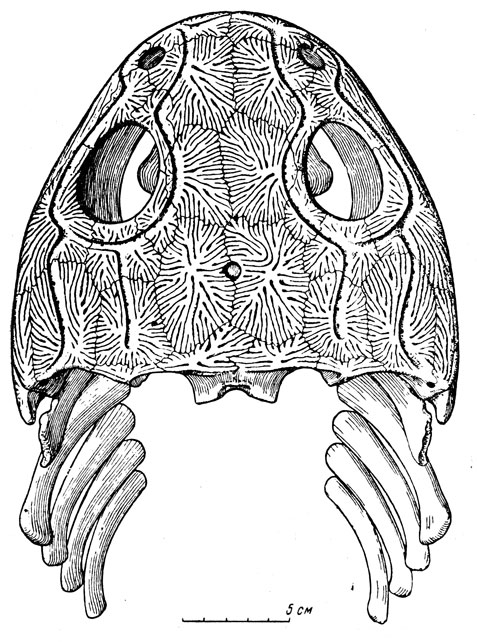
Рис. 101. Двинозавр (Dvinosaurus primus Amalitzky). Череп со скелетом жаберных дуг, вид сверху. Верхнепермские отложения Северной Двины (А. П. Быстров, 1938)
По данным Д. Ватсона, П. П. Сушкина (1928, 1936) и А. П. Быстрова (1935, 1938, 1939-1947), этот рахитомный лабиринтодонт характеризуется "личиночным" характером строения не только кожных костей крыши черепа, но и строением скелета жаберных дуг, а также представляет собою не только постоянножаберпую ("переннибронхиатную") форму, как предполагали Д. М. Ватсон и П. П. Сушкин (1936), но и неогеническую личиночную, которая подобна аксолотлю современной мексиканской саламандры амблистомы.
Эти данные вполне согласуются с имеющимися предположениями относительно общей аридности обстановки, в которой обитала северодвинская фауна; но многочисленность парейазавров сама по себе с убедительностью говорит одновременно и об изобилии какой-то растительности, служившей пищей этим крупным животным, по мнению Бунстра, бегемотоподобным по образу жизни.
Все изложенное в этой главе имело в виду лишь самый общий обзор двух фаун, точнее двух фаунистических комплексов, без какой-либо претензии на полноту данных о строении^ образе жизни, палеогеографическом распространении перечисленных позвоночных и т. д. Подробное рассмотрение всех этих вопросов вышло бы далеко за пределы данного обзора.
Необходимо упомянуть в заключение, что за последние десятилетия в СССР обнаружены и другие верхнепермские комплексы, а в самое последнее время на севере Европейской части СССР сделаны замечательно интересные находки примитивных котилозавров капторинид типа Cotylorhinchus и эдафозавров и других, считавшихся чисто американскими формами. Большими раскопками Палеонтологического института добыта интереснейшая новая верхнепермская фауна около г. Очера, ана еще не изучена. Имеются все основания предполагать, что постановка раскопок вновь открытых местонахождений принесет нам в самом недалеком будущем много нового, подчас совершенно неожиданного, даст новое освещение фауне континентальных пермских отложений и многим вопросам стратиграфии, палезоогеографии и палеогеографии этого замечательного периода в истории не только рептилий, но истории Жизни и Земли вообще.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'