
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Из истории динозавров
Динозавры - основная группа мезозойской фауны наземных позвоночных. Предки динозавров - текодонты. Развитие двуногого хождения. Гигантизм хищников. Переход к растительной пище и четвероногому хождению. Гиганты Земли - зауроподы. Загадка птицетазовых динозавров. Открытие гетеродонтозавра. Приспособление к обитанию в воде у орнитопод. Оборона стегозавров, панцирных и рогатых динозавров от хищников. Пробелы в истории динозавров. Почему вымерли динозавры?
После знакомства с отдельными представителями динозавров, открытых в Центральной и Средней Азии, можно обратиться теперь к истории этой группы, самой многочисленной и разнообразной среди наземных позвоночных мезозоя. Современниками динозавров, населявшими континенты, были прежде всего их ближайшие родственники - крокодилы и летающие ящеры, или птерозавры, объединяемые вместе с динозаврами в общий подкласс архозавров ("древних ящеров"). Второй подкласс составляют черепахи, но, подобно крокодилам и летающим ящерам, они не дали большого разнообразия форм, свойственного динозаврам. Другие известные нам наземные рептилии - ящерицы и змеи, появившиеся в середине мезозоя, оставались на протяжении "средней эры жизни" немногочисленными группами, получившими быстрое развитие и широкое распространение лишь в кайнозое. Еще более редкими были птицы и млекопитающие, игравшие весьма незначительную роль в составе мезозойской фауны наземных позвоночных. Два довольно обширных подкласса мезозойских рептилий - ихтиозавры и так называемые синаптозавры, наиболее известными представителями которых являются плезиозавры, - населяли моря. Наконец, два самых ранних подкласса рептилий - котилозавры и зверообразные - полностью вымирают в триасе, когда появляются первые динозавры. Вместе с древними рептилиями в триасе вымирают и последние крупные амфибии - лабиринтодонты, более известные под старым названием стегоцефалов. Таким образом, динозавры оказываются ведущей мезозойской группой наземных позвоночных, очень быстро развивающейся и достигающей широкого, всесветного распространения. Поэтому с полным основанием мезозой называют "эрой динозавров".
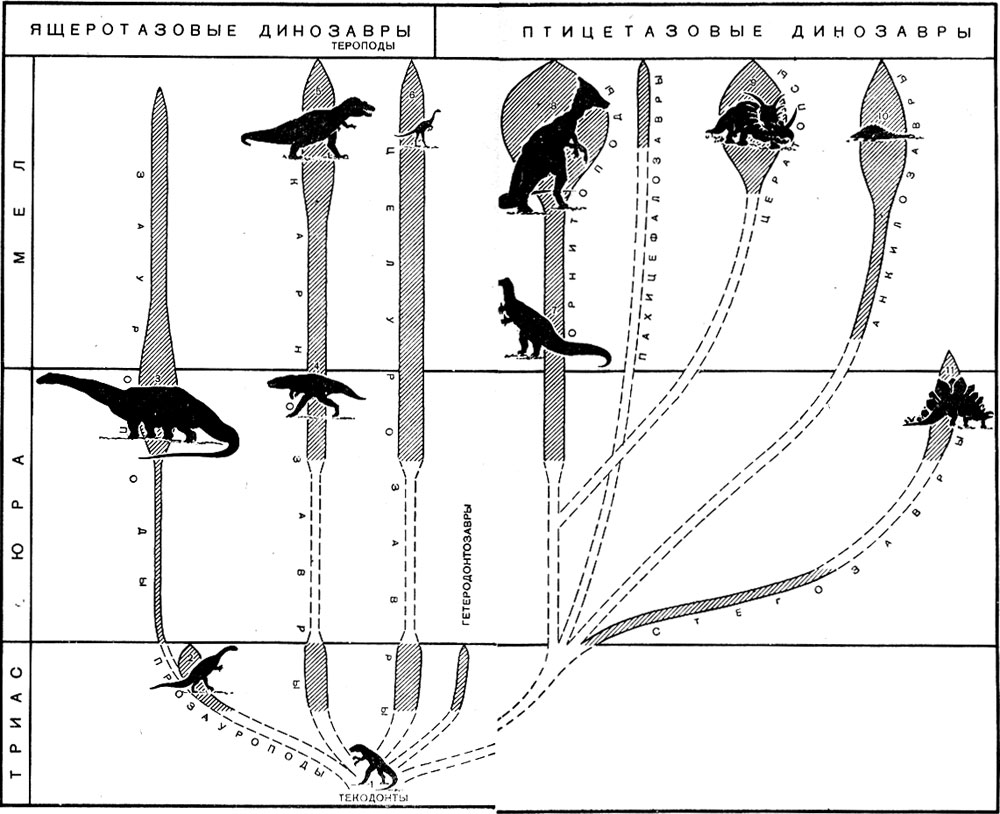
Схема эволюции динозавров
Архозавры, помимо динозавров, крокодилов и летающих ящеров, включают еще один надотряд пресмыкающихся - текодонтов, т. е. "ячеистозубых", названных так за то, что зубы у них сидели в глубоких ячейках - альвеолах, в отличие от других рептилий, у которых зубы прирастают к стенкам челюстей. Текодонтность зубов - характерный признак всех архозавров. Текодонты - очень важная в эволюционном отношении группа, так как именно от них в середине триаса или чуть позже произошли все остальные архозавры и птицы, которых зоологи выделили в самостоятельный класс, хотя с точки зрения эволюционной систематики они должны иметь такой же ранг, как и остальные группы архозавров. Вопрос о происхождении самих текодонтов пока недостаточно ясен, - одни палеонтологи считают их потомками пермских эозухий, другие - потомками котилозавров, из которых выводят вообще все основные группы (подклассы) рептилий.
Текодонты, появившиеся в самом начале триаса, вымирают в конце этого периода, уступив жизненную арену своим более прогрессивным потомкам - в первую очередь динозаврам и крокодилам, ранние представители которых так похожи на своих предков, что иногда их даже трудно различить. Поэтому указания на находки средне-триасовых крокодилов и динозавров могут быть взяты под сомнение, соответствуя в действительности, возможно, текодонтам. Последние были сравнительно "небольшими" хищными пресмыкающимися до 4-5 метров длины. Одни из них жили в воде и напоминали крокодилов, а другие, похожие на крупных ящериц, обитали на открытых пространствах. Сухопутные текодонты приспособились к двуногому хождению - на задних лапах, которые у них были значительно длиннее передних. Двуногое хождение обеспечивало текодонтам высокий наблюдательный пост, облегчая поиски добычи. Именно от таких текодонтов и произошли динозавры, унаследовавшие двуногий способ передвижения. Летающие потомки текодонтов - птерозавры и птицы - скорее всего, произошли от лазающих древесных форм, перешедших со временем к планирующим прыжкам-полетам.
Из динозавров первыми появились хищные небольшие и легкие подвижные формы, которые вскоре - все это происходило в конце триасового периода - дали по крайней мере две ветви. Их главная ветвь привела в конце мезозоя к гигантским карнозаврам - тарбозаврам, тираннозаврам и им подобным, достигавшим в высоту (на задних ногах) 5-6 метров и больше.
Развитие гигантизма у карнозавров, по-видимому, было связано с параллельным развитием гигантизма у растительноядных динозавров, служивших им пищей. Для хищников крупный размер означал возможность большой добычи, так же, как и для растительноядных животных крупный размер являлся одной из форм защиты от хищников. Так было не только у динозавров, но и на протяжении всей истории животного мира. Между хищниками и растительноядными животными происходило своего рода "соревнование" в росте, хотя конечно, всегда был какой-то предел, определявшийся комплексом биомеханических и физиологических закономерностей.
Гигантизм карнозавров начался с развития зубов и черепа в целом как главного орудия нападения: большие зубы-кинжалы необходимы были для закалывания крупных жертв. Возможно, увеличение головы не могло бы достигнуть у карнозавров таких больших размеров, если бы параллельно не стали уменьшаться передние конечности, облегчая тем самым переднюю часть тела. Ее противовесом служил хвост, но это могло оказаться недостаточным без уменьшения веса передней части тела, и карнозавру волей-неволей пришлось бы опуститься на все четыре ноги, потеряв свой высокий наблюдательный пост при двуногой позе.
Параллельно с карнозаврами, обладавшими огромной головой, с сильными зубами и сократившимися до мизерных размеров передними лапами, развивалась другая ветвь птицеподобных хищных динозавров - целурозавров. Их поздние представители (струтиомимус и другие) имели облегченную конструкцию: маленькую голову с клювом, лишенным зубов, и пустотелые кости. Передние лапы у них были цепкими и хорошо развитыми, задние - с длинной цевкой, как у птиц, приспособленными для быстрого бега.
Наконец, среди хищных динозавров были, вероятно, и узко специализованные группы, приспособившиеся к питанию общественными насекомыми (джилантайзавры - теризинозавры) и, возможно, плодами покрытосемянных (дейнохейрусы). Эти группы были экологическими предшественниками неполнозубых.
Такая трактовка образа жизни некоторых хищных динозавров, кажущаяся на первый взгляд, необычной, на самом деле имеет серьезное обоснование. Дело в том, что вторая половина мезозоя - время уже достаточно широкого распространения покрытосемянных, или цветковых растений и связанного с ними развития различных групп насекомых, в том числе и перепончатокрылых, многие группы которых образуют большие сообщества, или колонии. Естественно, что появление больших пищевых ресурсов в виде концентраций насекомых, съедобных плодов растений, устраиваемых на деревьях гнезд птиц и, возможно, птерозавров (и та, и другая группы опять-таки характерны для второй половины мезозоя) должны были найти и потребителей, которыми в первую очередь и могли оказаться сухопутные динозавры - хищники.
В кайнозое эти адаптации динозавров повторили различные группы млекопитающих (неполнозубые, приматы и др). В настоящее время везде, где есть насекомые, живущие колониями, имеются и их потребители: в Америке - муравьеды и броненосцы, в Африке - трубкозубы и частично панголины, в Южной Азии - панголины. В Австралии эту функцию приняли на себя сумчатые (сумчатый муравьед, когти которого на передних лапах удивительно похожи на серповидные фаланги карнозавров!), а в наших широтах эти обязанности "по совместительству" выполняют медведи.
И карнозавры, и целурозавры остались двуногими формами. В то же время от хищных динозавров отделилась боковая ветвь - прозауроподы, от которых позднее произошли зауроподы - гигантские четвероногие динозавры, такие, как диплодок. Прозауроподы жили очень недолго - в самом конце триасового периода. Это были уже крупные, до 8-10 метров длины, животные, начавшие переход к четвероногому хождению: их передние конечности не намного короче задних. Зубы у прозауропод не такие острые и сильные, как у хищных динозавров. Голова у них осталась небольшой. Возможно, они были всеядными животными, а их потомки зауроподы окончательно перешли на питание растительностью, и зубы их стали похожи на долотца. Не исключено, однако, что некоторая степень всеядности (за счет мелких водных животных) сохранилась и у зауропод. Они приспособились к жизни в больших водоемах, найдя, возможно, в них спасение от своих кровожадных собратьев. Жизнь в воде, где при передвижении облегчается, по сравнению с сушей, нагрузка на скелет, позволила зауроподам достигнуть чудовищных размеров - они превосходили в 2-3 раза самых крупных наземных животных. Как известно, наиболее крупные современные животные - киты и гигантские кальмары - также живут в воде. На суше такие животные не смогли бы существовать из-за огромного веса. При быстром движении их тело невероятно перегревалось бы благодаря огромному количеству теплопродукции, выделяемой при этом. Здесь вступают в силу физиологические ограничители, которые, конечно, есть и для водных животных, но у последних они, несомненно, большего диапазона. Огромная нагрузка на задние конечности при двуногой позе лимитировала бы гигантизм зауропод, если бы они "вовремя", не перешли к четвероногому хождению. Однако, возможно, они сохранили способность вставать на задние ноги в воде (чтобы подышать воздухом и оглядеться) и благодаря этому могли заходить на значительную глубину. Зауроподы - это "динозавры-водолазы".
Если у наземных хищников - карнозавров - нагрузка на скелет была велика и суставы костей вследствие этого имеют вид хорошо отработанных шарниров, то у зауропод с уменьшением нагрузки на скелет в воде это утратилось, а эпифизы (концы) костей остались хрящевыми, позволяя дальнейшее увеличение в размерах. Неимоверно длинный хвост зауропод служил для плавания, а почти такая же длинная шея позволяла животному "обедать", не сходя с места, что экономило его энергию. Накормить такую махину было не так просто! Правда, нужно иметь в виду, что рептилиям с их более замедленным обменом веществ по сравнению с теплокровными и более активными птицами и млекопитающими требуется и соответственно меньшее количество пищи. В то же время не исключена возможность, что динозавры были теплокровными животными.
В этом отношении интересны исследования профессора И. Д. Стрельникова, изучавшего физиологию пресмыкающихся и других позвоночных. Согласно его данным, постоянная температура тела, приведшая к теплокровности, исторически должна была возникнуть как регулятор теплообмена у животных, вырабатывающих большое количество тепловой энергии при движении, например во время полета в воздухе (птицы, летающие ящеры), либо у крупных животных (рептилии, млекопитающие). Температура тела у современных крупных рептилий крокодилов почти постоянная, колеблющаяся в пределах суток на несколько градусов (например, у аллигаторов от 27-28° до 32-35°), да и сердце у них четырехкамерное с полным отделением артериальной крови от венозной, как у птиц и млекопитающих. Все это дает основание полагать, что "двоюродные братья" крокодилов - динозавры - могли быть также теплокровными животными.
Зауроподы вместе с прозауроподами и хищными динозаврами объединяются в отряд ящеротазовых динозавров, которому противопоставляют отряд птицетазовых динозавров. Ко второму отряду относятся орнитоподы (игуанодонты, утконосые динозавры и другие), стегозавры, анкилозавры и рогатые динозавры. У ящеротазовых динозавров таз состоит из трех "лучей" - подвздошной, лобковой и седалищной костей (все они, разумеется, парные), как у всех рептилий и у большинства позвоночных вообще. У птицетазовых динозавров таз состоит из четырех "лучей" благодаря тому, что лобковая кость имеет хорошо развитый задний отросток, сохранившийся также у птиц. Возможно при этом, что утрата птицами основной или передней части лобковой кости является приспособлением к носке более крупных яиц как обладающих соответственно большим запасом питательных веществ и тем самым обеспечивающих лучшее по сравнению с рептилиями развитие зародышу.
Все птицетазовые динозавры были растительноядными животными. Относительно происхождения птицетазовых динозавров имеются две точки зрения: одни палеонтологи считают их потомками ранних хищных динозавров, другие рассматривают как самостоятельную ветвь, ведущую свое начало непосредственно от текодонтов. До последнего времени считалось, что первые птпцетазовые динозавры появились в начале юры, пока несколько лет назад в Южной Африке в отложениях верхнего триаса не был найден небольшой, величиной около метра, птицетазовый динозавр, названный гетеродонтозавром, т. е. "разнозубым ящером". Только у этого единственного динозавра задние зубы растительноядного типа, напоминающие зубы орни-топод, с притупленной коронкой и медиальным гребнем посередине, тогда как передние остались острыми, такой же формы, как у хищников. Возможно, гетеродонтозавр, сочетающий в себе признаки хищных и птицетазовых динозавров, и должен быть поставлен в начало родословной последних. Во всяком случае, гетеродонтозавр - пример перехода от хищного образа жизни к растительноядному, скорее всего, через всеядность, аналогично тому, как это случилось с нрозауроподами и случалось неоднократно на протяжении истории позвоночных. Находка гетеродонто-завра, если его не рассматривать как непосредственного, независимого от хищников потомка текодонтов, свидетельствует в пользу происхождения птицетазовых динозавров от хищных.
Из всех птицетазовых динозавров только орнитоподы ("птиценогие") сохранили хождение на задних лапах, унаследованное от их предков. Таковы игуанодонты (поздняя юра - ранний мел) и их потомки - утконосые динозавры (поздний мел).
Игуанодонты были обитателями суши, вероятно, прибрежных зарослей, где они находили обилие растительной пищи и возможность укрытия от хищных динозавров. Череп игуанодонта внешне гораздо больше похож на череп лошади, нежели любого пресмыкающегося. Это удивительное внешнее сходство дополняется и некоторым сходством в строении зубной системы. Как известно, лошадь имеет высокую, или так называемую гипсодонтную коронку зуба, которая развилась как противодействие быстрому снаши-панию зуба, поскольку при питании животного к нему в рот вместе с травой попадали и землистые частицы. Игуанодонты, а еще в большей степени утконосые динозавры, как бы предвосхитили гипсодонтность зубов лошади, но только у динозавров она достигнута за счет наличия трех зубов и более в одном вертикальном ряду. Число таких вертикальных зубных рядов у некоторых поздних утконосых динозаврам достигает 200, по 5-6 зубов в каждом. Утконосые динозавры, которых я уже достаточно подробно описал раньше, как и зауроподы, лишенные специальных защитных средств, спасались от хищных динозавров в водоемах, находя здесь себе и пищу.
Все сухопутные птицетазовые динозавры - стегозавры, панцирные и рогатые, утратили двуногое хождение и приобрели мощные защитные средства против хищных динозавров. Первыми перешли к четвероногому хождению стегозавры ("покрытые ящеры"), представляющие, скорее всего, боковую ветвь орнптопод. Ранние представители стегозавров, судя по пропорциям передних и задних конечностей, еще сохраняли черты двуногости, свойственной орнитоподам. Особенно многочисленными стегозавры были в конце юрского периода, достигая при этом внушительных размеров - 6 метров длины. Их спина и хвост были усажены в два ряда огромными костными шипами. Четвероногая поза защищала от хищников, нападавших сверху, не только спину, но и брюхо стегозавров.
В меловой период на смену стегозаврам появились панцирные динозавры (анкилозавры) и рогатые (цератопсы), которые тоже "рискнули" остаться жить на суше бок о бок со своими грозными собратьями - карнозаврами, ставшими к этому времени еще более крупными. По величине ни анкилозавры, ни цератопсы не уступали стегозаврам, но в дополнение к шипам стегозавров у анкилозавров появился еще и костный панцирь на теле из срастающихся щитков с мелкими шипами, а у цератопсов - от одного до пяти рогов на голове. У ранних анкилозавров и цератопсов в строении скелета также сохранились черты, указывающие на двуногое хождение и их вероятное происхождение от орнитопод, хотя некоторые палеонтологи считают анкилозавров потомками стегозавров. Поздние анкилозавры и цератопсы были типично четвероногими по способу передвижения. Эти ящеры вместе с гигантскими кар-нозаврами и утконосыми динозаврами завершают "летопись" истории, длившейся более 100 миллионов лет.
Несмотря на то что сейчас известны сотни видов динозавров, история этой группы заключает еще много пробелов, и то, что мы знаем, - лишь фрагменты этой истории. Это объясняется разными причинами и прежде всего - "неполнотой геологической летописи", т. е. незначительным, как правило, процентом захоронения всех живших видов. При этом наше представление о численности видового состава вымершей фауны нарушается еще условиями обитания и особенностями захоронения в зависимости от группы животных. Например, около половины всех находок по динозаврам, если иметь в виду число костей, составляют утконосые динозавры, но не потому, что их было так много, а потому, что они жили в водоемах, где и происходило их захоронение. Число водных животных, попадающих в захоронение, во много раз превышает число сухопутных животных, ибо из последних посмертно в водоем попадают лишь отдельные экземпляры, за исключением случаев массовой гибели при наводнении или других стихийных бедствиях. Захоронение же крупных животных вне водной среды, где происходит отложение осадков, практически немыслимо. Только мелкие животные (птички, насекомые и др.) могут быть погребены, например, песками во время сильной песчаной бури. Далее надо добавить, что не все геологические отложения одинаково вскрыты и. тем более не все одинаково изучены. Раскопки местонахождений ископаемых животных зависят от многих факторов, начиная с геологических и кончая экономическими и политическими, и носят весьма неравномерный, выборочный характер.
В истории динозавровых фаун четко выделяются три этапа, о которых нам более всего известно,- поздний триас, поздняя гора и поздний мел, тогда как о динозаврах промежуточных геологических эпох имеются гораздо более скудные сведения.
Поздний триас - эпоха становления групп ящерота-зовых динозавров и появления птицетазовых. Крупные местонахождения позднего триаса известны в Южном Китае, Южной Африке, Южной Америке и Западной Европе, откуда описано довольно много видов хищных динозавров - целурозавров и карнозавров, а также прозауроподы. В Южной Африке указаны первые птицетазовые динозавры и недостоверные пока остатки зауропод.
А затем - до поздней юры, мы почти ничего не знаем о судьбе динозавров. Прозауроподы вымирают, а им на смену приходят многочисленные зауроподы, широко распространенные по всему свету. Их остатками переполнены континентальные свиты тендагуру в Восточной Африке и моррисон в Северной Америке (США). Позднеюрские зауроподы довольно многочисленны в местонахождениях Центрального Китая и Западной Европы, известны из Южной Америки и Австралии. Вместе с зауроподамы в местонахождениях встречаются многочисленные остатки крупных уже карнозавров - мегалозавров и давно ушедших от своих предков представителей птицетазовых динозавров - орнитопод (игуанодонтов) и стегозавров. Этот динозавровый комплекс переходит и в ранний мел, но богатые местонахождения (за исключением Берниссарского) отсутствуют, и число видов динозавров невелико, так же как и в первой половине позднего мела.
Последний этап приходится на вторую половину позднего мела, когда континентальные отложения Северной Америки (США и Канада) и Центральной Азии вновь оказываются переполненными остатками динозавров из различных групп. Зауроподы к этому времени уже редки, стегозавры и игуанодонты исчезают так же, как и мегалозавры, но вместо них появляются многочисленные анкилозавры и цератопсы, утконосые динозавры и гигантские карнозавры. Этот фаунистический комплекс существует до конца мелового периода (и мезозоя, следовательно). С его исчезновением завершается история динозавров. Последние их представители вымирают на границе мезозойской и кайнозойской эр.
В Северной Америке есть местонахождения, где костеносные слои с остатками позднемеловых (последних) динозавров непосредственно сменяются слоями, заключающими остатки уже палеоценовых млекопитающих. Здесь и проводится граница между мезозоем и кайнозоем в отложениях, которой соответствует граница между этими двумя подразделениями во времени, и постановка вопроса: "Почему динозавры вымерли именно в конце мезозоя?" - неправильна по существу, поскольку как раз время, когда вымирают последние динозавры (а также морские рептилии и некоторые другие группы животных), принимается за границу между эрами. Необходимо подчеркнуть также, что некоторые группы динозавров вымерли значительно раньше. Так, прозауроподы вымерли еще в конце триаса, стегозавры - в раннем мелу, а отдельные семейства динозавров и тем более виды вымирали на протяжении всего мезозоя. Если судить по имеющимся в настоящее время находкам, то наибольшей численности динозавры достигали во второй половине позднемеловой эпохи, точнее, в камцанский и маастрихтский века, после чего их видовой состав начинает быстро сокращаться (не следует забывать, что один геологический век охватывает несколько миллионов лет). Поэтому, когда говорят о "катастрофе" в конце мезозоя, имея в виду вымирание динозавров и других крупных рептилий и неизбежно предполагая нри этом кратковременность такого явления, то это сильно преувеличено.
Каковы же все-таки причины вымирания динозавров? По этому поводу выдвинуто множество гипотез, нередко, к сожалению, недостаточно обоснованных и, как правило, ограничивающихся каким-либо единственным фактором. Рассмотрим некоторые из популярных гипотез.
До сих пор многие палеонтологи считают, что причина заключается в "конкуренции" - появившиеся в триасовый период млекопитающие как более высокоорганизованные в конце мезозоя якобы вытеснили динозавров. Но каким путем они могли это сделать? Немногочисленные и мелкие, величиной с крысу, млекопитающие мезозоя не смогли бы уничтожить динозавров ни физически, ни поедая их пищу, тем более что среди мезозойских млекопитающих неизвестны, например, водные формы. Более же высокий уровень млекопитающих, в первую очередь за счет развития центральной нервной системы и совершенства других органов, совсем не доказывает, что они могли истреблять более низко организованных животных.
Развитие центральной системы означает необходимость управления более сложным организмом, у которого усложняется процесс обмена веществ и увеличивается число функций. Все это определяется взаимодействием организма с окружающей средой. Совершенно закономерное усложнение живых существ представляет лишь естественный процесс развития материи от более простых форм к более сложным, причем простые формы не перестают существовать от появления более сложных. В настоящее время низко организованных беспозвоночных животных существует в тысячи раз больше, чем позвоночных. При этом любая муха или даже одноклеточная амеба "благоденствует" нисколько не хуже носорога или льва, и последние никак не угрожают первым. Исключение лишь одно - человек, мозг которого по сравнению с любыми животными достиг столь высокого совершенства, что он может сознательно направлять свою деятельность и в состоянии искусственно истребить не только любой вид животных и растений (что, к сожалению, он иногда и делает), но и все живое на Земле, включая и самого себя.
Обильные местонахождения скорлупы яиц динозавров на юге Франции дали повод высказать некоторым палеонтологам мысль о том, что это - последствия грандиозного побоища, которое было устроено динозаврам их прогрессивными соперниками - млекопитающими. Последние якобы были наделены особым инстинктом уничтожения динозавров (и в первую очередь гнезд с их яйцами) как своих исконных врагов, державших млекопитающих "в черном теле" на протяжении всего мезозоя. Такое заключение вряд ли может претендовать на научную достоверность.
У животных не существует инстинкта истребления другого вида. Вполне возможно, что некоторые млекопитающие могли питаться яйцами динозавров, притом вряд ли крупных. Но и в настоящее время яйцами рептилий питаются млекопитающие, птицы и сами рептилии, а рыбы поедают икру друг друга, однако это не приводит к вымиранию. Вообще в природе трудно указать пример физического истребления одного вида другим, так как существование видов складывалось исторически, и между ними всегда имеется равновесие, которое если и нарушается, то преимущественно другими факторами.
Возвращаясь к вопросу о вытеснении или истреблении мелкими млекопитающими гигантских динозавров, мы, скорее, должны прийти к противоположному выводу, а именно: динозавры и другие рептилии, широко распространенные в мезозое на суше, в воде и воздухе, тормозили развитие более высоко организованных, но малочисленных птиц и млекопитающих. Эти два высших класса позвоночных получили быстрое развитие лишь в начале кайнозойской эры - непосредственно после вымирания большинства групп мезозойских рептилий.
Самая новейшая и модная гипотеза - космическая. Авторы ее утверждают, что в конце мезозоя Земля подверглась интенсивному космическому облучению, приведшему к вымиранию динозавров и морских рептилий. Это предположение основывается на том, что кости динозавров оказались радиоактивными. Но при проверке выяснилось, что радиоактивность костей динозавров связана не с геологическим временем, а лишь с отдельными местонахождениями. И если, например, в том же местонахождении, но несколько выше по разрезу встречаются кости значительно более поздних млекопитающих, то они практически столь же радиоактивны, тогда как кости динозавров одного геологического возраста, но из разных местонахождений имеют и разную радиоактивность. Радиоактивность костей позвоночных (являющихся хорошим поглотителем) отмечается, начиная с палеозоя, в разные периоды и среди разных групп, но всегда связана с определенными местонахождениями. Это означает, что радиоактивность костей ископаемых животных носит вторичный характер и находится в прямой зависимости от радиоактивных рудных тел, расположенных поблизости. Невозможно было бы объяснить и избирательное действгте облучения только на динозавров и морских рептилий, и еще труднее объяснить этим вымирание их отдельных видов и групп на протяжении всего мезозоя.
Существуют и другие гипотезы (эпидемий, эволюционного старения групп, климатические, геологические и т. д.), но все они, как и рассмотренные, пытаются объяснить вымирание динозавров только каким-ниубдь одним фактором. В то же время напрашивается и более широкий вопрос: каковы причины вымирания других групп животных? Существует ли общность этих причин?
Основной показатель процветания любого вида животных - его многочисленность. А это в свою очередь означает, что он прежде всего обеспечен достаточной или даже избыточной кормовой базой и живет в благоприятных условиях, как географических (климат, ландшафт), так и биологических (малочисленность прямых и косвенных врагов). Все это находится во взаимосвязи и до поры до времени в равновесии. Виду не грозит вымирание. Но равновесие по законам, действующим в природе, не может быть постоянным, застывшим, ибо все развивается. Изменение климата, характера пищи и других условий обитания, несомненно, должно оказаться неблагоприятным если не для всех, то для каких-то животных, что будет приводить к изменению численности видов и иных соотношений между ними.
Животные, которые смогут приспособиться к жизни в новых условиях, сохранятся, несколько изменившись, т. е. превратятся в новые виды, а неприспособленные вымрут. Причины же изменения физико-географических условий (соотношение воды и суши, температура, характер ландшафта) кроются в непрерывных преобразованиях земной коры: со дна океанов поднимаются горные цепи, а последние, со временем разрушаясь, превращаются в равнины, затопляемые водами. Так, например, в настоящее время Скандинавия поднимается, а область Дальнего Востока испытывает погружение. Скорость вертикального движения земной коры, да и то в наиболее активных участках, не превышает одного метра в столетие, поэтому человеку трудно заметить существенные преобразования. Перераспределение воды и суши вызывает изменение климата в сторону континентальности или влажности, похолодания или потепления. Вместе с тем меняется растительный покров, меняются пищевые ресурсы сначала для растительноядных животных, а затем и для хищных, которые питаются растительноядными. Эти процессы совершались на протяжении всей истории Земли, вызывая изменения в животном мире - появление новых, более приспособленных видов и вымирание старых, менее приспособленных.
Какова же была обстановка в мезозое? К концу триасового периода закончилось разрушение большинства горных цепей, воздвигнутых в палеозое. Материки превратились в огромные равнины, на которые в следующий период - юрский - воды начали свое наступление. Климат стал более мягким и теплым, захватывая не только тропические и субтропические зоны, но и области современных умеренных широт. Обилие воды на континентах (моря, огромные озера и большие реки) благоприятствовало развитию пышной флоры - важной кормовой базы. Рептилии, бывшие в конце триаса наиболее высокоорганизованной и достаточно распространенной группой наземных позвоночных (к этому времени вымирает большинство групп первых наземных позвоночных - амфибий), оказались в весьма благоприятных условиях для своего быстрого развития. В юрский период они освоили не только всю сушу, но распространились в моря и пресные водоемы, приспособились к жизни в воздухе. Это были самые многочисленные, крупные и активные животные того времени.
В меловой период обстановка заметно изменилась. Последовал очередной цикл интенсивного горообразования, начавшийся кое-где даже в юрский период. Особенно заметны были его проявления в Америке и Азии - местах наибольшего распространения меловых динозавров. Поднятия горных систем вызывали осушение материков, изменение климата в направлении континенталь-ности. Это не могло не отразиться отрицательно на теплолюбивых рептилиях, какими были динозавры. К тому же большинство из них было прямо или косвенно связано с водоемами или с прибрежными районами. Теперь же все это нарушилось. С сокращением водоемов исчезла и пышная растительность, произраставшая здесь. Иными словами, природа "ликвидировала" естественные зоны обитания динозавров. Для таких крупных животных, как динозавры, заметное уменьшение кормовой базы должно было сказаться отрицательно и привести к уменьшению их численности и видового состава. Вымирание растительноядных форм должно было повлечь и вымирание хищных динозавров.
Почему же уцелели крокодилы, жившие одновременно и рядом с динозаврами? Хищники должны вымирать последними. После того как хищные динозавры доели своих растительноядных собратьев, которым не хватало пищи и негде было укрыться, они обрекли и самих себя. Крокодилы же, даже в сократившихся водоемах, могли найти себе пищу как за счет динозавровой молоди (утко-носых динозавров и зауропод), так и рыбы. Наконец, крокодилы могли свободно перебираться из одного водоема в другой или же пережидать засушливый период, зарывшись глубоко в ил. Таким образом, крокодилы оказались в более благоприятных условиях. Выжили и черепахи как наименее притязательные в смысле пищи. Ящерицам и змеям, немногочисленным еще в меловой период, также нетрудно было найти себе пропитание.
С вымиранием динозавров, в течение 150 миллионов лет господствовавших на всех континентах, остальные группы рептилий - крокодилы, черепахи, змеи и ящерицы - достигли в кайнозое широкого распространения и сохранились доныне.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'