
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Глава тринадцатая. Амфибии и рептилии
Амфибии
Выход амфибий на сушу. В предыдущей главе мы остановились на том, что первые амфибии, только что развившиеся из кистеперых рыб, которые выползли на сушу, стали знакомиться с сушей, которую им предстояло заселить. Конечно, обзор у них был еще небольшой, так как на своих слабых и все еще очень коротких ногах они были вынуждены ползать по земле, почти как рыбы, оказавшиеся на берегу. Амфибии могли передвигаться только медленно и неуклюже, но по крайней мере на суше у них не было врагов опаснее насекомых, которые, как можно предполагать, их практически не тревожили.
Но даже при отсутствии опасности со стороны хищников или серьезных конкурентов любые условия природной среды на суше представляют гораздо больше трудностей для жизни ее обитателей, чем водная среда (в море, озере или реке). В море условия жизни сравнительно однородны. Температура, освещенность и количество пищи изменяются относительно мало. Поэтому морские позвоночные в условиях стабильной среды изменялись меньше, чем наземные позвоночные. На суше животные подвергаются непосредственному воздействию гораздо более многочисленных и более действенных факторов, из которых достаточно назвать чередование тепла и холода, палящего солнца и дождя, а также внезапные ураганы, наводнения и пожары.
В этой новой среде позднедевонские амфибии, только недавно развившиеся из рыб, были поставлены в такие же условия, как потерпевший кораблекрушение Робинзон Крузо, неожиданно попавший в совершенно новую, незнакомую обстановку. Робинзон нашел или, проявив немалую изобретательность, создал приспособления, которые помогли ему справиться со всеми трудностями и выжить. Другими словами, он приспособился к новой суровой жизни.
Оказавшись на суше, амфибии приспособились к некоторым свойствам своей новой среды. Возможно, они "пошли" в определенной степени на компромисс, ограничившись в своем расселении такими областями, где эти новые свойства проявлялись слабее всего. С точки зрения истории эволюции их реакция была совершенно нормальной. Организм остается на месте до тех пор, пока он не вытесняется изменившимися условиями. Затем, если организм вообще способен эволюционировать, то он изменяется ровно настолько, чтобы прийти в соответствие с новыми условиями среды.
Очевидно, такая эволюция, содержащая элементы компромисса, дала возможность амфибиям преодолеть опасности жизни на суше, поскольку, как можно судить по их ископаемым остаткам, они продолжали существовать и образовывать новые виды в течение всего палеозоя. По крайней мере 70 миллионов лет они были, бесспорно, господствующими во всем мире наземными животными. Было найдено больше ста видов ископаемых палеозойских амфибий. Такое большое число означает, что животные приспособились ко многим условиям жизни вблизи воды (фото 38). Некоторые из них жили вдоль берегов рек, как современные лягушки, другие - в самой воде. Одни амфибии напоминали ящериц, другие - саламандр, а третьи совсем не имели ног и походили на змей. Многие были похожи на короткохвостых крокодилов, но это сходство было чисто внешним, так как крокодилы относятся к рептилиям - более специализированной огруппе позвоночных. Размеры амфибий колебались в больших пределах; длина их тела составляла от нескольких сантиметров до трех метров. Многие из них, возможно, имели чешуйчатую или гладкую кожу; другие, подобно своим двоякодышащим предшественникам, имели на голове защитный покров из костяных пластинок. Вероятно, в большинстве своем они были плотоядными и поедали насекомых, червей или небольших рыб.
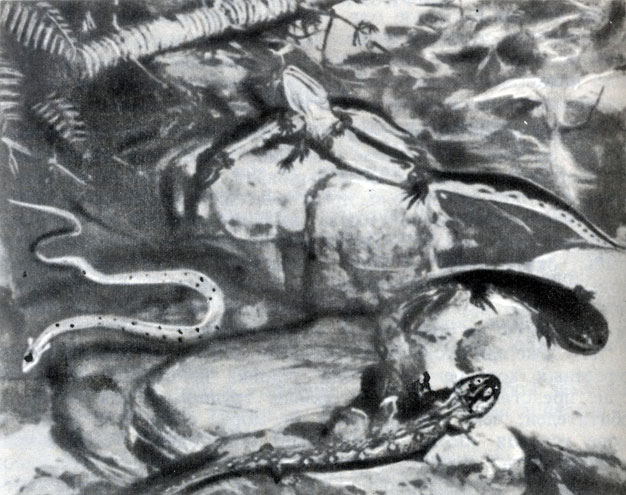
Фото 38. Четыре типа амфибий, живших в протоке с чистой водой в одном из болот каменноугольного периода. Рисунок кожи амфибий, который не сохранился у ископаемых, показан предположительно по аналогии с ныне живущими амфибиями. Реконструкция
Изменения в процессе эволюции. Изучая многие виды ископаемых палеозойских амфибий, мы можем выделить несколько основных изменений в анатомии амфибий. Эти изменения произошли в течение первых десятков миллионов лет после выхода амфибий на сушу. Небесполезно составить перечень этих изменений, так как некоторые из них сыграли роль в истории развития человеческого скелета.
1. Развитие шарового сустава конечностей. Плавник рыбы представляет собой лопасть. Чтобы обеспечить движение в воде, ему достаточно колебаться только вперед и назад относительно остального скелета. Но в наземных условиях необходима также способность двигаться в стороны, а это требует большей гибкости в бедренном и плечевом сочленениях конечностей. Такая необходимость возникла благодаря окружающим условиям, и в результате естественного отбора произошло развитие универсальных шаровых суставов. Эти суставы оказались настолько полезными, что они были унаследованы от амфибий более поздними наземными позвоночными и в результате стали непременной принадлежностью человеческого скелета.
2. Развитие конечности с пятью пальцами. В плавнике кистеперой рыбы уже были кости "запястья" и "кисти", но они имели продолжение в виде тонкой перепонки, усиленной большим количеством тонких поддерживающих костей - ребер; вся эта часть конечности напоминала часть конструкции зонтика с натянутой тканью и спицами. По мере развития рыб "запястье", "кисть" и их продолжение постепенно приобрели компактную закругленную форму (рис. 42), число ребер уменьшилось до пяти, а перепонка исчезла. В результате образовалась пятипалая конечность, очень полезное приспособление для движения по неровной поверхности. Амфибии сохранили эту особенность и передали ее своим потомкам.
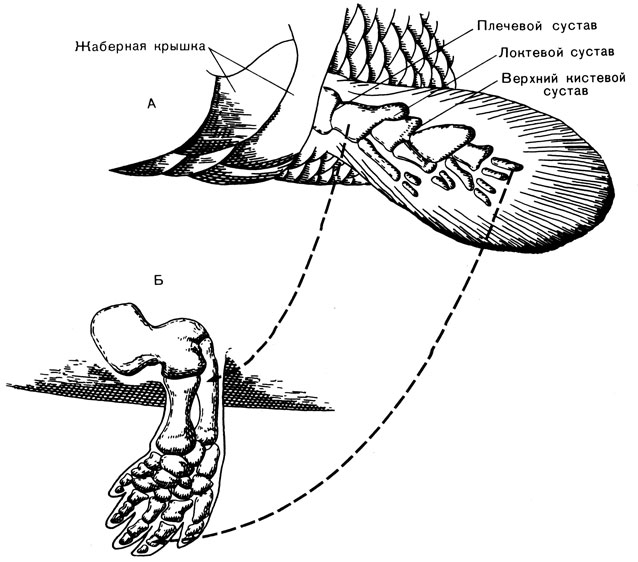
Рис. 42. Превращение плавников рыб в пятипалую конечность (не в масштабе). А. левый передний плавник девонской кистеперой рыбы. Б. Левая передняя конечность палеозойской амфибии. (Пунктирными линиями показаны соответствующие друг другу кости на рис. А и Б.)
3. Удлинение конечностей. Когда кистеперые рыбы стали впервые использовать свои плавники для опоры или придания телу устойчивости, они не могли приподняться над поверхностью ила. Всякое движение у них носило характер ползания. Напротив, амфибии, хотя их конечности все еще были короткими и малоэффективными, могли удерживать свое тело, хотя бы какое-то время, приподнятым над поверхностью земли, так что ползание превратилось в ковыляние. И это было началом процесса ходьбы.
4. Потеря панциря. Панцирь из костяных пластинок, который покрывал голову кистеперых рыб, очевидно, был необходим для защиты от хищников в воде. Но после выхода на сушу, где в те времена еще не было хищников, эта необходимость отпала. Панцирь стал бесполезной и обременительной тяжестью, как солдатские доспехи в Европе XVI в. после того, как вошло в употребление огнестрельное оружие. В результате эволюционного процесса панцирь исчез. Одна за другой группы животных лишались панциря, поскольку он был уже не нужен в окружающих условиях.
5. Увеличение размеров мозга. Процесс увеличения объема черепной коробки (полости в черепе, содержащей мозг) ясно прослеживается по черепам ископаемых амфибий. Но даже если бы такие ископаемые свидетельства отсутствовали, мы бы все равно были уверены, что мозг амфибий должен был вскоре превзойти по размерам мозг двоякодышащих рыб. Поскольку наземное животное подвергается в гораздо большей мере воздействиям окружающей среды по сравнению с животными, обитающими в воде, то условия суши в сильной степени вынуждают его развивать свои умственные способности, с тем чтобы оно могло соответственно реагировать на внешние факторы. Не обладая более совершенным механизмом реакций, амфибии не смогли бы выжить.
Упомянутые пять изменений были важны сами по себе. Но даже вместе взятые, они еще не могли скрыть очевидное сходство, существовавшее между ископаемыми рыбами и амфибиями. Взгляните снова на рисунок 31, и вы увидите, насколько сильно это сходство.
Мы уверены,что, помимо пяти изменений, прослеживающихся в ископаемых остатках, за 50 миллионов (или около того) лет, которые протекли между позднедевонским и позднекаменно-угольным временем, амфибии усовершенствовались еще в одном направлении, хотя по ископаемым остаткам этого не видно. Должно было произойти развитие все еще довольно примитивного легкого в более крупный и более развитый орган дыхания. При более быстром передвижении, которое обеспечивали удлинившиеся ноги, требовалось больше кислорода. Улучшение снабжения кислородом было также важно, если животное на короткое время погружалось в воду, как это, должно быть, делала безногая змеевидная амфибия, которую мы видим на фото 38.
Несмотря на исторический шаг, который сделали амфибии, будучи еще кистеперыми рыбами, и в ходе которого они одержали двойную победу - начали дышать воздухом и двигаться по суше, - несмотря на то что они первыми перешли к внутреннему оплодотворению, дальше этого амфибии не продвинулись. Они не сумели перейти к такому способу размножения, который позволил бы им порвать с водной средой. Они навсегда остались привязанными к воде, потому что им приходилось возвращаться к ней, чтобы откладывать яйца. Яйца современных амфибий, мягкие и непрочные, как у рыб, должны откладываться в воде, где они могут быть оплодотворены, и оставаться там, пока из них не выведется молодь. На воздухе они быстро высыхают и не развиваются. После выхода из яиц молодь находится в личиночной фазе (как, например, современные головастики) и дышит с помощью жабр, как рыбы.
Из-за такой особенности процесса размножения амфибии занимают среди животных приблизительно такое же положение, как бессеменные в царстве растений. И тем и другим для размножения требуется вода. Поэтому амфибии никогда не могли заселить возвышенные и сухие территории и по сию пору стремятся оставаться в местах, где по крайней мере в некоторые сезоны есть вода, в которую они могут откладывать яйца.
Несмотря на все эти несовершенства в строении амфибий, мы не можем так просто сбросить их со счета, поскольку они занимают действительно важное место в эволюции живых существ. Амфибии не только первыми из позвоночных прочно обосновались на суше, но и внесли свой вклад в эволюционный процесс. Вклад этот состоял в развитии таких особенностей строения, которые сделали возможным появление рептилий, что в свою очередь вызвало новую волну усовершенствований. Подобно тому как один из видов девонских рыб "выработал" легкое, так и один из видов амфибий в позднекаменноугольное время "усовершенствовал" строение яйца и тем самым положил начало линии рептилий. Развитие такого яйца было большим достижением и стало еще одной вехой на пути эволюции животных. Сейчас амфибии занимают далеко не господствующее положение среди животных, но в течение по крайней мере 50 миллионов лет палеозоя они были доминирующей и развивающейся группой. Пятьдесят миллионов лет - относительно продолжительный период; он больше, чем тот позднейший отрезок времени, в течение которого все человеческие расы и обезьяноподобные предки человека заселили Землю. Это время было достаточным для многочисленных "проб и ошибок", которые привели к созданию амниотического яйца.
Рептилии
Амниотическое яйцо. Появление принципиально новой "конструкции" яйца (рис. 43) ознаменовало новую эпоху, поскольку это яйцо не обязательно было откладывать в водной среде. Оно было устроено таким образом, что имело запас воды внутри себя, хотя и откладывалось на суше. Подобно космическому кораблю, амниотическое яйцо содержало привычную "микросреду" и все необходимое для жизнеобеспечения внутри своей защитной оболочки. "Обитателем" яйца был эмбрион рептилии; таким образом, создав яйцо, амфибии положили начало эволюционной линии рептилий. В настоящее время подобного устройства яйца нет ни у одной амфибии, зато им обладают все рептилии, а также все птицы и два вида млекопитающих. Конструкция яйца была настолько удачной, что основные ее черты остались неизменными на протяжении 300 миллионов лет с момента его появления.
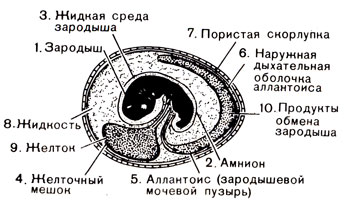
Рис. 43. Основные черты строения амниотического яйца
Помимо всего прочего, это яйцо имеет три оболочки, образующие три мешка. Первый, внутренний, мешок - амнион (от которого яйцо получило название амниотического) - содержит эмбрион, плавающий в жидкости. Жидкость и есть искусственная водная среда, заменяющая озерную или речную воду, в которой амфибии и рыбы откладывали яйца и в которой из яиц появлялись головастики, дышащие при помощи жабр, а позднее превращавшиеся во взрослую особь, дышащую воздухом.
Второй мешок содержит желток, большой запас пищи для эмбриона. Третий - зародышевый мочевой пузырь - связан с почками эмбриона и служит для сбора отходов - мочи эмбриона. Все три мешка заключены в твердую, но проницаемую оболочку; изнутри она выстлана другой оболочкой, позволяющей дышать находящемуся внутри яйца эмбриону. Эта оболочка, играющая роль легкого, обеспечивает проникновение внутрь яйца кислорода из окружающей среды и удаление из яйца углекислого газа. Остальное пространство внутри яйца заполнено жидкостью, которая разделяет все три мешка и играет роль гидравлического амортизатора, защищающего эмбрион.
Оплодотворение амниотического яйца было внутренним и происходило внутри тела самки. Яйца рыб оплодотворялись после того, как они отложены, и для того, чтобы выжить, рыбы должны были откладывать много яиц. Как и голосеменные растения, они должны были отличаться плодовитостью. Некоторые рыбы откладывают почти миллион икринок в каждый сезон размножения. Икринок погибает так много, что необходимо очень большое их количество, чтобы обеспечить выживание хотя бы немногих. Внутреннее оплодотворение привело к гораздо большей надежности размножения, потому что вероятность выживания отдельного оплодотворенного яйца значительно возросла.
Очевидно, что появление амниотического яйца явилось еще одним звеном в цепи усовершенствования способа размножения, начавшегося на ранних этапах существования биосферы с появлением полового размножения растений. Возникновение внутреннего оплодотворения и амниотического яйца было очень важным этапом, но на нем усовершенствование не закончилось.
Все меры предосторожности, обеспечивающие безопасность беспомощного эмбриона, покоящегося в своем мешке, как до, так и после откладывания яиц, выглядят так, как будто бы они были предварительно продуманы и спланированы, но это было не совсем так. По сути дела, это было ответной реакцией на воздействие окружающей среды, осуществлявшейся через естественный отбор в каждом новом поколении амфибий в течение очень долгого времени. Каким же именно было это воздействие? Вполне возможно, что основное влияние оказало постоянное увеличение в озерах и реках численности и разнообразия рыб и беспозвоночных хищников, охотно поедавших икру и небольших личинок - головастиков, беззащитную молодь амфибий. Создавшееся в результате этого положение не представляло опасности для взрослых особей, но ставило под угрозу выживание всего их сообщества. Появление любого приспособления, которое позволило бы яйцам постоянно находиться вне воды и дало бы возможность молоди пройти эту стадию личинок (когда они наиболее уязвимы) внутри яйца, а не в полной опасностей внешней среде, было бы величайшим достижением.
Сущность происшедшего изменения можно сформулировать двояко. Мы могли бы сказать: "Эта угроза, очевидно, заставила амфибий откладывать яйца в более безопасных местах, там, где их не могли достать хищники, а именно на берегах рек и озер выше уровня воды". С другой стороны, мы могли бы сделать хотя и менее эффектное, но зато более точное предположение о том, что небольшое преимущество с точки зрения возможности выживания имели те амфибии, у которых генетические изменения шли в сторону увеличения прочности оболочки яйца, содержавшего зачатки всех уже описанных нами составных частей. Дальнейшее развитие и закрепление этих полезных изменений в ходе последовательной смены поколений было только вопросом времени. А времени для этого было вполне достаточно, как мы уже видели, рассматривая другие события в истории Земли. От появления первых известных нам амфибий и до первых рептилий, предположительно обладавших яйцом такого типа, прошло 50 миллионов лет.
Амниотическое яйцо неизбежно должно было возникнуть. Раньше или позже должно было появиться или оно, или другое подобное устройство, которое привело бы к тем же самым результатам. Поэтому вопрос состоял не в том, что именно должно появиться, а в том, когда это произойдет.
Хотя в любой день мы можем позавтракать современным амниотическим яйцом, до сих пор никто еще не находил ископаемых амниотических яиц в палеозойских слоях. Наиболее ранние их находки относятся к слоям мезозойского возраста. Но позднекаменноугольные и пермские слои изобилуют скелетами ископаемых рептилий, а все рептилии обладают яйцами амниотического типа. Более того, у нас есть доказательства постепенного перехода от амфибий к рептилиям, основанные на изучении не ископаемых яиц, а ископаемых скелетов. В этих скелетах можно видеть в разнообразных сочетаниях признаки как амфибий, так и рептилий. Все это, вместе взятое, позволяет установить факт перехода от амфибий к рептилиям (фото 39). И если нас спросят: "Что появилось раньше, рептилия или яйцо?" - то мы, основываясь на ископаемых доказательствах постепенного перехода, должны будем ответить: "Ни то ни другое; яйцо и рептилия развивались одновременно и постепенно, путем длинного ряда небольших последовательных изменений".

Фото 39. Сеймурия (Seymouria), наземное позвоночное животное из нижнепермских слоев, и его гнездо у подножия дерева. Реконструкция, В скелете этого животного (длиной 60 см) присутствуют в равной мере признаки рептилий и амфибий. Сначала предполагали, что это рептилия, но теперь считают, что более логично относить его к амфибиям. Но каково бы ни было положение сеймурии в классификации, это животное представляет собой превосходный пример промежуточной стадии эволюционного перехода одной группы в другую
Только что мы говорили о признаках амфибий и рептилий. Каково же принципиальное различие между этими группами животных? Если начать с различий, заметных по ископаемым скелетам, то мы должны обратить внимание на строение шеи и конечностей. Рептилии имеют настоящую шею с одним шаровым суставом; таким образом, их голова может свободно поворачиваться относительно позвоночника. Животное, которое может, поворачивая голову, смотреть в разные стороны, может быть более подвижным, быстрее ориентироваться как в поисках пищи, так и при бегстве от врагов. У первых рептилий врагов, конечно, не было. Но прошло немного времени, и враги появились из среды самих же рептилий. Рептилии начали поедать друг друга; что-то подобное этому мы видим на фото 42. Что же касается конечностей, то коротенькие и слабые конечности амфибий, развившиеся из рыбьих плавников, в процессе перехода стали сильнее и удлинились. Таким образом, они могли приподнять тело рептилий vнaд землей и дали им возможность двигаться быстрее.
Эти различия и некоторые другие устанавливаются по ископаемым скелетам. Кроме того, мы видим, что и мягкие части тел ныне живущих рептилий и амфибий существенно различаются. Прежде всего вместо проницаемой кожи амфибий рептилии обладают непроницаемым покровом, часто покрытым чешуйками или пластинками жесткого рогового вещества. Очевидно, эти изменения предназначались для того, чтобы уменьшить потерю воды на испарение через кожу, но это могло произойти только после значительного усовершенствования легких, потому что дыхание амфибий частично осуществлялось через их тонкую кожу.
Кроме того, мышечная система и кровообращение рептилий более совершенны, чем у амфибий; они имеют более развитое сердце-насос, обеспечивающий циркуляцию крови. Наконец, мозг рептилий, хотя все еще небольшой по размеру, уже обладает зачатками мозговых полушарий, этих крупных лобных долей мозга, обусловливающих умственный уровень животного. Как известно, эта часть мозга имеет большие размеры у млекопитающих и вообще отсутствует у амфибий; появление ее у рептилий, возможно, объясняется улучшением кровообращения. Однако, несмотря на все большие преимущества, рептилии не имели сколько-нибудь эффективного способа регулирования температуры тела. Температура их тела зависела от солнца: когда температура окружающей среды понижалась или избыточно повышалась, рептилии становились вялыми и малоактивными.
Последствия возникновения амниотического яйца. Возникновение нового, более совершенного яйца имело два следствия, очень важных для животных, которые обладали таким яйцом. Наличие амниотического яйца повысило вероятность выживания, а также дало возможность животным заселять новые более разнообразные типы территорий. Стало выживать большее число зародышей, поскольку изменилось место откладки яиц, а зародыши стали появляться из яиц уже более развитыми. Появилась возможность откладки яиц за пределами досягаемости различных хищников, уничтожавших яйца и молодь амфибий в воде. Кроме того, благодаря приспособлениям, имевшимся внутри яйца, эмбрион мог полностью завершить свое развитие и подготовиться к борьбе за существование к моменту выхода из яйца. Значительно увеличилось также потенциальное жизненное пространство наземных позвоночных. Причина этого, очевидно, заключалась в том, что рептилиям уже не нужно было возвращаться к воде каждый раз, как наступал сезон размножения, и они не должны были селиться вблизи водоемов. Ничто не препятствовало им заселять более сухие районы, где прежде не ступала нога позвоночного и где до тех пор у них не было конкурентов. В то время как рептилии расселялись, амфибии остались привязанными к рекам и озерам, где большинство их обитает и по сей день. Процесс отделения рептилий от их предков амфибий и разрыв с привычными прибрежными условиями обитания напоминает отделение амфибий от их предшественников - рыб и выход их из водной среды, в которой они раньше обитали совместно. Завоевание суши позвоночными животными продвинулось еще на один шаг.
Климат и эволюция рептилий. После того как амниотическое яйцо стало окончательно служить признаком более прогрессивной формы жизни, появился новый фактор, который, очевидно, способствовал дальнейшему развитию как самого яйца, так и рептилий. Сравнительное изучение пермских и верхнекаменноугольных слоев позволяет обнаружить между ними большие различия, указывающие на изменение природной среды. Климат стал холоднее, и огромные болота каменноугольного периода постепенно исчезли. Уровень поверхности, по крайней мере некоторых континентов, повысился, и образовались многочисленные высокогорные хребты. Причины таких изменений все еще не ясны, но, возможно, это связано с взаимодействием движущихся плит коры и столкновениями континентов.
Но какова бы ни была причина, следствием явилось то, что влажные районы - естественная среда амфибий - стали встречаться реже. Тем самым был положен конец расселению амфибий - процессу, который продолжался на протяжении всего каменноугольного периода и которому способствовало наличие обширных болотистых низменностей, расположенных почти на уровне моря. Для существовавших тогда многочисленных и разнообразных амфибий не оставалось места. Жизнь постепенно становилась трудной для животных, зависящих от воды. Но для животных, обладавших амниотическим яйцом, это изменение климата не было столь губительным. Только что появившиеся рептилии могли довольствоваться многочисленными болотами и реками. Однако, не обладая средствами для поддержания постоянной температуры тела, они были вынуждены избегать более холодных областей.
Ползающие рептилии. После того как с возникновением амниотического яйца появились рептилии, они полностью использовали преимущества, которые им предоставлял этот способ размножения. Рептилии быстро изменили форму своего тела в соответствии с разнообразными условиями среды обитания. Чтобы упростить наш рассказ, мы можем опустить детали и въщедитъ всего две группы - "консервативную" группу ползающих рептилий и видоизмененную группу, называемую динозаврами. Последние были настолько многочисленны и своеобразны, что мы должны посвятить им отдельную главу, а сейчас мы кратко остановимся на ползающих рептилиях. Говоря "ползающие", мы имеем в виду, что эти рептилии продолжали традицию, унаследованную от амфибий, - их удлиненное тело с короткими ногами находилось при движении у самой поверхности земли. По форме тела ползающие рептилии в какой-то мере напоминали крокодилов (фото 40). Многие из них вели сходный с крокодилами образ жизни; очевидно, они проводили много времени в реках и озерах, охотясь за рыбой. Развитие позднепалеозойских ползающих рептилий было прослежено по ископаемым остаткам в более молодых слоях; они постепенно разделились на две ветви, давшие начало современным крокодилам и ящерицам.

Фото 40. Rutiodon, один из многочисленных видов ползающих рептилий. Реконструкция. Наличие его ископаемых остатков в нижнетриасовых слоях Европы и Северной Америки позволяет предполагать, что в раннетриасовое время эти континенты, возможно, соединялись. На переднем плане - цикадовые растения, позади рептилий - хвощи. На заднем плане справа - папоротники, а за ними - хвойные растения. Реконструкция
Одна из форм ползающих рептилий, ископаемые остатки которой в большом количестве обнаружены в пермских слоях, имела характерное приспособление - бросающийся в глаза пару-сообразный вырост на спине (фото 41). У ископаемых скелетов этот "парус" состоит из длинных тонких костяных отростков, которые отходили вверх от спинного хребта. У живой рептилии эти отростки наверняка соединялись перепонкой, как пальцы утиной лапы. Измерения, проведенные на многих ископаемых скелетах, показали, что площадь этого выроста была пропорциональна объему тела рептилии. Очевидно, вырост был пронизан кровеносными сосудами, и поэтому при нагревании солнечными лучами он мог обеспечивать быстрое повышение температуры тела холоднокровной рептилии, его действие можно сравнить с действием автомобильного радиатора, охлаждающего воду, которая циркулирует по многочисленным трубкам. Первые, хотя и несовершенные устройства такого типа появились у двух различных родов ползающих рептилий. Один из них, диметродон, изображенный на фото 41, был хищником, насколько мы можем судить по строению его зубов. Но подобный же вырост появился и у совсем другой, травоядной рептилии. У нее он отличался тем, что на длинных отростках находились короткие поперечные выступы. Хотя это была несколько видоизмененная конструкция выроста, его назначение было, очевидно, таким же, как и у рептилий другого типа.

Фото 41. Диметродон (Dimetrodon), пермская ползающая рептилия длиной более трех метров, имевшая на спине удивительное приспособление для терморегуляции. На переднем плане хвощи; на заднем - папоротники и другие деревья. Реконструкция
Рептилии, подобные млекопитающим. Конец палеозойской эры ознаменовался быстрым изменением рельефа и климата. Поэтому неудивительно, что в это же время происходило и активное развитие рептилий, которые были тогда наиболее развитыми и доминирующими животными. Среди вновь образовавшихся к этому времени рептилий особенно важная роль принадлежит одной группе, потому что именно она впоследствии дала начало новой эволюционной ветви, называемой млекопитающими. Наиболее ранние свидетельства об этой группе содержатся в ископаемых остатках из позднекаменноугольных и пермских слоев, найденных в Южной Африке и Аргентине. Возможно, что эти две области представляли в пермское время единый массив суши, как это показано (в виде предположения) на рисунке 27. Если это так, то не удивительно, что на этих двух континентах, в настоящее время разделенных тысячами километров морского пространства, обнаружены ископаемые остатки одинаковых или очень схожих наземных животных.

Фото 42. Два вида рептилий, подобных млекопитающим, найденные в триасовых слоях Южной Африки. Три хищника (Cynognathus, слева) готовятся напасть на травоядную рептилию (Kannemeyeria) длиной около двух метров. Растительность представлена цикадовыми и хвойными. Реконструкция
Эти ископаемые остатки представлены скелетами рептилий, но рептилий, у которых уже начинают проявляться несомненные признаки млекопитающих; позднее в триасовых слоях появляется несколько родов таких рептилий, и они еще в большей степени похожи на млекопитающих. Отличительными их признаками, прослеживающимися по скелетам, являются длинные шеи, дифференцированные зубы и более развитый мозг. Предположительная реконструкция их облика дана на фото 42. Конечности удлинились, причем суставы на передних конечностях - "локтевые" - сместились назад, а на задних - "коленные" - вперед. Благодаря этому тело рептилии слегка приподнялось и стало не таким приземистым. Подобное положение тела мы видим у млекопитающих (рис. 44), у которых кости конечностей надежно поддерживают тело и освобождают мышцы от излишней нагрузки. Зубы, которые у рептилий были все одинаковыми и имели коническую форму, начинают подразделяться на резцы, клыки и коренные. Увеличился и развился мозг, особенно его лобные доли.
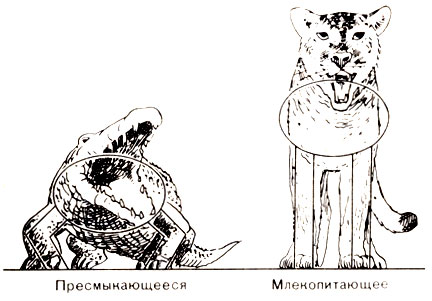
Рис. 44. Положение тела рептилии и млекопитающего
К другим признакам млекопитающих, которые не устанавливаются по скелетам, относятся:
1) молочные железы для вскармливания детенышей;
2) волосяной покров, защищающий тело от влияний внешней среды и препятствующий потере тепла;
3) теплая кровь;
4) четырехкамерное сердце - наиболее совершенный насос, предназначенный для снабжения мозга (и всего организма) кровью вне зависимости от колебаний внешней температуры.
Рептилии после захода солнца становятся холодными и вялыми, млекопитающие же сохраняют относительно высокую температуру тела. Возможно, жившие в триасе рептилии, подобные млекопитающим, обладали большей частью этих признаков, по крайней мере в зачаточной форме. Что касается размножения, то предполагается, что они откладывали яйца так же, как и наиболее примитивные из современных млекопитающих, но точно это не установлено. Однако мы можем с уверенностью утверждать, что развитию более длинных конечностей сопутствует усовершенствование мозга. Такие конечности позволяют уже не ползать, а передвигаться с большой скоростью, что требует значительно лучшей мышечной координации и вызывает необходимость в более сложном контролирующем устройстве. Любой конструктор самолетов может сказать, что, чем большее число выполняемых операций вы закладываете в конструкцию самолета, тем большим числом контролирующих приборов вам придется его оснастить.
Рептилии, подобные млекопитающим, были многочисленны и разнообразны. Некоторые из них превышали в длину три метра; другие были не больше мыши. И все они вымерли. Но перед тем, как исчезнуть, одна из них - вероятно плотоядная, изображенная на фото 42, - положила начало ветви настоящих млекопитающих, что явилось еще одним крупнейшим достижением эволюции.
Суммируя, мы можем наметить четыре пути, по которым шло эволюционное развитие рептилий (рис. 45). Первый - это наиболее консервативный путь ползающих рептилий, которые никак не проявили себя и навсегда остались ползающими. Второй путь - это путь динозавров, образовавших множество крайне своеобразных разновидностей и тем не менее прекративших свое существование до начала кайнозойской эры. Третий путь - это путь птиц, которые развились из рептилий незадолго до начала юрского периода. Четвертый, и последний, путь начался с рептилий, подобных млекопитающим. Он быстро привел к млекопитающим и в кайнозое разделился на множество троп, одна из которых привела к человеку.
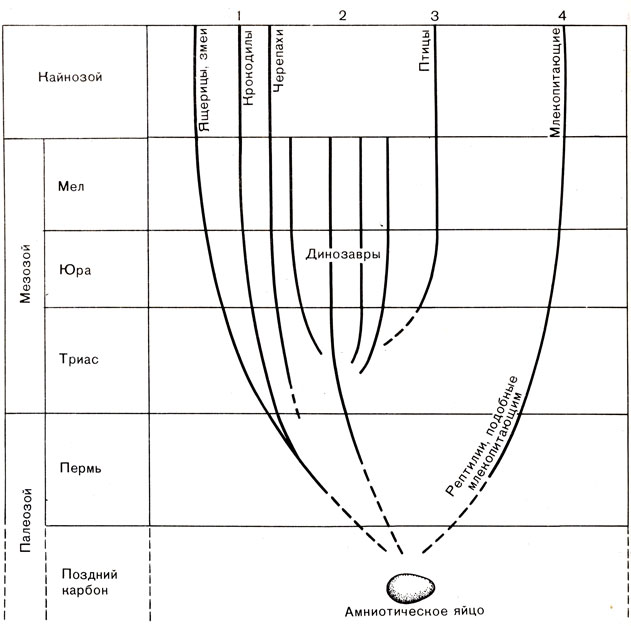
Рис. 45. Пути развития рептилий, ставшие возможными благодаря появлению амниотического яйца
На этих путях эволюции происходило много неожиданного. Одной из наиболее драматических была история динозавров, о которых мы расскажем в следующей главе.
Литература
Gregory W. К., 1951, Evolution emerging: The Macmillan Co., New York, vol. 1, p. 246-287.
Rome r A. S., 1964, The vertebrate story: 4th ed., University of Chicago Press, p. 87-175
Smith H. W., 1961, From fish to philosopher: American Museum of Natural History, New York, p. 85-101,120-121. (Paperback.)
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'