
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Глава четырнадцатая. Век динозавров
Рептилии на суше
За палеозойской эрой последовал один из самых замечательных периодов в истории жизни на Земле - царствование мезозойских рептилий. В пределах мезозоя, на протяжении 190 миллионов лет, произошло поразительное распространение рептилий. Рептилии, появившиеся в результате эволюции в позднекаменноугольное время, благодаря преимуществам размножения с помощью амниотического яйца распространились на суше, заселили моря и поднялись в воздух, используя только что появившиеся крылья. Одна из ветвей рептилий дала начало птицам, которые конкурировали с самими летающими рептилиями. Другая ветвь, как мы уже видели, развилась в ветвь млекопитающих. И все же наиболее яркие персонажи в драме рептилий - динозавры. Они и все их родичи, плавающие и летающие, вымерли в мезозойскую эру. Вымерли полностью, до последней особи, предоставив заселять Землю новым группам животных, главным образом потомкам мезозойских млекопитающих.
Летопись ископаемых свидетельствует о существовании сотен родов динозавров размерами от цыпленка до десятков метров, имевших десятки самых разнообразных приспособлений к жизни в различных условиях. И все же, как и все другие рептилии, все динозавры, вероятно, были холоднокровными и прямо или косвенно зависели от наличия в большом количестве лиственной растительности. Однако ископаемые динозавры были найдены на всех континентах, кроме Антарктиды. Из этого обстоятельства мы можем заключить, что в мезозойское время были широко распространены низменности с мягким климатом и пышной растительностью, так как животные, подобные динозаврам, не могли существовать в высоких горах с крутыми склонами и в холодном климате. Этот вывод, по-видимому, согласуется с имеющимися у нас данными о перемещениях плит земной коры. Взглянув еще раз на рисунок 27, мы увидим, что большая часть территории суши, которая сейчас располагается в средних или высоких широтах, в середине мезозоя находилась (предположительно) в более низких широтах. Не исключено, что южная часть Северной Америки и южная часть Европы тогда примыкали к экватору. Если бы континенты занимали в мезозое то же положение, что и сейчас, то маловероятно, чтобы рептилии были столь многочисленны и чтобы достигали таких огромных размеров.
С помощью карты, представленной на рисунке 38, мы можем объяснить расцвет мезозойских рептилий с другой точки зрения. К концу мезозоя территория Северной Америки по сравнению с настоящим временем в большей степени была занята мелководными морями и в меньшей степени - сушей, в большинстве своем низменной, а Мексиканский залив соединялся с Северным Ледовитым океаном. При этих условиях климат центральной и северной частей континента, возможно, был более мягким, чем сейчас, особенно зимой. И в Европе и на других континентах в мезозое были распространены обширные моря.
Таким образом, расцвет рептилий в мезозойскую эпоху, который на первый взгляд кажется необъяснимым, в конце концов удовлетворительно объясняется наличием окружающих условий, которые были благоприятны для холоднокровных животных. Таким образом, как и при рассмотрении более ранних этапов истории живых существ, мы еще раз убеждаемся, что условия окружающей среды оказывают определяющее влияние на развитие животного мира путем естественного отбора.
Типы динозавров
Мы уже говорили о том, что динозавров существовало несколько сот видов. Но все они принадлежали к двум четко выраженным подразделениям, которые произошли от одного общего предка в триасовое время, прежде чем появились динозавры как таковые. Название "динозавр" является скорее популярным, чем научным. Оно означает "страшный ящер", и когда его впервые ввели, оно относилось к очень крупным и свирепым животным. Но динозавров такого типа было относительно немного среди тех многочисленных рептилий, которых мы в настоящее время относим к динозаврам. В эту группу входит большое число рептилий, не обладавших ни свирепостью, ни большими размерами.
Упоминавшиеся основные два подразделения динозавров ученые различают на основании строения их тазовых костей. К одному относились динозавры, у которых тазовые кости имели такое же строение, как у ящериц, а ко второму - динозавры, тазовые кости которых напоминали птичьи. Это важное конструктивное различие в устройстве показано на рисунке 46. Нет необходимости останавливаться на этом более подробно, нас интересует в первую очередь внешний облик и образ жизни динозавров. Поэтому мы можем перейти к описанию отдельных наиболее выдающихся представителей мира динозавров. Триасовые динозавры были довольно примитивными и имели скромные размеры. Все они опирались на задние ноги, а передние, гораздо меньшие по размерам, до земли не доставали (рис. 47). Шеи у них были гораздо длиннее, чем у пермских ползающих рептилий. Однако, хотя динозавры и стали двуногими, они не были прямо стоящими, как опирающийся на две ноги человек. Когда они ходили или бегали, тело у них занимало положение ближе к горизонтальному, чем к вертикальному, хотя, несомненно, они иногда могли выпрямляться, как это зачастую делают белки. Что касается ног динозавров, то, посмотрев на оставленные ими на влажном песке и иле следы (фото 18), на которых отчетливо видны отпечатки трех или четырех длинных пальцев и еще одного короткого, дополнительного, лишь изредка касавшегося земли, мы можем понять, почему первые исследователи этих следов принимали их за следы птиц.
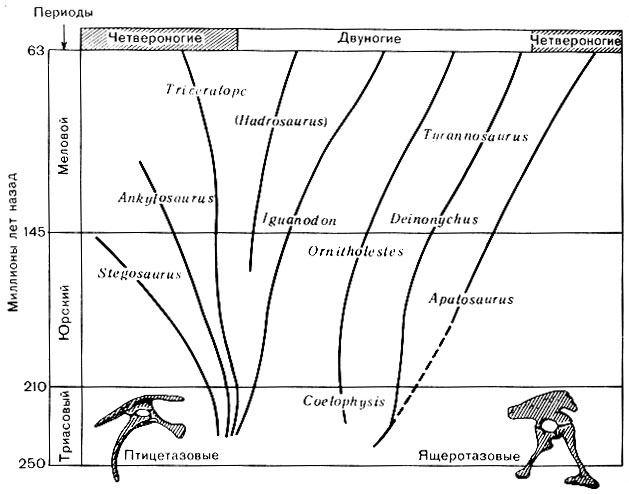
Рис. 46. Связь между группами динозавров, упомянутыми в книге
Большинство динозавров было хищниками, как и их пермские предки; присутствие среди триасовых ископаемых разновидностей, обладавших необычного вида броней, выростами и шипами, говорит о том, что они уже начинали "принимать" защитные меры против своих врагов - других хищных динозавров.
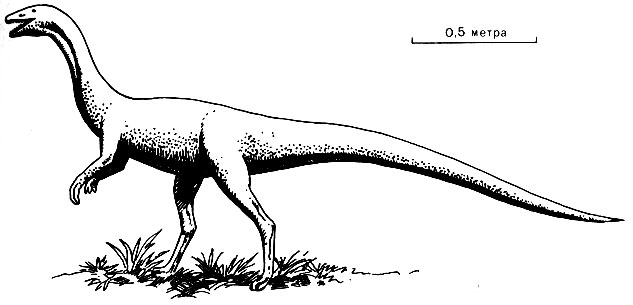
Рис. 47. Coelophysis, типичный динозавр триасового времени. Весьма вероятно, что мелкие следы, показанные на фото 17, оставлены именно этим динозавром
Естественно, эта группа довольно примитивных триасовых динозавров включала предков всех более поздних динозавров. Лучше всего подразделять их по способу питания, образу жизни и особенностям строения. Мы можем выделить растительноядных и плотоядных, двуногих и четвероногих динозавров, а также динозавров, имевших броню, костяные пластины или защитные рога, и таких, кто не имел этих приспособлений. Мы разделим рассматриваемых нами ящеров на четыре большие группы.
Растительноядные двуногие. Хотя почти все раннемезозойские динозавры были хищниками, среди их потомков встречалось много растительноядных особей. Судя по оставленным ими следам, они довольно часто передвигались на четырех ногах. Среди них обычным был игуанодон (рис. 48), животное плотного сложения, достигавшее около 11 метров в длину. В одном месте было найдено более 20 скелетов в полной сохранности, по найденным совместно с ними скелетам черепах, крокодилов и рыб можно думать, что эти динозавры жили в болотах. "Руки" у них имели пять пальцев, причем "большой" палец представлял собой большой острый шип, вероятно служивший неплохим орудием защиты. Очевидно, эти ящеры питались, пригибая передними конечностями ветви деревьев и объедая побеги. Их следы показывают, что они перемещались шагом и, вероятно, не очень быстро, лишь временами делая короткие прыжки.
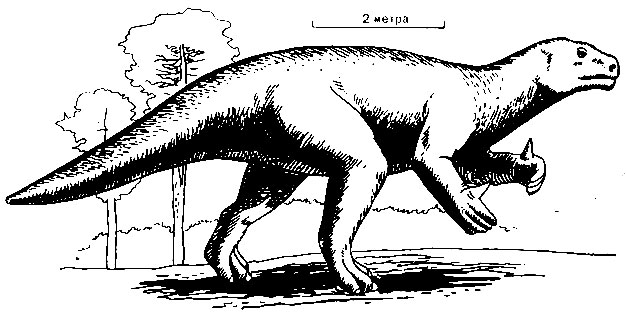
Рис. 48. Игуанодон, крупный двуногий травоядный динозавр, обитавший в Европе
Другая группа растительноядных двуногих ящеров, достигавших 6-12 метров в длину и носивших название гадрозавров, по образу жизни напоминала амфибий и обитала в болотах или на их заболоченных берегах (фото 43). Между пальцами ног у них имелись небольшие перепонки, а хвост был тонким, как у крокодилов, и при движении в воде действовал, как весло. Ноздри были расположены так, чтобы почти все тело могло быть погружено в воду. Рот состоял из рогового клюва, подобного утиному. На челюсти находилось до тысячи зубов, длинных, очень тонких, расположенных вплотную друг к другу. Когда роговой клюв доставал мягкие растения из болота, верхняя и нижняя челюсти, на которых росли зубы, начинали двигаться вперед и назад и тереться друг о друга, как две проволочные щетки, перетирая таким образом пищу.

Фото 43. Гадрозавры (1), 'бронированный' динозавр, подобный анкилозавру, (2) и плотоядный динозавр Struthiomimus (3). Дерево слева относится к покрытосеменным. Реконструкция
Плотоядные двуногие. Там, где встречаются растительноядные животные, всегда есть и хищники, которые на них охотятся. Среди динозавров было множество бегающих на двух ногах хищников различного размера и облика. Один из них, Ornitholestes, длиной всего около двух метров, имел настолько "изящное" строение, что весил предположительно менее 25 килограммов. Это было подвижное животное, приспособленное к быстрому бегу; хватательные передние конечности с тремя очень длинными пальцами могли поймать даже очень мелкую ящерицу, которая пыталась спастись бегством. Другой динозавр, Struthiomimus (фото 43, под третьим номером), был немного больше и напоминал страуса. У него даже был беззубый клюв. Разбитый череп родственного ему динозавра был найден в ископаемом гнезде, содержавшем яйца динозавров. Это обстоятельство, а также общий облик животного, имевшего небольшой вес и гибкие "руки", приводит нас к выводу о том, что Struthiomimus питался яйцами и грабил гнезда.
Еще один динозавр, Deinonychus, длиной около 2,5 метров, который, возможно, был потомком динозавра Ornitholestes, отличался двумя очень интересными приспособлениями, позволявшими ему вести хищный образ жизни. Второй палец на каждой задней ноге был снабжен когтем, гораздо более длинным и острым, чем все остальные когти. Этот палец имел особый сустав, позволявший ему приподниматься над землей и поворачиваться на 180° (рис. 49), что позволяло рептилии наносить своей добыче сильный удар ногой, удар, который мог распороть брюхо животному такого же размера, как и сам хищник. Кроме того, в длинном хвосте этого динозавра находились сухожилия, которые могли мгновенно "сращивать" кости вместе, превращая хвост в жесткий противовес всему телу. Обладая подобными когтями и хвостом, такой динозавр; должен был быть очень подвижным и опасным животным.
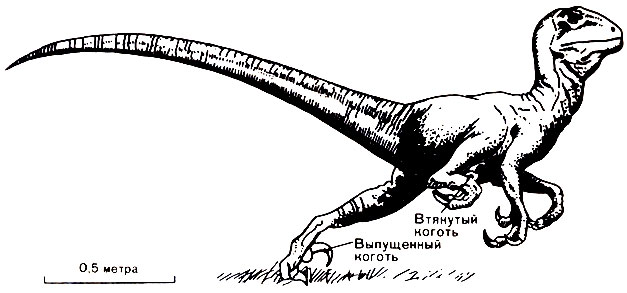
Рис. 49. Deinonychus, хищник, вооруженный острыми когтями
Некоторые двуногие хищники имели гораздо большие размеры, превосходя 9 метров в длину. Один из них, тираннозавр, был самым большим из известных наземных хищников; он имел длину до 15 метров, высоту до 6 метров и предположительно весил 7-8 тонн (фото 44). Длина его черепа составляла 1-2 метра, а во рту находилось множество острых зазубренных зубов пятнадцатисантиметровой длины. Так как его передние конечности были очень короткими, очевидно, он не пользовался ими, когда нападал на добычу и поедал ее. Главной добычей тираннозавра были растительноядные динозавры, как, например, гадрозавры и динозавры, вооруженные рогами.

Фото 44. Тираннозавр, самый крупный хищник, нападает на трицератопса, который приготовился к защите. Голова трицератопса покрыта панцирным шлемом. Деревья представляют собой пальмы, относящиеся 'к покрытосеменным растениям. Реконструкция
Земноводные четвероногие. Перейдем к гигантским динозаврам, которые так часто описываются в популярной литературе, что их облик знаком не только ученым. В палеонтологической летописи имеются данные по крайней мере о четырех различных родах, внешне очень похожих; мы упомянем только двух из них. На первый взгляд может показаться странным, что, хотя динозавры из этих двух родов были четвероногими, передние ноги у них были гораздо короче задних. Но в действительности этого можно было ожидать, потому что они были потомками триасовых двуногих динозавров с короткими передними конечностями. Пожалуй, лучше всего известен род Apatosaurus (фото 45) - огромные малоподвижные травоядные, достигавшие 23 метров в длину; их короткое туловище поддерживалось массивными колоннообразными ногами, снабженными когтями. Впереди находилась длинная гибкая шея с маленькой головой, которую в задней части туловища уравновешивал длинный гибкий хвост, сужающийся к концу. Животное должно было весить более 30 тонн, то есть вчетверо или впятеро больше самого большого африканского слона.

Фото 45. Apatosaurus, четвероногий динозавр, подобный амфибиям, длиной более 20 м, на берегу юрского водоема. Два других таких же динозавра пасутся в воде. По сравнению с ними крокодил на переднем плане выглядит совсем маленьким. Растительность состоит из цикадовых и хвощей. Реконструкция
По мере того как возрастали размеры и вес предков этого динозавра, происходила эволюция скелета в сторону снижения его веса путем образования полостей и отверстий в позвонках; тем самым вес снижался там, где нагрузки были невелики, и сохранялся там, где была важна прочность, например в колонновидных ногах. Отпечаток ноги этого динозавра, оставленный в мезозойских ил ах, превышает 90 сантиметров в длину.
Другой гигантский динозавр, диплодок, также был травоядным, во многих отношениях подобным описанному выше. Основное отличие от него состояло в том, что диплодок был несколько длиннее (длина одного экземпляра, согласно расчетам, превосходила 29 метров при высоте почти 14 метров), но не так массивен, вес его предположительно составлял 10-12тонн. Без сомнения, большую часть времени эти гиганты проводили в болотах и реках, поедая мягкие растения. Вдали от берега, среди болотистых островов, они были в большей безопасности от крупных хищников; поэтому такие места были для них не только "столовой", но и убежищем. Для еще большей безопасности ноздри этих гигантов помещались на самой верхней части головы, что позволяло им спокойно дышать, почти полностью погрузившись в воду и находясь таким образом вне поля зрения врагов. Эти и некоторые другие динозавры проглатывали свою растительную пищу целиком и перетирали ее уже после того, как она попадала к ним в желудок. Как цыплята, в зобу у которых обычно находится много камешков, динозавры заглатывали камни размером с картофелину и с помощью этих орудий своими сильными желудочными мышцами измельчали пищу. Иногда кучки таких камней, когда-то окатанных и отшлифованных в желудках динозавров, находят вместе с их скелетами, причем они располагаются там, где находилось брюхо большого динозавра.
Вероятно, эти огромные животные откладывали яйца, хотя это не подтверждено еще находками; яйца в воде погибали, поэтому они должны были откладывать их на суше, а может быть на островах или в других местах, куда трудно было проникнуть хищникам.
Огромные четвероногие динозавры, подобные амфибиям, обладали еще меньшим мозгом по отношению к весу тела, чем другие динозавры, хотя и вся эта группа не слишком славилась своими умственными способностями. У диплодока истинный мозг весил только около семи граммов на тонну веса тела. Мы говорим "истинный мозг", потому что диплодок, как и многие другие динозавры, имел дополнительный координирующий центр гораздо большего размера, расположенный в позвоночнике, вблизи таза. Этот центр соединялся с истинным мозгом через спинной и управлял движением задних ног и хвоста. Хотя такое устройство может показаться неудобным, мы должны признать, что оно действовало "исправно", потому что им обладали многие различные виды динозавров, жившие на протяжении десятков миллионов лет. Этому, конечно, способствовала среда обитания динозавров с мягким климатом и мало изменявшимися природными условиями; в подобной среде почти не возникало проблем, требующих умственного напряжения.
Четвероногие, снабженные броней или рогами. В наш перечень разнообразных динозавров входит также довольно разнородная группа видов, которые, не будучи близкородственными, обладали необычного вида броней, или рогами, или и тем и другим. Несмотря на то что их предки в триасовое время были двуногими, эти динозавры снова опустились на все четыре конечности. И все же их передние ноги все еще были короче задних, как у Apatosaurus. Будучи травоядными, они нуждались в защите от хищных рептилий; это и вызывало развитие брони и защитных рогов.
Наиболее выделяющимся из этих бронированных рептилий был стегозавр. На его скелете, имеющем в длину около 6 метров и вес предположительно 4 тонны, видны толстые треугольные костные пластинки, окаймляющие спинной хребет, с которым они, вероятно, соединялись связками. Возможно, эти пластинки, самые крупные из которых достигали размера 75 сантиметров, защищали позвоночник от двуногих хищников, которые, вероятно, при нападении пытались вцепиться в загривок, как это делает терьер, убивая крысу. Кроме того, стегозавр был вооружен парой крепких толстых шипов длиной около 60 сантиметров, расположенных на конце хвоста. Один удар такого хвоста мог, вероятно, сбить с ног довольно крупного противника и к тому же нанести ему серьезные повреждения.
Анкилозавр и его родственники (фото 43) имели, вероятно, такую защиту, какую имеют современные броненосцы. Достигая 6 метров в длину и 2,5 метра в ширину, они имели высоту менее 1,5 метра. Позади мощного, толстого, снабженного клювом черепа вся верхняя половина тела у них была покрыта тяжелыми костными пластинками. У некоторых из них вдоль всего тела, от плеч до хвоста, напоминавшего тяжелую лопатку или дубинку, тянулись также огромные шипы. Обладая такой защитной броней, эти рептилии двигались, вероятно, медленно. Но при приближении опасности они могли прижиматься к земле, поджав под себя лапы, и защищаться от нападения, нанося удары хвостом.
Другим способом, используя рога, защищался трицератопс и его многочисленные родственники (фото 44). Эти громоздкие короткохвостые четвероногие достигали 7,5 метра в длину и трех метров в высоту. Их наиболее характерной особенностью был огромный тяжелый череп, который протягивался назад в виде большого щита, защищавшего шею. Спереди череп был снабжен двумя рогами, выступавшими над узким клювом, похожим на клюв попугая. Внутри черепа находился мозг, небольшой, по нашему мнению, но достаточно крупный для динозавра. Наличие такого мозга позволяет предположить, что эти животные, обладавшие защитным шлемом и рогами, были довольно подвижны. Об этом свидетельствует незащищенность задней части их тела, не имевшей ни брони, ни какого-либо оружия. Совершенно ясно, что они могли быстро поворачиваться, чтобы отразить нападение врага рогами. Следами таких древних битв являются, возможно, шрамы, часто встречающиеся на ископаемых остатках шейной брони.
Рассуждая о сражениях между динозаврами, мы невольно задаемся вопросом, происходили ли они в тишине или сопровождались громкими воплями, как это происходит при драках современных кошек и собак. Специалисты по анатомии динозавров могут сообщить нам то немногое, что известно по этому вопросу. Представляется, что конфигурация маленьких косточек у основания языка динозавров близка к той, которую имеют те же косточки у некоторых ныне живущих видов животных. Основываясь на этой аналогии, можно предполагать, что по крайней мере некоторые динозавры могли издавать квакающие или лающие звуки, как это делают современные крокодилы. Поэтому, если в палеозое на суше скорее всего царило молчание, нарушавшееся лишь шумом ветра, потоков и прибоя, то мезозойские ландшафты уже могли быть оживлены звуками, которые издавали животные.
Протоцератопс, родственный трицератопсу, но менее сложно организованный небольшой динозавр, с клювом, но без рогов, живший в Азии, получил широкую известность в связи с находкой его яиц и гнезд палеонтологической экспедицией в Монголии в двадцатых годах нашего века. В позднем мезозое эта территория была такой же засушливой, как и сейчас, и яйца были отложены в небольшие углубления в песке, который в настоящее время превратился в песчаник. Самки динозавров выкапывали ямки и откладывали в них до 15 яиц длиной 15-20 сантиметров. Было найдено несколько таких гнезд, и по крайней мере в двух яйцах были обнаружены крошечные кости детенышей динозавров, которые не смогли вылупиться. Были найдены также яйца и других типов динозавров, как большего, так и меньшего размеров.
Морские рептилии
При изучении жизни в мезозое едва ли не сильнее всего поражает то, что почти половина всех известных видов рептилий жила не на суше, а в воде, в реках, эстуариях и даже в море. Мы уже отмечали, что в мезозое на континентах широко распространились мелководные моря, поэтому для водных животных не было недостатка в жизненном пространстве.
В мезозойских слоях встречается большое количество ископаемых рептилий, приспособленных для жизни в воде. Этот факт может означать только то, что некоторые рептилии вернулись обратно в море, на родину, где когда-то давно появились предки динозавров - рыбы. Этот факт требует некоторого объяснения, поскольку на первый взгляд здесь происходил регресс. Но мы не можем считать возвращение рептилий в море шагом назад с эволюционной точки зрения только на том основании, что девонские рыбы вышли из моря на сушу и развились в рептилий, пройдя стадию амфибий. Напротив, это положение иллюстрирует принцип, согласно которому каждая активно развивающаяся группа организмов стремится занять все разновидности среды, в которых она может существовать. В действительности перемещение рептилий в море не слишком отличается от заселения амфибиями рек и озер в позднекамен-ноугольное время (фото 38). В воде имелась пища и конкуренция была не слишком жестокой, поэтому сначала амфибии, а затем рептилии переселились в воду. Уже перед концом палеозоя некоторые рептилии стали водными обитателями и начали приспосабливаться к новому образу жизни. Эта адаптация шла в основном по пути усовершенствования способа передвижения в водной среде. Конечно, рептилии продолжали дышать воздухом так же, как дышит воздухом современный кит, млекопитающее, хотя и сходное по форме тела с рыбой. Более того, мезозойские морские рептилии развились не из какой-либо одной наземной рептилии, которая приняла решение уйти обратно в воду. Ископаемые скелеты неоспоримо свидетельствуют о том, что они имели различных предков и появились в различное время. Тем самым ископаемые остатки показывают, насколько разнообразным был отклик организмов на изменение условий среды, в результате которого создалось обширное пространство, изобилующее пищей и пригодное для заселения.
Обширная информация была получена при изучении ископаемых остатков, содержащихся в морских аргиллитах и меловых известняках; в этих тонкообломочных породах сохраняются не только кости, но также отпечатки кожи и чешуи. За исключением наиболее мелких и примитивных видов, большинство морских рептилий было хищниками и принадлежало к трем главный группам: их-тиозавры, плезиозавры и мозазавры. Кратко характеризуя их, мы должны в первую очередь отметить, что ихтиозавры приобрели сходную с рыбами удлиненную форму (рис. 50) и были превосходно приспособлены к быстрому плаванию в погоне за рыбой или головоногими моллюсками. Эти животные, достигавшие 9 метров в длину, имели голую кожу, спинной плавник и хвост, как у рыбы, а их четыре конечности превратились в подобие тюленьих ласт и использовались для управления движением тела при плавании. Все пальцы в этих ластах были тесно соединены, и для увеличения прочности в них существовали дополнительные кости. Большие глаза ихтиозавров были приспособлены для того, чтобы хорошо видеть в воде. У них даже появилось одно очень существенное усовершенствование процесса размножения. Будучи животными, которые дышали воздухом, но жили в морской воде, они не могли откладывать яйца. Поэтому у ихтиозавров появился способ размножения, при котором зародыш развивался внутри тела матери и, достигая зрелости, рождался живым. Они стали живородящими. Этот факт устанавливается находками превосходно сохранившихся остатков самок ихтиозавров с полностью сформировавшимися детенышами внутри их тела, число детенышей достигает семи.
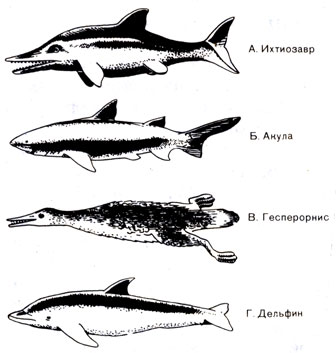
Рис. 50. Четыре группы животных, которые приобрели обтекаемую форму тела в результате приспособления к жизни в воде: А. рептилия, Б. рыба, В. птица, Г. млекопитающее. Первоначально они имели различный облик, но в ходе эволюции приобрели внешнее сходство
Вторая группа включает плезиозавров, которые в отличие от рыбообразных ихтиозавров сохранили исходную форму тела рептилий, достигая 7,5-12 метров в длину. Если бы не хвост, плезиозавр походил бы на гигантского лебедя. Конечно, предком плезиозавра была совсем не та наземная рептилия, которая дала начало ихтиозаврам. Ноги у плезиозавров превратились в длинные плавники, а голова, посаженная на длинной шее, была снабжена острыми зубами, которые смыкались и надежно удерживали самую скользкую рыбу. Такие зубы исключали пережевывание; плезиозавр заглатывал добычу целиком и затем измельчал ее в желудке с помощью камешков. О рационе плезиозавров можно судить на примере содержимого желудка одного из них, который, по-видимому, погиб прежде, чем камни в его желудке успели измельчить проглоченную им пищу в должной степени. Было установлено, что содержавшиеся в желудке кости и обломки раковин принадлежат рыбам, летающим рептилиям и головоногим, которые проглатывались целиком, вместе с раковиной.
Третья группа морских рептилий получила название мозазавров, потому что они впервые были обнаружены вблизи реки Мозель, в северо-восточной Франции. Их можно было бы назвать "запоздалыми", потому что они появились тольдо в позднеме-ловое время, когда ихтиозавры заселяли моря уже в продолжение почти 150 миллионов лет. Предками мозазавров были скорее ящерицы, чем динозавры. Длина их достигала 9 метров, они имели чешуйчатую кожу, а челюсти у них были устроены таким образом, что они могли широко открывать рот, как змеи.
Обтекаемое тело как приспособление к условиям жизни в водной среде встречается не только у ихтиозавров и мозазавров. То же самое можно видеть у целого ряда животных, живших и раньше, и позже мезозоя, и в мезозое (рис. 50).
Рептилии в воздухе
История расцвета рептилий в мезозое не заканчивается тем, что изложено выше. Рептилии не только распространились на суше и заполнили моря, они также поднялись в воздух, следуя сразу по двум линиям эволюции. Они научились летать и как рептилии, и кроме того, двигаясь совсем по другому пути развития, они научились летать и как птицы. Насколько можно судить по ископаемым остаткам, настоящие летающие рептилии были не такими многочисленными, как морские. Однако они были первыми животными, поднявшимися в воздух после насекомых, сделавших это еще в девонское время. Естественно, что воздушная среда труднее для завоевания и более опасна, чем море. Для того чтобы двигаться в воздухе или хотя бы пассивно парить, требуются более специализированные приспособления, большая энергия и мастерство (под которым мы подразумеваем подвижность и быструю реакцию), чем для передвижения в воде. В основном именно поэтому человек построил корабли задолго до аэропланов. Промежуток между этими изобретениями человека составил около нескольких тысяч лет. А между возникновением рептилий в позднекаменноугольное время и их проникновением в воздух (юрское время) прошло около 80 миллионов лет.
Мы многое знаем о строении и облике летающих рептилий благодаря тому, что в южной части Германии [ФРГ, Бавария. - Ред.] широко распространены осадочные породы необычного типа. Эти породы представляют собой пласты известняка позднеюрского возраста, настолько тонкозернистого, что он применялся для гравировки иллюстраций для книг (до того, как для этой цели стали использовать стальные и медные пластинки) и за это получил название литографского камня. Необыкновенно тонкозернистый состав этих известняков заставляет предполагать, что они отлагались в мелководных лагунах, защищенных от волнения открытого моря песчаными барами или коралловыми рифами. Рыхлые отложения на дне лагун сохраняли отпечатки даже мельчайших деталей растений или тел животных, которые опускались на дно и заносились илом. В результате литографский камень славится содержащимися в нем ископаемыми остатками растений, беспозвоночных, рыб и рептилий.
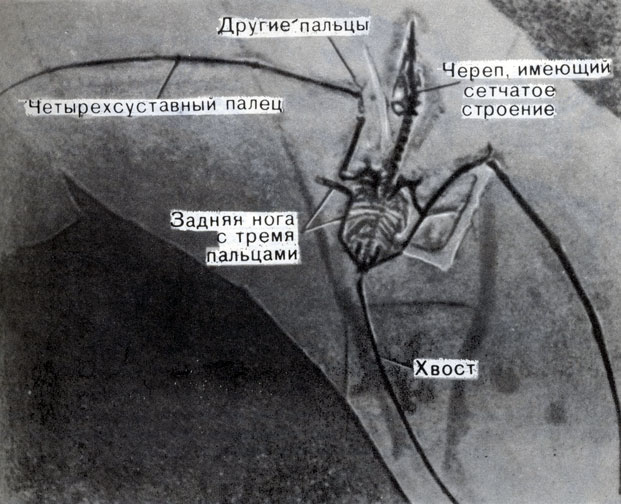
Фото 46 Скелет Rhamphorhynchus, примитивной летающей рептилии, найденный в литографских известняках в Германии
В этих отложениях были найдены многие крылатые рептилии, кроме того, подобные остатки были найдены и в других слоях мезозойского возраста в различных местах. Рассматривая остатки одной из юрских примитивных рептилий, сохранившейся до мельчайших подробностей (фото 46), мы видим, что ее тело приспособилось к полету следующим образом: 1) уменьшился вес; 2) появились "устройства" для управления полетом; 3) был создан механизм для полета. Вот некоторые из этих приспособлений:
1. Малые размеры тела; хотя некоторые летающие рептилии были с индейку, другие были не больше канарейки. Скелет был облегчен благодаря развитию тонких полых костей крыльев, а у некоторых видов и череп имел почти сетчатое строение и состоял из тонких костей.
2. Необычайно, хорошим развитием отличались глаза и часть мозга, управляющая зрением.
3. Наиболее замечательной особенностью были крылья. Глядя на рисунки 51 и 52, мы можем легко себе представить, что четвертый палец на передней конечности, словом "мизинец", - был необычайно удлинен по отношению к остальным. От кончика этого пальца к задней ноге и далее к хвосту протягивалась тонкая перепонка из кожи, образовывавшая крыло.

Рис. 51. Птеранодон (Pteranodon), летающая рептилия с выростом на черепе; он пролетал огромные расстояния над обширными меловыми морями на территории нынешних штатов Канзас и Небраска
Все эти три группы приспособлений, вместе взятые, создавали устройство, которое хотя и неуклюже, но могло летать. Образование крыла, сопровождавшееся усовершенствованием глаза и уменьшением общего веса, сделало возможным полет привело к удивительным пропорциям тела. Например, одна из летающих рептилий с размахом крыльев в 90 сантиметров, согласно расчетам, при жизни весила меньше 450 грамм. Кожа таких рептилий была голая, а челюсти снабжены многочисленными острыми зубами, обычными для рептилий. Вероятно, эти животные, как современные канюки, больше парили, чем летали. Происходя от наземных хищников, они, очевидно, остались плотоядными и, медленно скользя над водой, высматривали морских животных или крупных насекомых. Строение их скеле показывает, что они не могли ходить. Очевидно, они приземлялись не на поверхность земли, а на ветви деревьев или выступы скал на которых висели, как современные летучие мыши.
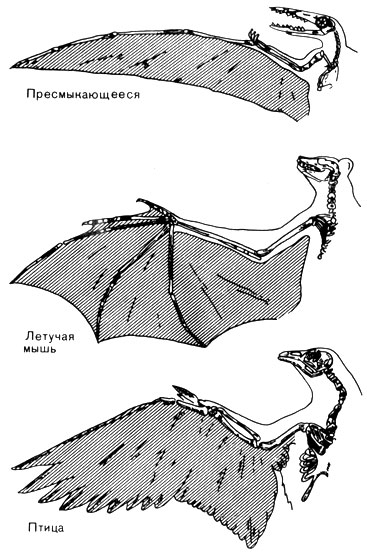
Рис. 52. Схема сравнения крыла летающей рептилии, летучей мыши и птицы. Все эти крылья появились в различное время. У рептилии все крыло поддерживается только одним пальцем. У летучей мыши внешняя часть крыла усилена четырьмя пальцами. У птицы большая часть крыла поддерживается костями плеча и предплечья, а несущая поверхность образована легкими жесткими перьями. Из всех трех видов такое крыло наиболее соответствует своему назначению
Позже развитие летающих рептилий, остатки которых найдены в отложениях мелководных морей мелового времени, пошло по пути замены зубов длинным клювом, который, конечно, лучше соответствовал их образу жизни. У одного из родов в задней части черепа появился особый выступ, или гребень (рис. 51), который, возможно, уравновешивал длинный клюв и облегчал рептилии маневрирование при ветре. Но главное изменение касалось увеличения площади крыльев, очевидно, чтобы лучше поддерживать тело в воздухе. Одна из летающих рептилий для поддержки тела, которое предположительно весило менее 12 килограммов, имела крылья с размахом 7,5 метра. Такой размах крыльев позволяет считать этих рептилий самыми крупными летающими животными за всю историю существования жизни на Земле. Хотя до самого конца своего расцвета летающие рептилии отличались хрупким сложением, все же они просуществовали более 100 миллионов лет.
Но несмотря на то, что крыло рептилий выполняло свои функции и просуществовало долго, оно было менее удачным приспособлением для полета, чем появившееся независимо от него крыло птицы и позднее млекопитающих - летучих мышей. На рисунке 52 показаны все три крыла, и, как можно видеть, крыло птицы из них самое совершенное.
Птицы
В юрское время у рептилий, живших по берегам теплых морей, существовали различные виды полета. Мы уже видели, что несколько видов наземных рептилий поднялись в воздух, используя для этого только что описанные кожистые крылья. Но один из видов пошел еще дальше. В одном из карьеров при разработке литографского камня в середине XIX в. был найден ископаемый скелет рептилии, размером не больше вороны, имевшей большие глаза, зубы, как у рептилий, и пальцы с когтями на передних конечностях. Поразительно, что при этом были обнаружены очень четкие отпечатки перьев, крепившихся к предплечью и к позвонкам длинного хвоста. Это, несомненно, была птица. Она получила родовое имя археоптерикс (Archaeopteiyx) ("древнее крыло") и видовое название Uthographica по названию горной породы (фото 47). В том же самом слое были обнаружены еще два ископаемых скелета и отдельный отпечаток пера.

Фото 47. Археоптерикс (Archaeopteryx), древнейшая известная нам птица, сидит на ветке хвойного дерева, собираясь съесть пойманную ящерицу. На переднем плане справа - цикадовые растения; сзади - хвойные деревья и еще одна такая же птица. Реконструкция
Конечно, эти находки представляли необычайный интерес и потому были тщательно изучены. Результаты исследований могут быть, очевидно, сведены к следующему: археоптерикс по своим основным чертам - летающая рептилия, но так как, по определению, птицы имеют перья, а рептилии - нет, то ее можно отнести к птицам. Особенности строения археоптерикса позволяют уверенно сказать, что эта древнейшая из известных нам птиц произошла от жившей на земле двуногой рептилии. Наличие перьев убедительно свидетельствует о том, что она имела теплую кровь, потому что одной из основных функций перьев является термоизоляция. У многих птиц кровь даже теплее, чем у человека. Их перьевой покров и высокая двигательная активность позволяют им поддерживать нормальную температуру тела около 39,5° С.
Перья состоят из того же прочного рогового вещества, что и чешуя. Некоторые ученые предполагают, что небольшая рептилия, которая была предком этих примитивных птиц, имела чешую и что сначала чешуйки стали волнистыми по краям, возможно, потому, что такая форма предохраняла кожу от перегрева солнечными лучами. Волнистые края оказались полезны и в другом отношении, поскольку они уменьшали потерю телом тепла, и постепенно эти чешуйки превратились в перья. Жесткость и небольшой вес вещества перьев сделал их идеальным приспособлением для полета.
Хотя первая птица и имела перья, она, как и ее родственники - летающие рептилии с кожистыми крыльями, летала плохо. Ее строение указывает на то,что птица, вероятно, была хорошо приспособлена к планирующему полету. Возможно, она жила на суше и, будучи хищником, питалась или мелкими животными, или падалью. То, что ее остатки были найдены в морских известняках, свидетельствует лишь о том, что отдельные экземпляры были унесены в море ветром или течением и погребены в мягком донном иле. Хрупкие тела птиц, погибших на суше, просто не сохранились.
К меловому периоду неуклюжесть полета у птиц исчезла и многие из них вместо зубов обзавелись клювом. Некоторые птицы приспособились к жизни на воде. Примером является весьма походившая на гагару плавающая и ныряющая птица гесперорнис (Hesperornis) (рис. 50), длина которой была около двух метров и которая все еще обладала зубами и крыльями, хотя и не такими сильными и меньшего размера, чем у летающих птиц. Существование птиц, которые почти покинули воздушную среду, перейдя к плаванию, свидетельствует о том, что на ранних этапах развития птицы охотились на рыбу так же, как это постоянно делали рептилии с начала мезозойской эры.
Конец огромных рептилий
Окончание мелового периода, означавшее конец всей мезозойской эры, можно назвать "кризисом" в истории биосферы, потому что в это время произошло вымирание многих групп животных. Наиболее заметный урон понесли рептилии. Вымерли все динозавры, все летающие рептилии и все морские рептилии, за исключением морских черепах; выжили и продолжили линию рептилий только ящерицы, змеи и черепахи. Из беспозвоночных исчезли большинство головоногих, в том числе все белемниты, а также некоторые линии морских двустворчатых моллюсков и улиток.
Вымирание носило выборочный характер, потому что млекопитающие и наземные растения были им мало затронуты или совсем не затронуты, а рыбы и многие беспозвоночные полностью уцелели. Поэтому попытки приписать это вымирание какой-либо одной причине оказались неудачными. Пока время основных событий в истории Земли не было установлено с помощью радиометрического датирования, конец мезозоя обычно именовался временем "великого вымирания". Однако сейчас мы сознаем, что это выражение не соответствует истине. По крайней мере два обстоятельства свидетельствуют о том, что вымирание не носило характера катастрофы, уничтожившей все живое.
Во-первых, оно было избирательным, затрагивая одни виды и щадя другие. Кроме того, оно не было ограничено каким-либо одним видом природной среды, охватывая сушу, море и воздух. Во-вторых, хотя наиболее заметным было вымирание видов в конце мелового периода, оно заняло в целом значительный промежуток времени. В частности, различные группы рептилий вымирали в отдельные моменты времени в течение всего мезозоя. Поэтому, какова бы ни была причина этого явления, очевидно, она не вызвала "внезапного" истребления видов, по крайней мере в том смысле этого слова, в каком мы применяем его к событиям истории человеческого общества. Даже наиболее ярко выраженное вымирание, происходившее в конце мелового периода, вероятно, тянулось несколько миллионов лет.
Рассматривая геологические данные, говорящие о том, что происходило в конце мелового периода, мы видим, что континенты в целом стали выше. В то же время и, возможно, главным образом в результате этого поднятия сократились площади обширных мелководных морей на континентах и исчезли болотистые низменности, располагавшиеся по берегам этих морей. Кроме того, понизилась температура, частично благодаря поднятию и сокращению площади морей.
Мы должны признать, что действительная причина вымирания до сих пор не установлена. Ранее выдвигавшиеся объяснения - болезнь, недостаток пищи и самое туманное - "потеря жизненной силы" - совершенно не в состоянии объяснить, почему произошло выборочное исчезновение некоторых из обитателей суши, моря и воздуха, а не полное вымирание обитателей какой-либо одной среды. Представляется, что млекопитающие вышли из этой катастрофы целыми и невредимыми.
Недавно было выдвинуто предположение, что конец мезозоя ознаменовался серией обращений магнитного поля Земли (описанных в шестой главе) и что эти обращения могли каким-либо образом повлиять на биосферу, например путем изменения интенсивности радиации, поступающей на земную поверхность. Против этого выдвигались возражения, но, возможно, сейчас еще слишком рано оценивать все "за" и все "против". Достаточно сказать, что вымирание, которым ознаменовался конец "века динозавров", до сих пор представляет собой одну из величайших загадок, связанных с историей жизни на Земле.
Литература
Augusta Josef, Burian Zdenek. 1961, Prehistoric reptiles and birds: Paul Hamlyn, London.
Colbert E. H., 1951, The dinosaur book: NcGraw-Hill Book Co., Inc., New York.
Colbert. E. H., 1961, Dinosaurs. Their discovery and their world: E. P. Dutton &Co.. Inc., New York.
Fentоn C.L., Fentоn M. A., 1958, The fossil book: Doubleday & Co.. New York, p. 329-374.
Кurten Bjorn, 1968, The age of the dinosaurs: Weidenfeld and Nicolson, London. (Paperback.)
Swintоn W. E., 1958, Fossil birds: British Museum (Natural History), London.
Swintоn W. E.. 1970, The dinosaurs: Wiley-Interscience, New York.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'