
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Плечевой пояс и передняя конечность
Скапулокоракоид. Обычно коракоид составляет примерно третью или четвертую часть длины всей структуры. Это выпуклое кнаружи, нередко утолщенное в центре и утончающееся по периферии образование, у ряда форм, возможно, переходившее по краям в хрящ. Скапулокоракоидный шов более или менее прямой. Коракоидное отверстие косо пронизывает кость. Ниже и сзади коракоидного отверстия по наружной поверхности коракоида расположена выпуклость с заостренной вершиной, несущей следы прикрепления сухожилий двуглавой мышцы. Гленоидная впадина довольно глубокая и широкая, слегка обращенная наружу, с выпуклыми гребневидными краями. Супрагленоидное утолщение в форме гребня или выступа, разделенного вдоль по вершине скапулокоракоидным швом. Внутренняя поверхность скапулокоракоида широко вогнутая.
Лопатка с расширенной вентральной частью и суженной дорзальной ветвью, края которой часто субпараллельны друг другу. От гленоидной впадины, захватывая нижнюю часть задней половины дорзальной ветви, простирается утолщение, рельефно выраженное на ее внутренней поверхности. Акромиальный отросток хорошо развит. Его передний край нередко значительно утончен, возможно, переходил в хрящ.
У разных групп хищных динозавров при общем типе строения скапулокоракоида наблюдались и немалые отличия, преимущественно в форме и пропорциях отдельных элементов структуры. Так, у теризинозаврид коракоид пропорционально очень крупный, широкий, тогда как лопаточная ветвь сильно укорочена и по длине даже меньше коракоида, что необычно для теропод. К примеру, скапулокоракоид у дейнохейрид, вполне сопоставимый по общим размерам с теризинозавровым, вполне соответствует общему типу и состоит из относительно узкой и удлиненной лопатки и относительно небольшой коракоидной части. Кроме того, вблизи переднего края лопаточного расширения около скапулокоракоидного шва у теризинозаврид располагалось округлое отверстие, не отмеченное у других теропод, через которое, по-видимому, проходили кровеносные сосуды и нервы. Тираннозавриды характеризуются сильно удлиненной лопаточной ветвью, слегка расширявшейся дорзально. У мелких теропод - целурид, дромеозаврид, овирапторид - скапулокоракоиды обычного типа с удлиненной лопаточной ветвью и относительно небольшим коракоидом. Однако у овирапторид отмечено необычное строение переднего края акромиального отростка, который значительно утолщен и уплощен спереди с образованием хорошо выраженной узкой продольной площадки (рис. 16). К ней, как оказалось, плотно прилегает ветвь сросшихся ключиц, соединяющих таким образом скапулокоракоиды. У других теропод подобной площадки не образуется, однако в плечевом поясе у археоптерикса (Ostrom, 1976 а, с) отмечается совершенно сходные по положению и форме сочленовные площадки, что дает веское основание считать их местом присоединения ключиц.
Ключицы. Ключицы в ископаемом состоянии у динозавров, в том числе и у хищных, находили редко и плохо сохранившимися. У двух представителей теропод - овираптора и сегнозавра - ключицы первоначально описывались, как межключицы (Os-born, 1924; Camp, 1936). Лишь последние находки овирапторид дали полное представление о строении этого элемента у теропод (рис. 17).
Ключицы у овирапторид по общим очертаниям вполне сходны с вилочкой (furcula) у птиц и представляют собой V-образную сросшуюся структуру с широко расходившимися дорзально вогнутыми ветвями, концы которых клиновидно уплощаются. Область расхождения ветвей с клювовидным каудально направленным укороченным отростком расширена. Уплощенными концами ветвей ключицы (рис. 18) ложились на акромиальные отростки лопаток, соединяя скапулокоракоиды. В этом положении клювовидный задний отросток располагался между близко расположенными коракоидами и, вероятно, имел с ними связь.
В соединении ключиц со скапулокоракоидами привлекает внимание специальное приспособление, отмечавшееся выше - передние края акромиальных отростков уплощены с образованием площадки треугольного очертания, принимающей наружный конец ключицы. Предполагается, что эта площадка служит признаком сохранения ключиц в плечевом поясе динозавров. Однако у других теропод краевая часть акромиона не имеет следов сочленовной площадки, зачастую она явственно уплощена вплоть до состояния тонкой лопасти, что, возможно, говорит об отсутствии ключиц или же допускает лишь лигаментарное соединение с ними. Следует отметить, что сходная по строению вилочка у птиц своими концами соединяется с коракоидами, а не с акромиальными отростками лопаток, имеющими соответствующие суставные площадки.
У археоптериксов найдены ключицы (Ostrom, 1976 а, с), чрезвычайно сходные с овирапторидными, однако способ соединения с плечевым поясом остался неизвестным до тех пор, пока не выяснилось специфическое соединение ключиц с акромиальными отростками у овирапторид.
Плечевая кость. Различия в строении плечевой кости составляют одну из основ при выделении хватательной (относительно сильно развитой) и нехватательной (относительно слабо развитой) передней конечности у теропод. В первом случае крайне характерно присутствие высокого и протяженного дельтовидного гребня, имевшегося у большинства теропод. Вместе с тем, в случае нехватательной передней конечности дельтовидный гребень был слабо развит, что наблюдается у дейнохейрид и орнитомимид.
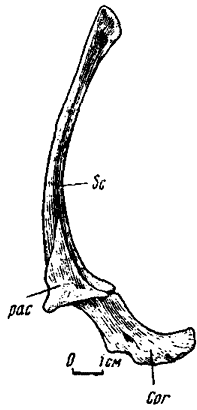
Рис. 16. Правый скапулокоракоид Oviraptor philoceratops (№ 100/42, спереди)
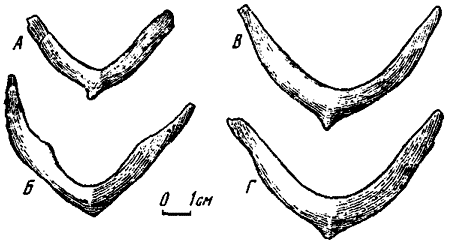
Рис. 17. Ключицы спереди: А - Oviraptor philoceratops (№ 100/36); Б - Oviraptor philoceratops (№ 100/42); В - Ingenia yanshini (№ 100/30); Г - Ingenia yanshini (№ 100/33)
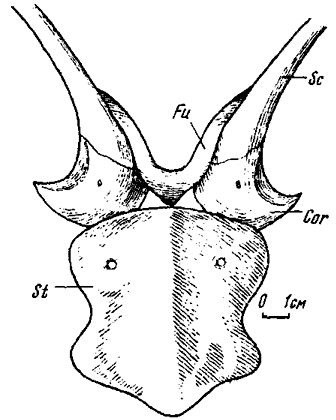
Рис . 18. Соединение ключиц, скапулокоракоидов и грудины у овирапторид (схематическая реконструкция)
Дельтовидный гребень у тираннозаврид был сравнительно высоким, однако существенно укороченным, что сразу же отличает его от структуры развитого типа. При достаточном развитии дельтовидный гребень занимает около трети всей длины плечевой кости, а у теризинозаврид даже немногим более половины. В случае ослабления гребень обычно не выходит за пределы одной трети, у тираннозаврид - даже четверти длины всей кости. Характерной особенностью служит также степень сигмоидальной изогнутости плечевой кости, что наиболее резко выражено у теризинозаврид и мелких теропод и практически утрачено в нехватательной передней конечности. Достаточная оформленность суставной головки, в особенности у теропод с развитой передней конечностью, свидетельствует о хорошей подвижности в плечевом суставе. Дистальный блок плечевой кости был значительно расширенным и передне-заднеуплощенным. В этом случае мыщелки дистального конца имели достаточное развитие и характеризовались смещенностью вперед, вследствие чего эпиподии в нормальном положении должны были быть несколько согнутыми в локтевом суставе. В конечности же нехватательного типа дистальный блок расширен и уплощен, слабо развитые мыщелки обращены более дистально и, следовательно, конечность характеризовалась большей выпрямленностью.
Предплечье. Локтевая кость более массивная, чем лучевая, чаще выпрямленная, у орнитомимид и дейнохейрид слегка выгнутая вперед. Ствол кости трехстороннеуплощенный в связи с расположением лучевой кости и соответствующих групп мышц. Вырезка локтевого сустава довольно глубокая: ее суставная поверхность треугольно-округлая, подразделяется на медиальную и латеральную части, из них латеральная входит между мыщелками плечевой кости, имеет скорее не суставную, а опорную функцию и представляет собой новообразование архозавров, отмечаемое у текодонтов, динозавров, крокодилов (Romer, 1956). Олекранон развитый. Дистальный конец кости нередко уплощен, его присуставная зона со следами прикрепления мышц - разгибателей и сгибателей кости.
Лучевая кость сигмоидально изогнутая (теризинозавриды), выпрямленная (дромеозавриды, овирапториды), слабо вогнутая вперед (дейнохейриды, орнитомимиды). Проксимальный и дистальный концы уплощены и расширены в пересекающихся под разными углами плоскостях. Присуставные края дистальной части со следами прикрепления луче-локтевых или луче-карпальных связок.
Кисть. Кисть у хищных динозавров имеет тенденцию к сокращению числа пальцев. В общем для наиболее ранних теропод характерна пятипалая кисть, для поздних же в связи с редукцией боковых пальцев в большинстве случаев устанавливается трехпалость и редко двупалость. Редукцию числа пальцев кисти можно рассматривать, как общую закономерность в развитии группы. В последние годы выяснилось, что эта общая тенденция осуществляется на основе двух разных типов строения карпальных элементов, приводивших в итоге к двум разным модификациям кисти, функциональные возможности которых были во многом неодинаковы. Как оказалось, структура запястья определяется шкивным и нешкивным характером его составляющих.
Шкивная структура запястья (рис. 19). В основе ее выделения лежит специфическое строение карпальных элементов, впервые подробно (хотя и не без отдельных ошибок) показанное на примере кисли американского дромеозаврида - дейнонихуса (Ostrom, 1969b). Она описана также у овирапторид и теризинозаврид (Барсболд, 1976б). Число карпальных элементов в этом типе кисти испытывает сокращение и в наиболее выраженной форме достигает двух - дистального и проксимального. При этом точное отождествление карпальных элементов большей частью едва ли - возможно. Тем более не поддается учету возможность слияния отдельных карпальных элементов при их численном сокращении.
Дистальный карпальный элемент имеет форму половины шкива (рис. 20), дуговая, обращенная к предплечью часть которого несет продольный желоб, ограниченный валикообразными краями. Пястная сторона полушкива с двумя суставными поверхностями, соответствовавшими проксимальным окончаниям метакарпальных костей, с которыми они прочно соединены. Проксимальный элемент заметно уменьшен, по форме округленно-конический, основанием неподвижно прикреплялся к дистальному суставному концу лучевой кости. Судя по прикреплению, проксимальный элемент может быть отождествлен с os radiale. Этот элемент входил в желоб шкива, тем самым формируя подвижное соединение. Шкив с прочно прикрепленной к нему кистью мог двигаться относительно неподвижного проксимального карпального элемента вдоль дуги окружности, размеры которой ограничивались длиной продольного желоба. Хотя желоб как будто бы ограничивал движение шкива с кистью преимущественно плоскость шкива (Барсболд, 1976б), ширина желоба и округленно-коническая форма проксимального карпального элемента свидетельствуют в пользу достаточной степени свободы движения и в иных плоскостях. По всей вероятности, шкивное запястное соединение вполне позволяло и вращательные движения кисти.
Шкивное строение запястья, но с большим числом дистальных карпальных элементов, найдено у теризинозаврид. В этом случае шкив состоит из двух дистальных элементов, больший (медиальный) из которых, возможно, гомологичен единому шкиву, рассмотренному выше. Меньший (латеральный) элемент, служащий продолжением большего, плотно входит в широкую вырезку последнего. Дуговой желоб отличается значительной шириной и на проксимальной поверхности меньшего элемента почти сходит на нет. Все это должно было также обеспечивать значительную степень свободы движения в запястье. Ширина желоба в определенной мере свидетельствует о достаточных размерах дистального элемента, число которого могло быть и более одного.
Нешкивная структура запястья (рис. 19Б). Такой тип структуры запястья определяется наличием большего числа карпальных элементов, не формирующих шкив. Он известен у тираннозаврид, орнитомимид, а также у более древних групп. Число карпальных элементов в этом типе не достигает такого сильного сокращения, как в шкивной структуре и в большинстве случаев равняется четырем - шести. В нешкивном запястье точное отождествление элементов также затруднено. Элементы запястья обычно уплощены, более или менее однообразны по форме и составляют дистальный и проксимальный ряды. Нередко дистальный элемент, начинающий ряд, соответствующий по положению первой метакарпальной кости, может выделяться более крупной величиной. В проксимальном ряду запястья суставным поверхностям лучевой и локтевой костей отвечают уплощенные элементы, которые нередко отождествляют с radiale и ulnare. Иногда между ними выделяют os intermedium. Независимо от того, насколько обоснована подобная гомологизация, нешкивная структура запястья вполне сохраняет свое единообразие у различных групп. Двурядная карпальная структура не меняется и при возможно минимальном числе элементов, равном четырем, отмеченному у монгольских тираннозаврид (тарбозавров). У орнитомимид в запястье предположительно было пять элементов (Osborn, 1917).
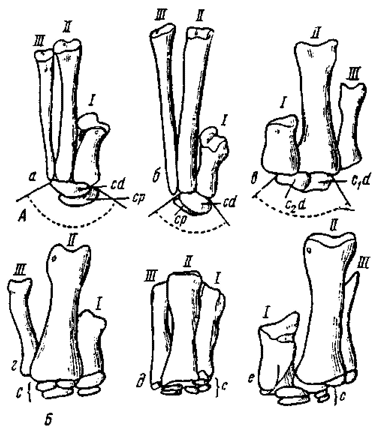
Рис. 19. Строение пястья и запястья (вид сверху, без масштаба, реконструкция): А - в шкивном типе: а - у велоцираптора; б - овираптора; в - теризинозавра; Б - в нешкивном типе: г - у аллозавра (Marsh, 1877); д - орнитомима (Romer, 1956); е - тарбозавра
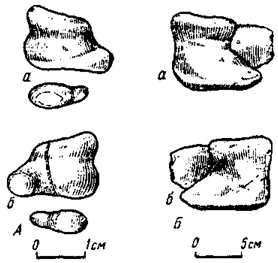
Рис. 20. Строение запястного шкива (схематизировано): А - у неопределенного монгольского дромеозаврида: а - с проксимальной стороны; б - с дистальной стороны; Б - у теризинозавра: а - с проксимальной стороны; б - с дистальной стороны
Не вызывает сомнений, что нешкивная структура карпальных элементов была гораздо менее компактной и обеспечивала меньшую подвижность, чем шкивная. Предполагается, что такая структура была более ограничена во вращательных движениях кисти, требующих большого совершенства суставного соединения в запястье.
Пясть. Позднемезозойские хищные динозавры характеризуются обычно присутствием трех элементов в пясти (рис. 19). Из них первый наиболее укорочен, чаще всего составляет по длине примерно половину второго, третий же значительно редуцирован. Однако у дейнохейрид и орнитомимид все составляющие пясти примерно равны по длине. Кроме того, первый метакарпальный элемент у большинства меловых теропод сильно уплощен медио-дорзально, тогда как у орнитомимид и, в особенности, у дейнохейрид подобная скошенная уплощенность выражена существенно слабее. Проксимальная суставная поверхность первого элемента характерной треугольной или трехлучевой формы, при этом у упомянутых выше двух групп центральная часть поверхности с широкой вогнутостью, отсутствующей у других теропод. Асимметричный суставной дистальный блок нередко обращен несколько медиально, что слегка отклоняет первый палец от второго.
Второй метакарпальный элемент обычно наиболее удлинен и довольно утолщен, дистальный суставной блок умеренно асимметричен. У овирапторид ингений второй элемент заметно редуцирован и утончен.
Третий метакарпальный элемент обычно наиболее редуцирован, чаще более укорочен, чем второй, однако у дейнохейрид он намного длиннее. Наибольшая степень редукции отмечена у тираннозаврид, у которых третий элемент превращен в "грифельную" косточку, лишенную пальца. Значительная редукция известна также у дромеозаврид и овирапторид, однако соответствующий палец сохраняется полностью. Проксимальная суставная поверхность кости при ее меньшей редукции треугольная, при большей - латерально сильно сжатая.
Как явствует из строения запястья, в особенности, шкивного типа, подвергшийся значительно редукции третий метакарпальный элемент практически был выключен из суставного соединения с ним. Во всяком случае, запястье не имеет такой связи с третьим элементом, как с первыми двумя, что особенно хорошо видно на примере у дромеозаврид и овирапторид.
Пальцы кисти. Общая фаланговая формула кисти у позднемезозойских теропод 2-3-4, однако разнообразие в строении пальцев, как оказалось, достаточно велико (рис. 21). Вместе с тем ряд черт прослеживается через все разнообразие вполне устойчиво.
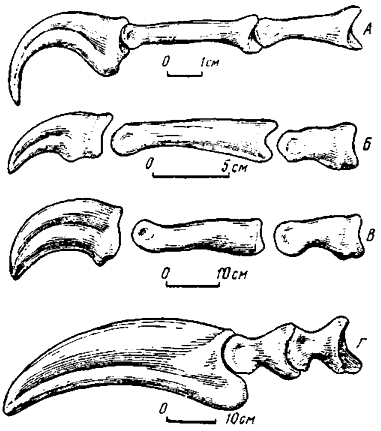
Рис. 21. Второй палец кисти изнутри: А - Velociraptor mongoliensis (Ostrom, 1969b); Б - Gallimimus bullatus (Osmolska, Roniewicz, et Barshold, 1972); В - Deinocheirus mirificus (Osmolska, Roniewicz, 1970); Г - Therizinosaurus cheloniformes Maleev (Барсболд, 1976в)
Первый палец или уступает по длине остальным, особенно существенно второму (дромеозавриды, заурорнитоидиды, тираннозавриды, некоторые овирапториды), или близко приближается к их длине (орнитомимиды, дейнохейриды), или же превосходит другие (ингении). Характерно, что базальная фаланга первого пальца наиболее удлинена по сравнению с остальными (исключая когтевые). Из остальных пальцев кисти второй или наиболее удлинен, или приблизительно равен третьему (в этом случае они примерно одинаковы и по степени редукции), или же длиннее третьего, но короче первого. Отмеченные признаки II и I пальцев для приведенных групп располагаются в одинаковой последовательности. Третий палец наиболее редуцирован, нередко значительно утончен, однако все фаланги сохраняются. Заметная степень редукции третьего пальца у поздних групп теропод широко распространена, однако лишь у двух групп заходит достаточно далеко. Как отмечалось, тираннозавриды практически лишены третьего пальца (присутствует лишь проксимальная часть базальной фаланги), а в кисти овирапторид ингений и второй, и третий пальцы испытывали нарастающую редукцию, однако они сохранили все фаланги. Весьма распространено удлинение предкогтевых фаланг, имеющее место и у некоторых хищных птиц. Отступление от этого правила наблюдается у ингений, фаланги которых в дистальном направлении уменьшаются вполне последовательно и, возможно, у теризинозаврид, судя по единственному второму пальцу с почти равновеликими предкогтевой и базальной фалангами. За исключением указанных групп, базальная фаланга второго пальца, а также базальная и вторая фаланги третьего пальца наиболее коротки. Фаланги у многих групп характеризуются хорошо развитыми расширенными гинглимоидными суставами. Вместе с тем у дейнохейрид, орнитомимид и тираннозаврид суставные блоки разработаны гораздо слабее.
Разнообразие пальцев кисти у хищных динозавров в значительной мере обязано когтевым фалангам, форма и размеры которых изменялись в широких пределах. Ныне не будет преувеличением сказать, что почти каждая более или менее обособленная группа хищных динозавров характеризуется специфическим строением когтевых фаланг (Taquet, 1976). Только длина когтевых фаланг варьировала от 1,0-1,5 см у некоторых овирапторид до 50-60 см и, возможно, более у теризинозаврид. Вместе с тем, отчетливо проявляются общие черты когтевых фаланг хищных динозавров. Как правило, они в большей или меньшей степени сжаты с боков, нередко дистально заострены, по-разному, но в большинстве случаев достаточно сильно изогнуты, вентральные и дорзальные бугры для прикрепления связок сгибателей и разгибателей часто хорошо развиты.
На фоне рассмотренных общих черт ярко выделяются отличительные особенности, свойственные многим группам. Дромеозавриды имели наиболее сжатые с боков и крюковидно изогнутые и заостренные когтевые фаланги с крупными связочными буграми, тогда как у орнитомимид они были более выпрямленными, со сравнительно слабым развитием бугров. Дейнохейриды отличались крупными, массивными, утолщенными когтевыми фалангами. Теризинозавриды, пожалуй, не имели себе равных по величине гипертрофированных когтевых фаланг, к тому же сильно расширенных и слабо изогнутых. У собственно овирапторов когтевые фаланги вполне соответствовали общетероподовому стандарту, однако у ингений вследствие необычно сильной редукции второго и третьего пальцев кисти когтевые фаланги крайне уменьшены и кроме того, отличаются значительной выпрямленностью и слабым развитием связочных бугров (рис.22).
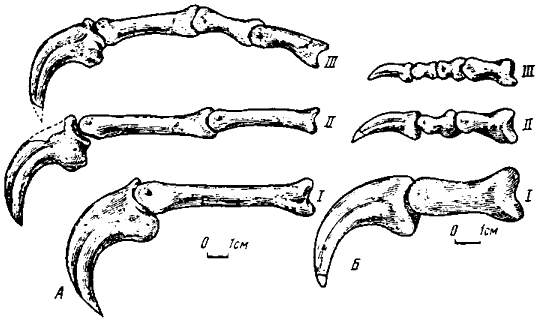
Pис. 22. Пальцы правой кисти изнутри: А - Oviraptor philoceratops (№ 100/42) : Б - Ingenia yanshini (№ 100/30)
В заключение можно сказать, что все разнообразие в строении кисти у хищных динозавров удается сводить к двум модификациям, в основе которых лежит шкивная или нешкивная структура запястья. Ранее считалось, что снабженная изогнутыми когтями кисть у хищных динозавров выполняла функции захватывания добычи. Некоторое отклонение первого пальца иногда неправильно рассматривалось, как противопоставление остальным, вероятно, по примеру высших приматов, что способствовало укоренению понятия о "хватательной" кисти у теропод. И сейчас, возможно, не стоит отказываться от этой концепции, хотя кисть у теропод первоначально скорее была приспособлена к раздиранию тела жертвы, чем к выполнению хватательного действия. Если считать, что заостренные, сильно изогнутые когти на удлиненных передкогтевых фалангах способствовали удерживанию, т.е., захватыванию жертвы передними конечностями, то в этом смысле кисть у дромеозаврид и у заурорнитоидид безусловно заслуживает такого определения. Удалось показать, что наиболее ярко выраженная хватательная кисть имеет в запястье шкивную структуру, допускавшую более совершенную подвижность. Взаимосвязь между повышенной степенью "хватательности" и более совершенной структурой запястья кажется вполне закономерной. Однако "хватательность" не всегда должна пониматься буквально. Есть существенные отклонения, и лучший пример - кисть у теризинозаврид. Уже наличие громадных когтей, превосходивших общую длину всех остальных структур в кисти, полностью исключает какую-либо хватательную функцию. Тем не менее, шкивная структура свидетельствует в пользу "хватательной" основы, на которой такая кисть развивалась. Раздельный шкив в запястье у теризинозаврид, состоящий из двух частей, вполне ясно показывает его относительную примитивность по сравнению с единым шкивом у дромеозаврид и овирапторид. Таким образом, на основе шкивной структуры запястья первоначально развивалась кисть хватательного типа, получившая ряд отклонений у различных специализированных групп, в том числе и у теризинозаврид. Необходимо отметить, что у всех групп с явно выраженным хватательным типом кисти мускулатура передних конечностей усилена, что ясно отражено в рельефности костей плеча (крупные дельтопекторальные гребни) и предплечья, а также в разработанности их суставных блоков.
Вторая модификация кисти строилась на нешкивной основе. Она характерна для орнитомимид, тираннозаврид и, вероятно, дейнохейрид. Кисть у первой и последней групп по ряду признаков довольно сходна, хотя размеры их совершенно несопоставимы. Это сходство, отмеченное выше, а также черты явного ослабления передней конечности, равно как и сравнительно малая степень сгибания пальцев (Ostrom, 1969b; Osmolska, Roniewicz, Barsbold, 1972; Osmolska, Roniewicz. 1970), отнюдь недостаточная для какого-либо захватывания, свидетельствует в пользу нехватательного типа кисти у обеих групп. Ослабление передних конечностей у орнитомимид и дейнохейрид ясно выражено в утончении костей плеча и предплечья, развитии у них менее совершенных суставных блоков (Барсболд, 1976а; Osmolska, Roniewicz, 1970; Osmolska, Roniewicz, Barsbold, 1972). Присутствие в кисти у орнитомимид нешкивной структуры запястья (у дейнохейрид соответствующие кости пока не найдены) дозволяет достаточно полно характеризовать тип кисти у этих теропод.
Неплохим дополнением к сказанному служит кисть у тираннозаврид. Она отличается двупалостью, иллюстрирующей дальнейшее углубление специализации передней конечности. Однако эта конечность подвержена еще и прогрессирующей репродукции, приведшей к ее ослаблению. Кисть остается нехватательной и в ее основе, как отмечалось, лежит нешкивная структура запястья. Кисть у тираннозаврид служит примером сохранения примитивных черт при общей специализации (Барсболд, 1976а).
Обе модификации кисти у хищных динозавров, безусловно, отражают специфику развития их ветвей в различных адаптивных зонах. Нешкивная структура, по-видимому, ближе к более примитивному исходному типу, на основе которого в дальнейшем происходило прогрессивное развитие, приведшее к высокоспециализированным вариациями хватательной кисти.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'