
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Тазовый пояс и задняя конечность
Таз. Еще в начальные периоды изучения динозавров придавалось таксономическое значение их тазу (Seeley, 1888), положенному в основу принятого и в настоящее время разделения на заурисхий и орнитисхий. В последние годы делались попытки более детального рассмотрения каждого из двух первоначально выделенных типов таза. С позиции современной науки к числу заслуживающих внимания обобщений следует отнести выделения брахиоилии и долихоилии в рамках заурисхиевого таза (Colbert. 1964). Были введены названия препубического и опистопубического таза, характерных для двух основных подразделений динозавров (Charig, 1972). Последние годы ознаменовались новыми находками в Монголии, немало расширившими существующие представления о морфологии таза у хищных динозавров. Речь идет о специфических тазовых структурах дромеозаврид и сегнозавров (Барсболд, 1976а: Перлэ, 1979: Барсболд, Перлэ, 1979; Barsbold, 1979; Barsbold, Perle, 1980). Перечисленные выше данные обосновывают разделение заурисхиевого таза у хищных динозавров на несколько основных модификаций (рис. 23, 24), отражающих главные тенденции развития группы. Однако вначале следует кратко рассмотреть и уточнить значение ряда понятий, применяемых в дальнейшем.
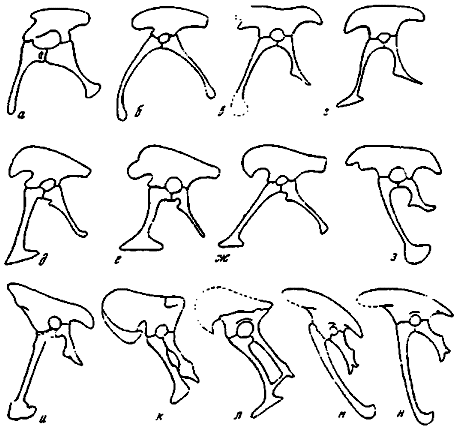
Рис. 23. Модификации заурисхиевого (преимущественно тероподового таза, вид слеза, без масштаба: а - брахиоилический таз текодонта (Colbert, 1964); препубический таз: 6 - подокезаврид (Colbert, 1912); в - заурорнитоидид; г - овирапторид; д - аллозаврид (Gilmore, 1920); е - тираннозаврид; ж - орнитомимид; з - опистопубический таз дромеозаврид; и - реконструкция таза дейнонихуса (Ostrom, 1976b); опистопубический газ: к - сегнозаврид; л - энигмозаврид; м - археоптерикса (берлинский экземпляр) ; н - реконструкция таза археоптерикса (Ostrom, 1976a)
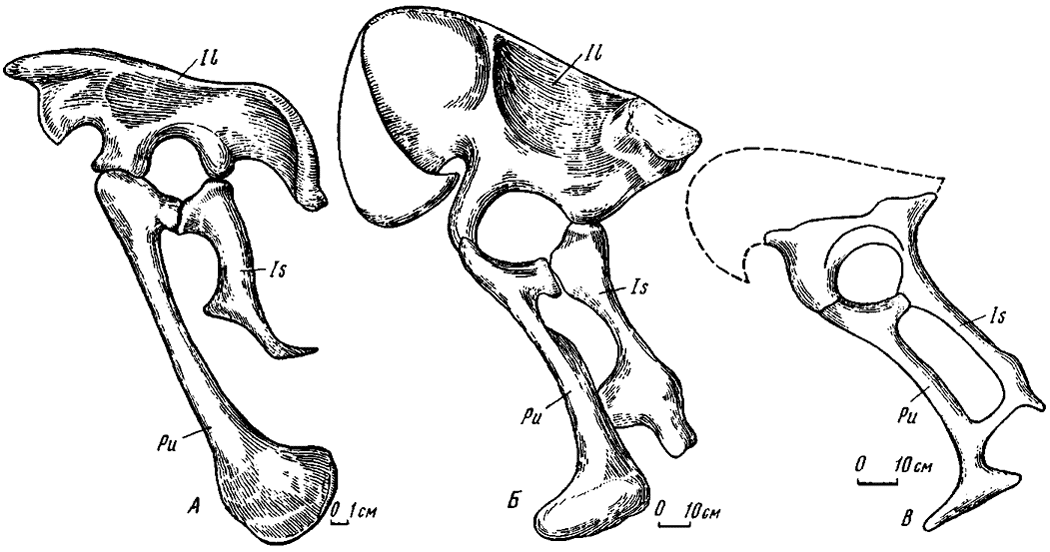
Рис. 24. Опистопубический таз слева: А - Adasaurus mongoliensis Barsbold, gen. et sp. nov. (№ 100/10); Б - Segnosaurus ghalbinensis Perle (№ 100/80); В - таз энигмозавра (№ 100/84)
Брахиоилию определяет преимущественно укороченность переднего фланга подвздошной кости, ярко выраженная у наиболее древних групп хищных динозавров (Colbert, 1964), которые не входят в состав подотряда Theropoda. В таком случае брахиоилическими являются не только ранние хищные динозавры и зауроподы, но и некоторые поздние текодонты. Представители же собственно теропод обладали долихоилией, т.е. удлиненным передним флангом подвздошной кости, характерным для самых различных их групп. Таким образом, брахиоилия и долихоилия дают возможность демаркировать наиболее древние группы хищных динозавров и устанавливать более приемлемую основу для выделения среди них собственно тероподовых и нетероподовых линий.
Названия препубический и опистопубический были введены при рассмотрении особенностей заурисхиевого и орнитисхиевого типов динозаврового таза с целью обобщенного определения ориентации лобковой кости (Charig, 1972). Однако появление нового монгольского материала дает основание употреблять эти названия в рамках только лишь заурисхиевого таза хищных динозавров (Barsbold, 1979). Такое применение терминов способствует их большей конкретизации, отражая действительное положение вещей - переднюю и заднюю ориентацию лобковой кости в заурисхиевом типе таза. Вместе с тем необходимо подчеркнуть, что подмеченные ранее морфологические особенности динозаврового таза зачастую рассматривались изолированно. На самом деле картина, вероятно, гораздо сложнее и выражается в различных комплексах признаков структур заурисхиевого таза. Корреляции этих признаков пока еще улавливаются с трудом, однако ныне совершенно ясно, что заурисхиевый (трехлучевой) таз хищных динозавров и ряда других архозавров подразделяется на препубическую и опистопубическую формы. В рамках обоих типов целесообразно выделение долихоилии, брахиоилии, а также совершенно специфического явления - альтиилии (Barsbold, 1979). При этом брахиоилия, как отмечалось, встречается у хищных динозавров и поздних текодонтов раннего этапа эволюции, не относящихся к собственно тероподовым линиям. Следовательно, последние характеризовались тремя основными модификациями таза: препубической и опистопубической долихоилией и альтиилией, возможно, связанной только лишь с опистопубической формой таза. Ниже следует рассмотрение этих модификаций (рис. 23, 24).
Препубическая долихоилия. Нормальный тероподовый таз, наиболее широко распространенный у более прогрессивных хищных динозавров, обладал ярко выраженной долихоилией и обычной передне-нижней ориентацией лобковой кости. Такой таз был свойствен подокезавридам, целуридам и т.д. и ряду более поздних ветвей - тираннозавридам, орнитомимидам, овирапторидам, заурорнитоидидам. Подвздошная кость с удлиненными передними и задними флангами, относительно высокая у крупных и низкая у мелких теропод. Передние фланги за исключением концевых краев вплотную прилегают к не выступающим выше них остистым отросткам крестцовых позвонков, в заднем же направлении подвздошные кости постепенно расходятся. Передний край подвздошной кости двураздельный, как у тираннозаврид или же цельный, как у орнитомимид и овирапторид. Задний фланг имеет различную конфигурацию, обычно более суживается, чем передний. Лобковая ножка более удлинена, чем седалищная. Лобковая кость с удлиненным стволом и дистальным окончанием различной формы, чаще передне-заднеудлиненной, или башмаковидной. У тираннозаврид дистальное образование очень крупное, продольно сильно удлиненное, массивное. У орнитомимид и особенно овирапторид дистальное башмаковидное образование сильно уменьшается и приобретает более тонкие очертания. В передне-дорзальной части дистального утолщения присимфизальный край узкий. Седалищная кость в разной мере короче лобковой, иногда она относительно длинная, выпрямленная, с небольшим проксимально смещенным обтюраторным отростком, что свойственно тираннозавридам и орнитомимидам. Кроме того, встречается укороченная седалищная кость со слегка выгнутым вперед стволом и крупным, расположенным на середине ее длины обтюраторным отростком, характерная для овирапторид и заурорнитоидид.
Опистопубическая долихоилия (рис. 24). Эта модификация была впервые выявлена на примере таза у монгольских дромеозаврид, в частности, у адазавра и велоцираптора, а также у других родственных видов и служит яркой характеристикой семейства (Барсболд, 1976а). Она отличается от нормального типа прежде всего задне-нижней направленностью лобковой кости, обосновывающей ее большое своеобразие. Вместе с тем подвздошная кость по общему очертанию вполне сходна с более низким вариантом таковой у нормального, т.е. препубического долихоилического таза. Край переднего фланга двураздельный, задний же отличается необычной утолщенностью. Лобковая кость с удлиненным стволом и округленным и сжатым с боков дистальным окончанием, составляющим, помимо обратной ориентации кости, еще одну ее отличительную особенность. Седалищная кость значительно короче лобковой, с дистально смещенным крупным обтюраторным отростком, как и в случае препубической долихоилии.
Альтиилия (рис. 24 Б, В). К этой модификации отнесен своеобразный тип; крупного таза, впервые описанный у монгольских сегнозавров (Перлэ, 1979; Барсболд, Перлэ, 1979; Barsbold, 1979). Подвздошная кость с сильно расширенным, высоким, крыловидно наружно отогнутым передним флангом, конфигурация и размеры которого совершенно необычны для таза хищных динозавров. Задний фланг сильно укорочен, что также несвойственно для других теропод. Отогнутость переднего и сокращенность заднего флангов подвздошных костей в тазу заурисхиевого типа до сих пор была известна лишь у зауропод.
В верхне-заднем крае альтиилического таза присутствует крупный массивный кубовидный выступ, служивший, по всей вероятности, местом прикрепления бедренно-хвостовых мышц и тем самым предположительно компенсировавший редуцированность заднего фланга. Подвздошные кости широко расставлены, т.е. находятся на значительном расстоянии от остистых отростков крестцовых позвонков, выступавших выше дорзального края подвздошных костей. Лобковая ножка сравнительно удлинена. Лобковая кость с удлиненным стволом и крупным дистальным окончанием, сжатым с боков, передняя часть которого была удлинена, задняя сильно укорочена. Отмечается также другая разновидность дистального окончания в виде некрупного, довольно низкого башмаковидного утолщения передне-верхний присимфизальный край которого с продольным широким углублением. Седалищная кость короче лобковой, с крупным, сильно выступающим обтюраторным отростком, соединявшимся или прочно сраставшимся с лобковой костью выше ее дистального конца. Задний край кости напротив обтюраторного отростка с отчетливо выраженной пологой выпуклостью. Дистальный конец седалищной кости в разной степени расширенный, прямой.
Таким образом, заурисхиевый таз у хищных динозавров претерпевал большие изменения. Брахиоилия, характерная для наиболее ранних стадий развития группы и свойственная также поздним текодонтам (Colbert, 1964), свидетельствует не только об их генетической общности, но также в определенной мере отражает начальные этапы становления специфического "трехлучевого" заурисхиевого таза, ставшего основой "для дальнейших преобразований. Следующим этапом в этом направлении было развитие нормальной, или препубической долихоилии, на более поздних этапах определявшей многообразие форм тазовых структур у различных групп теропод. Общепринятое" до сих пор представление о заурисхиевом тазе хищных динозавров основывается именно на подобной долихоилии. Как и нормальная долихоилия, брахиоилия находила свое выражение в препубической форме газа, обычной среди теропод. Однако выявляется еще одна форма - опистопубическая, ставшая известной в рамках заурисхиевого - таза лишь недавно. У дромеозаврид поворот лобковой кости произошел без видимых существенных изменений в остальных структурах, т.е. без нарушения долихоилии. Однако альтиилия представляет собой уже совершенно новую модификацию, отразившую глубокие морфологические изменения опистопубического заурисхиевого таза хищных динозавров. Вместе с тем альтиилия показывает определенное сходство подвздошных костей у специфических теропод и зауропод, тем самым, возможно, свидетельствуя в пользу единства происхождения подобных структур у обеих крупных групп заурисхий.
Бедренная кость. В большинстве случаев бедренная кость в разной степени длиннее голени, лишь у тираннозаврид немного короче. Для поздних теропод наиболее характерна сигмоидная изогнутость бедренной кости - ее ствол в сагиттальном плане изогнут выпуклостью вперед. Головка проксимального эпифиза с ясно обозначенной шейкой направлена медиально к телу кости и к мыщелкам дистального эпифиза. Это приводит к тому, что бедренная часть ориентирована в парасагиттальной плоскости, что связано с "нерасставленной" позицией задних конечностей. Вследствие этого вес тела передавался через дорзальный край ацетабулярного отверстия таза на суставной блок бедренной кости. Нерасставленность проподиев ограничивала их движение главным образом вертикальной плоскостью. Большой трохантер располагался на одной линий с головкой и рельефно слабо обособлен от продолжения дорзальной поверхности шейки. Впереди большого трохантера, отделяясь от него вертикально направленной щелью, находится малый трохантер в виде выпуклого утолщенного ругозистого гребня, проксимально достигающего одинакового с большим уровня. Наружная поверхность большого трохантера образует круто нисходящую расширенную ругозистую площадку. На медиальной поверхности ствола кости несколько ниже суставной головки располагается четвертинный трохантер в форме продольного гребня или нередко слабой выпуклости, отмеченной ругозистостью. У теропод четвертичный трохантер обычно смещен проксимально и находится на уровне четвертой или пятой части длины бедренной кости, что в норме считается характерным для бипедальных групп. Наибольшее отклонение фиксируется у овирапторид, у которых четвертый трохантер располагается почти на половине длины кости, при этом он у них выражен наиболее слабо. Дистальный эпифиз с развитыми мыщелками, межмыщелковая борозда глубокая и узкая, чаще слабо выражена на передней поверхности эпифиза, которая может быть уплощенной, как у овирапторид или значительно вогнуто-выпуклой вследствие сильного развития межмыщелковой борозды, как у тираннозаврид. Передняя поверхность дистального эпифиза у орнитомимид отличается от всех указанных форм - дистальный край без мыщелковой борозды, вместе с тем средняя часть эпифиза продольно-выпуклая, вдоль его медиального края протягивается глубокая борозда, отсутствующая у других теропод, сам же край развит в виде сильного уплощенного ругозистого гребня. Этот медиальный краевой гребень сильно удлинен у тираннозаврид, короткий и выпуклый у овирапторид, слабо выражен у дромеозаврид.
Голень. Большая берцовая кость значительно массивнее малой, с прямым стволом и расширениями на концах, ориентированными в субперпендикулярных плоскостях. Проксимальный конец более расширенный и утолщенный, с отогнутым кнемиальным гребнем. Проксимальная суставная поверхность состоит из двух выпуклых мыщелков, из них медиальный удлинен в передне-заднем направлении, латеральный - округлый, сильно выступающий наружу. Ниже него на уровне верхней трети или четверти длины всей кости развит высокий продольный гребень, к которому прилегает малая берцовая кость. Дистальное расширение большой берцовой кости характеризуется утолщенностью, сходящей на нет к латеральному краю, что обусловлено спецификой мезотарзального соединения.
Малая берцовая кость по длине немного уступает большой, с дистально сильно суженным стволом. Проксимальное расширение утолщено и слегка вогнуто изнутри. В проксимальной трети или четверти длины кости находится гребневидная ругозистая выпуклость (прикрепление малого разгибателя стопы), ниже которой утончение ствола становится более равномерным.
Мезотарзальное сочленение. Для теропод, равно как и для всех динозавров, характерно мезотарзальное соединение, свойственное архозаврам и многим лепидозаврам (Romer, 1956). Однако у теропод структура мезотарзального соединения отличается высокой специфичностью. У поздних линий это простая и компактная структура, образованная двумя проксимальными (астрагал и пяточная кость) и дистальными тарзальными элементами. Тело астрагала имеет форму асимметричного цилиндра, что характерно для теропод, и крайне высокий восходящий отросток. Пяточная кость, как правило, сильно редуцирована, плотно соединена с астрагалом и ее роль в мезотарзальном соединении, по-видимому, носит подчиненный характер. Оба проксимальных тарзальных элемента представляются как бы насадками с округленными суставными поверхностями, неподвижно установленными на дистальных концах костей голени. Такая структура обеспечивает жестко ограниченное в степенях свободы, направленное движение (Bakker, Galton, 1974; Charig, 1972: Krebs. 1965), важное при быстрой локомоции по твердому субстрату. Кроме того, мезотарзальное соединение связывается с пальцехождением, нерасставленной позицией конечностей и бипедализмом (Charig, 1972), характерными для теропод.
Мезотарзальное соединение у теропод, археоптерикса и птиц принципиально сходно. Вместе с тем характер его строения немало отличается от разных модификаций мезотарзального соединения у других заурисхий, а также орнитисхий. Мезотарзальное соединение у прозауропод и зауропод носило более сложный характер, как и у большинства текодонтов. Астрагал подвижно соединялся с пяточной костью, а также с большой берцовой посредством проксимального выступа, входившего в соответствующее углубление (Charig, Attridge, Crompton, 1964; Hughes, 1963). У орнитисхий астрагал без восходящего отростка, хотя и плотно сращен с пяточной костью.
У псевдозухий и современных крокодилов астрагал и пяточная кость, обладающая хорошо развитым бугорком для прикрепления икроножной мышцы, соединены подвижно (Romer, 1956; Bakker, Gallon, 1974). Структурно пяточная кость входит в состав стопы, тогда как астрагал функционально относится к голени в противоположность тероподам, у которых обе кости функционально относятся к голени. Псевдозухиево-крокодиловый тип соединения не ограничивал подвижность, и его характер свидетельствует в пользу стопохождения при частично или широко расставленных конечностях (Charig, 1972; Ostrom, 1976a). Этот тип послужил исходным для построения мезотарзального типа (Walker, 1964, 1972). Однако против этого свидетельствует то, что трудно включить пяточную кость в состав голени, чтобы не изменить первичных функций приведения и отведения стопы (Krebs, 1963, 1965, 1973). Сейчас стали известны псевдозухии лагерпетон и лагозухус (Romer, 1971, 1972), у которых развито мезотарзальное соединение, в силу чего вопрос об исходном типе ставится несколько иначе. На ранней стадии эволюции заурисхий допускается развитие примитивного прозауроподового типа мезотарзального соединения с отсутствием восходящего отростка астрагала, отмечаемого у единичных, отнесенных к тероподам триасовых заурисхий (Huene, 1942; Raath, 1969; Bakker, Galton, 1974).
Стопа. Можно выделить три основных типа строения стопы. Первый представлен нормальной тероподовой стопой, характеризовавшейся, как нередко отмечалось ранее, "птичьим" строением, хотя привычное название группы и означает "звероногие". Второй тип был обнаружен сравнительно недавно и представляет собой специфическую структуру с хищническим когтем. Третий тип, не совсем обычный для хищных динозавров, известен у сегнозавров.
Нормальная стопа (рис. 25). У поздних теропод стопа морфологически четырехпалая, функционально же скорее трехпалая. Она характерна для большинства групп и ее многочисленные вариации свойственны тираннозавридам, орнитомимидам, овирапторидам, целуридам и т.д. Стопа подобного типа в общем действительно сходна с птичьей компактной сжатой плюсной и расходящимися пальцами с заостренными когтевыми фалангами.
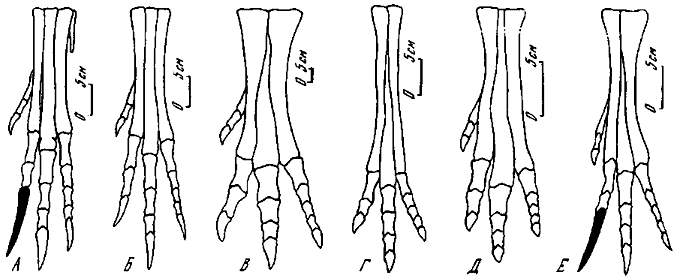
Рис. 25. Модификации нормальной стопы и стопы с отгибающимся вторым пальцем (вид спереди, схематизировано, хищнический коготь зачернен): А - велоцираптор; Б - овираптор; В - тарбозавр; Г - галлимимус; Д - гарудимимус; К - урорнитоидес
Плюсна состоит из трех вполне развитых (II, III, IV) метатарзальных и двух (1 и V) рудиментарных костей. Распознаются две модификации в ее строении. В первой из них III метатарзальная кость почти не испытывает проксимального сжатия, во второй она сильно сжата. В первом случае эта кость принимает немалое участие в формировании проксимальной суставной поверхности метатарзуса, тогда как во второй она практически исключалась из сустава. Соответственно 1 и IV метатарзальные кости в проксимальной части или полностью разделялись III метатарзальной, или же соединялись. Плюсна первой модификации наиболее ярко представлена у овирапторид и аллозаврид, ее отличает, кроме того, умеренная удлиненность или даже некоторая укороченность. Плюсна же у тираннозаврид и орнитомимид служит лучшим примером структуры с проксимальной редукцией III метатарзальной кости, ей также свойственно значительное удлинение. Вместе с тем, как выяснилось недавно, у более примитивных монгольских орнитомимид (гарудимимид) третий метатарзальный элемент практически не был проксимально сжатым и характеризовался значительной укороченностью. Дистальные головки основных метатарзальных костей, как правило, характеризуются выпуклой суставной поверхностью. I и V метатарзальные кости представлены лишь рудиментами соответственно проксимальной и дистальной частей. При этом первая кость несет редуцированный палец, а пятая полностью лишена его. У прогрессивных орнитомимид в противоположность примитивным (гарудимимидам) I метатарзальная кость полностью отсутствует (Барсболд, 1981).
Фаланговая формула стопы 2-3-4-5-0. Первый палец был существенно недоразвитым, не достигавшим субстрата. Нередко его считали противопоставленным, или обращенным назад по отношению к остальным пальцам, что, безусловно, неправильно. Сочленовная поверхность остаточного метатарзалъного элемента совершенно ясно опровергает возможность такого положения, более характерного настоящей птичьей стопы. Второй, третий и четвертый пальцы вполне развиты и служили опорой через третий палец, иногда бывший более удлиненным. Фаланги четвертого пальца наиболее укорочены. Когтевые фаланги довольно однообразны по форме. У первого пальца они сжаты с боков, у остальных вентрально несколько уплощены, при этом наибольшая степень уплощения характерна для орнитомимид.
Стопа с отгибающимся пальцем (рис. 26, 27). Впервые стала известна у американского дромеозаврида - дейнонихуса (Ostrom, 1969 а, b) и была неоднократно описана на монгольском материале, добавившем сюда еще и заурорнитоидид (Barsbold, 1974).
И в данном типе возможно выделение двух модификаций метатарзуса: с почти несжатым третьим элементом, характерным для дромеозаврид и с сильно сжатым, свойственным заурорнитоидидам. В обоих случаях метатарзус не отличался значительной удлиненностью. К числу существенных особенностей относится хорошо разработанный дистальный гинглимоидный составной блок II метатарзальной кости, тогда как третья и четвертая обладали выпуклыми суставными головками.
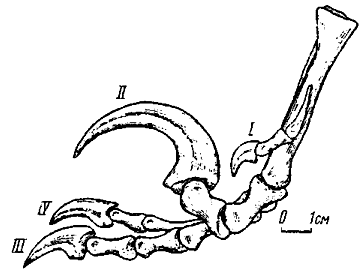
Рис. 26. Правая стопа дромеозаврид с отгибающимся вторым пальцем и хищническим когтем (изнутри, реконструкция с учетом данных Ostrom, 1969b)
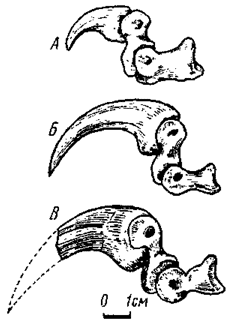
Рис. 27. Отгибающийся второй палец правой стопы с хищническим когтем, вил изнутри: А - Adasaurus mongoliensis (№ 100/20); Б - Velociraptor mongoliensis (№ 100/25); В - неопределенный монгольский дромеозаврид (№ 100/12)
В фаланговой формуле стопы изменений нет. Вместе с тем ее второй палец отличается крайне специфическим строением. Его фаланги заметно укорочены и базальная принимает участие в формировании гинглимоидного сустава со своей метатарзальной костью - на проксимальной суставной поверхности она несет срединный гребень, в норме отсутствующий у этого пальца. Дистальный блок с сильно развитыми и обращенными дорзально гинглимоидными дугами. Наиболее отклоняется строение предкогтевой фаланги - се проксимальная суставная поверхность с сильно развитой вентральной частью, языковидно вытянутой назад значительно дальше уровня укороченного и глубокого вырезанного дорзального края. Вследствие этого формируется специфический сустав, непохожий на обычный, обеспечивавший сильное отгибание назад предкогтевой фаланги. Этому способствовала и дорзальная обращенность гинглимоидных дуг базальной фаланги. Когтевая фаланга сильно заострена и сжата с боков, она почти в два-три раза превосходит по длине обычные когтевые фаланги. По всей вероятности, палец с хищническим когтем не участвовал в локомоции и в обычных условиях находился в отогнутом состоянии, не касаясь субстрата. Третий и четвертый пальцы были примерно одинаковы по длине и осуществляли опорную функцию при передвижении. Ось тяжести тела проходила между этими пальцами, существенно не отличавшимися по остальным признакам от одноименных пальцев нормальной тероподовой стопы.
Стопа третьего типа (рис. 28) известна у монгольских сегнозавров (Перлэ. 1979; Барсболд, Перлэ, 1979). Она существенно отличается от двух других типов. В метатарзус сегнозаврового типа входит не менее четырех элементов, V метатарзальная кость рудиментарна. Метатарзус отличается укороченностью, относительной утолщенностью составляющих, практически не выраженной компактностью строения, что сразу выводит его за рамки птичьего типа. I метатарзальная кость относительно крупная, расширенная, чего нет у других модификаций стопы, ее проксимальная часть с глубокой латеральной вогнутостью, которой она соединялась со смежным элементом. Дистальные суставные головки выпуклые, расширенные. Фаланговая формула не изменяется. Первый палец был наиболее укороченным, однако мог касаться субстрата. Остальные пальцы более или менее близки по длине, ось тяжести проходила через третий палец. Крайне специфично присутствие в четвертом пальце двух сильно укороченных, проксимально-дистально уплощенных фаланг, ранее встречавшихся у других хищных динозавров. Когтевые фаланги в различной степени сжаты с боков.
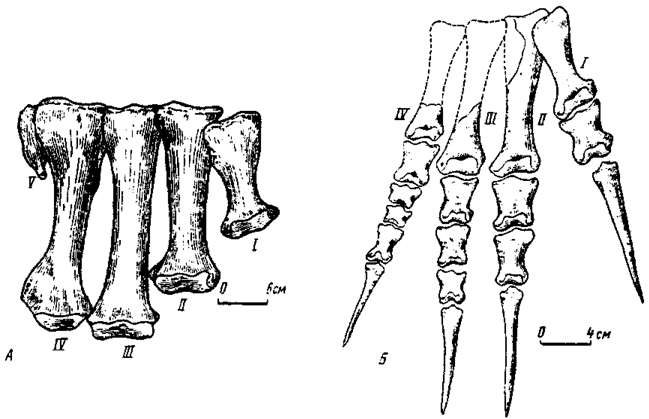
Рис.28. Стопа сегнозавров спереди: А - правый метатарзус Segnosaurus Perle (№100/80); Б - правая стопа (частично реконструированная) Erlikosaurus andrewsi Perle (№100/111)
Заключение. Три типа стопы у хищных динозавров различаются между собой столь существенно, что функциональные возможности каждой из них должны быть достаточно специфическими. Нормальная функционально трехпалая стопа поздних теропод представляет собой закономерное осуществление тенденции к сокращению пальцев исходной пятипалой стопы более ранних групп. Во второй половине мезозоя вырабатывается компактная стопа птичьего строения, в наиболее специализированной форме известная у тираннозаврид, прогрессивных орнитомимид и т.д. Проксимальное сжатие III метатарзальной кости и удлинение метатарзуса следует рассматривать, как дальнейший шаг в достижении совершенной опорной структуры, необходимой для быстрого передвижения по субстрату. У некоторых форм развивалась стопа близкого строения, но без проксимальной сжатости III метатарзального элемента, что свойственно более ранним и более примитивным группам (аллозавриды, гарудимимиды).
Стопа с отгибающимся пальцем происходит от нормальной стопы, приобретшей чрезвычайно специфическое новообразование в виде модифицированного второго пальца с хищническим когтем. С его приобретением стопа становится орудием защиты и нападения (Ostrom, 1969 а, b), что, вероятно, нисколько не ухудшило способности к маневренному и быстрому передвижению. Площадь соприкосновения с субстратом даже уменьшилась вследствие исключения пальца с хищническим когтем.
Стопа сегнозаврового типа принципиально отлична от двух других. Прежде всего, она некомпактна и укорочена, что свидетельствует в пользу иного, вероятно, менее совершенного способа локомоции. Эта стопа сходна с таковой у ряда орнитопод, к примеру, у гадрозавров, что подтверждается не только общим сходством этой структуры у названных групп, но и дисковидно уплощенными и укороченными фалангами пальцев. Поэтому есть основания предполагать, что стопа у сегнозавров связана с примерно сходным амфибиотическим образом жизни, по-видимому, не требовавшем совершенного, легкого и быстрого бега.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'