
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Функциональные особенности посткраниальных структур
Шейные позвонки. Естественный изгиб шеи в немалой степени зависит от размеров и, следовательно, веса головы, а также длины и гибкости шейного ряда позвонков (рис. 29). Вместе с тем с характером изгиба связана различная подвижность определенных участков шейных позвонков.
С увеличением и утяжелением черепа относительная длина шеи уменьшается и в этом плане тираннозавриды, пожалуй, не имели себе равных. Это выражается в сокращении не числа позвонков, а их длины. При наибольшем укорочении шея у тираннозаврид приближалась к вертикали, что связано с резко различным уровнем передних и задних суставных поверхностей позвонков. Ясно, что более вертикальное положение шеи отвечает несению и большей нагрузки. Шея образует естественный изгиб сигмоидного характера, и к вертикали приближается выпрямленный участок между изгибами. Судя по укороченности первых позвонков, сигмоидный характер естественного изгиба был сравнительно слабо выражен, и шея в целом была мало гибкой.
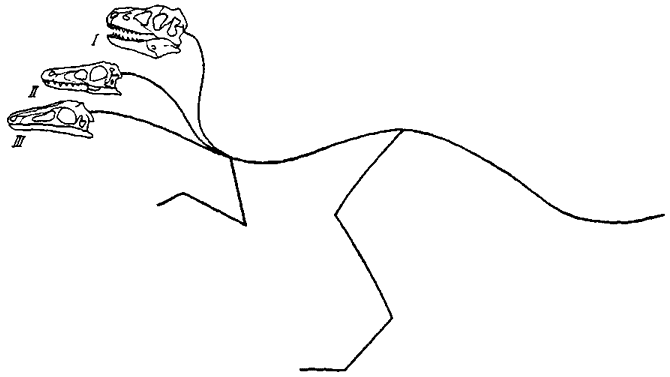
Рис. 29. Предполагаемая поза тела и постановка головы (хвост показан не полностью): I - крупного хищного динозавра типа тираннозаврид; II - умеренно крупного или мелкого хищного динозавра типа дромеозаврид; III - умеренно крупного беззубого динозавра типа орнитомимид
Тираннозавриды резко отличались от орнитомимид, обладавших удлиненной и гибкой шеей. Очень малый череп у орнитомимид создавал незначительную нагрузку на его предположительно мускулистую шею. Судя по положению суставных поверхностей, естественный изгиб шеи соответствовал пологой сигмоиде, при этом передние и последние позвонки, бывшие достаточно удлиненными, ориентировались более горизонтально и между ними протягивался основной участок шеи, сопрягавшийся с ними по изгибам дуги с большим радиусом. Предполагается, что шея у орнитомимид была более гибкой. Возможность большей подвижности выражается в том, что у передних позвонков презигапофизы отличаются существенной удлиненностью, что допускает повороты и вращение головы и шеи с большей амплитудой. Суставные фасетки зигапофизов ориентированы под большим углом к сагиттальной плоскости позвонков, что приближает их к горизонтали и способствует боковым движениям начального участка шейного ряда. Это явление отмечается и у других групп теропод с относительно удлиненной шеей. Выпрямленный участок сигмоиды между изгибами несет зигапофизы с фасетками, обращенными несколько медиально, что способствует вращательным движениям вокруг продольной оси. Задние позвонки показывают некоторое сокращение длины презигафизов и более горизонтальное положение их суставных фасеток, что в большей мере отвечает сгибанию и разгибанию и боковым движениям шеи, нежели вращательным. Это относится и к укороченной шее тираннозаврид, но подобные черты в удлиненной шее у орнитомимид выражены гораздо более отчетливо. Как показывают материалы монгольской коллекции, орнитомимиды достигали гигантских размеров, почти не уступая тираннозавридам. При этом череп их по мере увеличения размеров тела пропорционально сильно уменьшался. Шейные ребра были достаточно развитыми и прочно прикреплялись к позвонкам, что свидетельствует в пользу сильной мускулатуры, значительно превосходившей потребности поддержания головы. Развитие шейных мышц, по-видимому, отражает усиление гибкости и подвижности шеи у орнитомимид, частично сходных в этом отношении с длинношеими бегающими птицами. Другие группы - дромеозавриды, овирапториды - по структуре шеи и ее ориентировке занимают некоторое промежуточное положение между короткошеими тираннозавридами и длинношеими орнитомимидами.
Спинные позвонки. Серия спинных позвонков формирует более или менее горизонтальный ряд, точнее, слабо выпуклую арку (в противовес традиционным реконструкциям бипедальных динозавров, якобы опиравшихся хвостом о субстрат), что дополнительно доказывает расположение суставных поверхностей позвонков строго на одном уровне. Суставные фасетки зигапофизов, обращенные более медиально, обусловливают в большей мере способность к сгибанию и разгибанию спины в сагиттальной плоскости, чем к значительным боковым изгибам на всем протяжении спинного ряда. Кроме того, присутствие у дромеозаврид и тираннозаврид межпозвонкового соединения типа гипосфен-гипантр (Ostrom, 1969 а, b; Малеев, 1974), равно как и развитие мощных межкостных связок, места прикреплений которых найдены по краям остистых отростков практически у всех теропод (а также крокодилов), свидетельствуют о значительном общем ограничении подвижности в спине. Подобные связки хорошо развиты у современных страусов, благодаря которым позвонки, особенно на заднем участке спины, превращены в прочный, малоподвижный стержень, направленный горизонтально (Ostrom, 1969 b).
Хвостовые позвонки. Поперечные отростки у преобладающего большинства теропод развиты в передней части хвоста, что имеет место и у млекопитающих, тогда как у рептилий они прослеживаются на большем протяжении всего хвостового ряда позвонков (Ostrom 1969 b). Развитие поперечных отростков связывают с присутствием прикреплявшихся к ним боковых сгибателей хвоста. Кроме того, в передней части хвоста у наземных позвоночных всегда развиты дорзальные и вентральные мышцы, игравшие разную роль, но в основном служившие поднимателями и опускателями хвоста (Miller, 1969). У ящериц выделяют дорзальные разгибатели в области остистых отростков и вентральные сгибатели, расположенные вблизи гемальных отростков. Эти мышцы у теропод группировались преимущественно в передней части хвоста, о чем, в частности, свидетельствует большое развитие здесь невральных и гемальных дуг позвонков. У большинства теропод наиболее "активная" часть хвоста занимала существенно меньше передней половины его длины. Вместе с тем развитие зигапофизов и шевронов почти на всем протяжении хвоста достаточно ясно показывает, что и задний его отдел также обладал определенной межпозвонковой подвижностью. Удлинение и сближение презигапофизов, приподнятых к дорзальной поверхности позвонков, вероятно, увеличивало жесткость конструкции хвоста. Для всех позвонков характерно расположение суставных поверхностей на одном уровне, что отражает, по-видимому, или отсутствие, или незначительность его естественного изгиба.
Овирапториды отличаются значительным развитием поперечных отростков на значительной части хвоста. Кроме того, остистые отростки у них существенно удлинены, что обычно характерно для хвоста водных рептилий (Romer, 1956). Поперечные отростки, как отмечалось, связаны с развитием боковых сгибателей, и их усиление у овирапторид, возможно, связано с превращением хвоста в плавательный орган. Боковые сгибания хвоста в такой плотной среде, как вода, требуют не только большой силы от сгибателей; он должен активно преодолевать сопротивление среды, для чего сгибатели должны протягиваться вдоль возможно большего числа позвонков. Только при выполнении таких условий функционирование хвоста будет достаточно эффективным. Возможно, укорочение тел позвонков, свойственное овирапторидам, а также гадрозаврам, способным к плаванию, связано с усилением межпоперечных хвостовых мышц, прикреплявшихся через один - два позвонка к поперечным отросткам. В результате увеличивается частота прикрепления мышц на единицу длины, что обеспечивает выигрыш в силе.
Структура хвостовых позвонков у дромеозаврид представляет собой крайнюю степень специализации, выраженную в развитии окостеневавших нитей, в которые как бы продолжаются презигапофизы и шевроны. Плотно упакованные костные тяжи придают хвосту жесткость, вероятно, не достигаемую обычными средствами. Вместе с тем допускается ограниченная межпозвонковая подвижность вследствие того, что окостеневавшие нити не срастались между собой, позвонки хорошо обособлены друг от друга даже в дистальной части хвоста и нет слияния шевронов с телами позвонков. Окостеневавшие нити связываются с сухожилиями хвостовых сгибателей и разгибателей (Ostrom, 1969 а, b). Подниматели и опускатели также группировались преимущественно в основании хвоста. Следовательно, упруго жесткая хвостовая структура у дромеозаврид по подвижности подчинялась главным образом переднему отделу. Подобный специфический орган напоминал по своей упруго жесткой конституции стек, разве что гораздо более массивный, и при нормальном передвижении занимал, по-видимому, более или менее горизонтальное положение, во всяком случае, находился на весу. Дж. Остром допускает, что высокий инерционный момент хвоста коррелятивно связан с подвижностью и маневренностью дромеозаврид и способствовал резким поворотам при беге.
Общность структуры хвоста у теропод показывает приспособление к выполнению его главной функции - балансирования при бипедальном передвижении. Понятно, что такое функционирование требует прежде всего более или менее горизонтального положения, точнее, положения на весу самого балансира, служившего противовесом предкрестцовой части осевого скелета. На этой основе развиваются различные специфические приспособления, рассмотренные выше. Разнообразие функций и положение на весу хвоста у теропод приходит в противоречие с привычным представлением о нем, как о третьей опоре тела бипедального животного, широко принятом в традиционных реконструкциях динозавров. Впрочем, данное заключение далеко не ново: так, А. Ромер (Romer, 1956) уже писал о хвосте бипедальных динозавров, прежде всего, как о балансире при локомоции.
При бипедальном передвижении наиболее выгодными были дифференцированная подвижность и гибкость осевого скелета. Действительно, опорный участок его, представленный крестцом, прочно связанным с тазом, был неподвижным. Спинные позвонки как бы образуют конструкцию вынесенной фермы, поддерживающей внутренности и ориентированной по горизонтали от своего неподвижного основания (Ostrom, 1969 b). Боковые изгибы нецелесообразны при двуногохождении и спина была способна лишь к ограниченному сгибанию и разгибанию. Путем сопряжения по дуге осуществляется переход от туловища к гибкой и подвижной шее. Балансом, служившим также противовесом предкрестцовой части тела, был хвост, обычно державшийся на весу. Вполне возможно, что реконструкции галопирующих динозавров Т. Баккера (Bakker, 1975) не во всем являются беспочвенной фантазией, и, по крайне мере, тероподы были действительно подвижными двуногоходящими животными, хвост которых и в покое не всегда служил лишь пассивной опорой.
Общая ориентировка осевого скелета у теропод, определяющая постановку тела, представляется следующей; в различной степени сигмоидально изогнутая шея, от сравнительно укороченной, приближенной к вертикали до удлиненной пологой, подвижность которой соответственно увеличивалась; почти горизонтально расположенные, ограниченно подвижные спинные позвонки, переходящие в компактный крестец, представлявший неподвижную основу осевого скелета; наконец, гибкий хвост, в большинстве случаев державшийся на весу и обладавший разной степенью инерционности.
Передняя конечность. Структура передней конечности у теропод ясно показывает общность основы, приспособленной к выполнению интенсивных и разнообразных движении. Однако у разных групп передняя конечность нередко весьма отлична. В этом плане одним из лучших примеров служит развитие дельтовидного гребня плечевой кости, достигавшего у многих групп теропод одинаково высокого уровня. Это свидетельствует в пользу значительного развития грудных мышц, обусловливавших интенсивность и силу движений передних конечностей у таких хищников, как дромеозавриды, овирапториды и, особенно, теризинозавриды. Как известно, грудная мускулатура в совокупности с другими мышцами плечевого пояса выполняет существенную роль в приведении и сгибании передней конечности. Сильно развитыми были и мышцы, обеспечивавшие обратные движения, что подтверждается, например, усилением трехглавой мышцы, следы прикрепления которой особенно заметны у теризинозаврид и других перечисленных выше хищников с сильным развитием грудной мускулатуры. Об этом свидетельствует присутствие широкой грудины у дромеозаврид и овирапторид.
Передние конечности у теропод обладали довольно значительной подвижностью, определяемой достаточно совершенным строением плечевого сустава. Гленоидная впадина обычно глубокая и обращена наружу, суставная головка плеча хорошо выражена и все это вместе взятое обеспечивало выполнение довольно многообразных движений, в том числе и вращательных по длине оси конечности. У групп с усиленной функцией передних конечностей характерны значительное дистальное расширение плечевой кости и обращенность мыщелков вперед, что определяет согнутость конечностей в локтевом суставе в спокойном положении.
По всем рассматриваемым аспектам орнитомимиды, дейнохейриды и тираннозавриды во многом составляют противоположность вышеперечисленным группам. У тираннозаврид это объясняется крайней редукцией передних конечностей, ослабившей и ограничившей, но не устранившей полностью их функций. У орнитомимид крупные, а у дейнохейрид даже гипертрофированные передние конечности кажутся непропорционально ослабленными и потому гораздо менее эффективными в силе и интенсивности движений. Суставные соединения у групп с ослабленной функцией передних конечностей представляются менее совершенными, менее способными к вращательным движениям, в том числе и вокруг оси; в спокойном положении передние конечности были более выпрямленными, слабо эффективными в выполнении функций приведения и отведения, сгибания и разгибания, в особенности, сопряженных с большими усилиями.
Большое значение в определении функций передних конечностей обычно придается кисти. При этом шкивной характер запястья отчетливо определяет функциональные возможности данной структуры. Главный вид движений кисти в таком запястье определяется шкивной бороздой. Амплитуда бокового движения кисти в запястье почти в точности определялась отрезком окружности, занятой рабочей поверхностью шкива, и во всех случаях эта дуга не превышала 180°. Сгибание и разгибание кисти также определялись строением шкивной борозды, в основном ее шириной, отвечавшей степени свободы движений. Шкивной механизм при своей простоте и упорядоченности входящих в него структур обеспечивал разнообразную фиксированную подвижность в запястье (Ostrom, 1969 b, 1974 а).
Нешкивной механизм запястья уступает рассмотренному в совершенстве как структуры, так и производимых движений. Он существенно менее компактен, соответственно его подвижность носит менее фиксированный, менее упорядоченный характер.
Трехпалость, типичную для поздних теропод, вероятно, следует рассматривать как наиболее хорошо приспособленную для специфического способа "хватания". Возможно, такое количество пальцев отражает собой наилучшее решение, при котором "хватательная" способность кисти держится на достаточно высоком уровне. Само "хватание" у теропод было совсем не таким, как у млекопитающих с брахиальной локомоцией - приматов и отчасти неполнозубых (ленивцев). В отличие от них, у теропод при так называемом "хватании" скорее всего доминировал ударно-прокалывающий захват, имеющий большее сходство с действием когтистых лап задних конечностей хищных птиц. Лишь в этом специфическом смысле можно рассматривать кисть у теропод как "хватательную", точнее, захватывающую, или зацепляющую, структуру. Подобная способность кисти наиболее ярко выражена в строении пальцев, расходящихся при разгибании, снабжении их заостренными когтевыми и удлиненными предкогтевыми фалангами, что характерно и для некоторых хищных птиц (Ostrom, 1969 b). Противопоставление первого пальца кисти остальным неизвестно у теропод, хотя именно об этом писалось в связи с ее "хватательной" способностью (Osborn, 1917). Явственное же расхождение первого пальца связано с медиальным обращением дистального блока метакарпальной кости. Гинглимоидные суставы фаланг пальцев не допускают обхватывающего сгибания (Ostrom, 1969 b; Osmolska, Roniewicz, 1970), характерно для кисти лазающих животных.
В рамках "хватательного" типа находится, как отмечалось, место и для кисти дромеозаврид, теризинозаврид и овирапторид, столь развивающихся по строению и размерам пальцев. Кисть у дромеозаврид, бесспорно, наилучшим образом соответствовала этому типу. Заостренность и загнутость их когтевых фаланг, по-видимому, делает ударно-прокалывающий захват особенно эффективным. При достаточной изогнутости когтя заостренный конец при сгибании предкогтевой фаланги все более обращается назад, способствуя более глубокому вонзанию и, тем самым, более прочному захвату. В прямой связи с большим изгибом когтевых фаланг находится и степень развития бугорков для прикрепления сильных сгибателей пальцев; одновременно, когтевые фаланги сильно сжаты с боков.
Гигантские, уплощенные с боков когтевые фаланги в кисти у теризинозаврид едва ли были приспособлены к прокалывающему захвату, как у дромеозаврид. Вместе с тем укорочение фаланг пальцев указывает на возможное приспособление к нанесению удара гигантскими когтями (передние конечности у теризинозаврид, по-видимому, отличались силой).
Другое отклонение встречается у специализированных овирапторид - ингений, у которых кисть в целом сохраняет черты "хватательного" типа, однако необычно сильная редукция второго и третьего пальцев должна была уменьшать их участие в основной функции, отводя им скорее роль опор, удерживающих кожистую перепонку. У более примитивных овирапторид - собственно овирапторов - пальцы кисти находятся в рамках "хватательной" нормы.
Кисть у теропод с ослаблением функций передних конечностей, имея в основе "нехватательный" тип строения, также обладала значительным разнообразием. Слабо изогнутые когтевые фаланги у орнитомимид обладали мало рельефными бугорками для прикрепления сгибателей, что составляет прямую противоположность структурам дромеозаврид. С учетом ослабленности передних конечностей и малой сжатости когтевых фаланг можно вполне уверенно считать, что кисть у орнитомимид совсем не была приспособлена к ударно-прокалывающему захвату. У тираннозаврид общая редукция передних конечностей сильно ограничивала и функции кисти, лишенной к тому же третьего пальца. Нарастание редукции передних конечностей, прослеживаемое и у более ранних крупных хищников - аллозаврид и мегалозаврид и достигшее максимума у тираннозаврид, также происходило в рамках нешкивного запястья, обеспечивавшего меньшую эффективность в "хватании". Кисть у дейнохейрид имеет ряд как бы противоположных черт, выражающихся в общем ослаблении передних конечностей и отклонении кисти от "хватательного" типа в связи с развитием крупных массивных изогнутых когтевых фаланг. По всей вероятности, такая кисть могла производить лишь в общей форме те движения, которые требовались бы при исполнении ударно-прокалывающего захвата. Крупные передние конечности обладали немалым инерционным моментом, но по мощности мышц и степени приспособленности структуры в целом к такому захвату дейнохейриды сильно уступали теризинозавридам. Вероятно, кисть у дейнохейрид в большей или меньшей мере сохранила возможность произведения в какой-то форме ударных движений, хотя эффективность их едва ли была особо высокой. Вместе с тем, форма ударных движений передних конечностей не всегда связана с обязательным нападением на жертву, и можно найти немало других сфер их применения, как раз не требующих особой силы, к примеру, хотя бы раскапывание термитников и т.д. (Рождественский, 1970).
Задняя конечность. Структура задней конечности у теропод отражает общность ее основы в приспособлении к бипедальной локомоции. Хорошо оформленная головка бедренной кости, обращенная медиально и ориентированная почти перпендикулярно к длинной оси ствола, показывает передачу основной нагрузки вдоль этой оси и отражает нерасставленную постановку конечностей. Коленный изгиб также проходил в парасагиттальной или близкой ей плоскости, следовательно, ось головки бедренной кости была перпендикулярна (или близка к этому) плоскости сгибания в коленном суставе (Ostrom, 1969b). Такое положение проподий свойственно обычно конечностям птиц и млекопитающих. Зафиксированные в породе следы динозавров также вполне свидетельствуют в пользу подобной постановки конечностей и у этих животных (Ostrom, 1969b, 1972; Bakker, Galton, 1974). Известно, что нерасставленная позиция конечностей представляет собой одно из немногих анатомических признаков в пользу эндотермии динозавров (Ostrom, 1969b, 1972; Bakker, 1971a; Bakker, Galton, 1974).
У бипедальных теропод четвертый трохантер в большинстве случаев значительно смещен проксимально, что связано с увеличением роли бедренно-хвостовых ретракторов, по-видимому, выигрывавших в скорости за счет силы вследствие укорочения плеча рычаг, что существенно для подвижных форм. Толчок при шаге передавался через конечность на головку бедренной кости не в медиальном направлении, как бывает при горизонтальном положении бедра, а на верхний край вертлужной впадины (Bakker, Galton, 1974), что одновременно создает лучшие предпосылки для поддержания веса тяжести. В этом случае парасагиттальное положение бедра повышало эффективность движения конечности при отталкивании. Ранее считалось, что бипедальная локомоция дает выигрыш в расходовании энергии перед четвероногохождением, однако, как оказалось, этого не происходит и выигрыш состоит в большей маневренности (Bakker, 1972; Bakker, Galton, 1974) . Вся структура задних конечностей у теропод, совершенных бипедальных животных, была направлена на достижение максимума в этом выигрыше. Это находит отражение и в строении стопы, функционально трехпалой, первоначально приспособленной к передвижению по более или менее твердому субстрату с опорой на пальцы. Характерно, что стопа у большинства теропод отличается компактностью и суженностью, что свойственно современным птицам и многим млекопитающим. Специализация стопы выражена у ряда хищных динозавров; так у прогрессивных орнитомимид отсутствует первый палец, плюсна испытывает пропорционально наибольшее удлинение, наблюдаемое среди теропод. Эти черты наравне с вентральным уплощением когтевых фаланг стопы свидетельствуют о приспособлении к быстрому бегу. Однако стопа других групп - дромеозаврид и заурорнитоидид - имеет резкие отличительные особенности из-за развития отгибающегося второго пальца с хищническим когтем (Ostrom, 1969a, b; Barsbold, 1974). Предкогтевая фаланга с этим когтем могла сильно отгибаться вследствие специфического строения сустава между ней и базальной фалангой, т.е. палец мог как бы убираться назад и в спокойном состоянии не касаться субстрата своим заостренным, сильным удлиненным когтем. Стопа в локомоторном отношении становилась двупалой, и это, возможно, было одновременно приспособлением к быстрому маневренному бегу, так как площадь опоры стопы при этом сокращается. Вместе с тем стопа приобрела дополнительную функцию, как эффективное орудие нанесения ударов, раздирающих тело жертвы или врага. Допускается, что удар "хищническим" когтем наносился одной конечностью, в то же время как вторая выполняла функцию опорную. Это предполагает высокую степень равновесия и подвижности животного (Ostrom, 1969b). Однако более вероятно, что когти обеих задних конечностей пускались в ход одновременно, для чего хищник должен был повиснуть на жертве, уцепившись за нее передними лапами, или же вместе с ней упасть на спину, что делают в схватке некоторые кошачьи. В уникальной находке сцепившихся скелетов хищного велоцираптора и растительноядного протоцератопса положение задних конечностей хищника свидетельствует в пользу именно такого предположения (Барсболд, 1974).
Заключение. Естественный изгиб шеи немало зависит от размеров и, следовательно, веса головы, а также длины и гибкости шейного ряда позвонков. Увеличение черепа сказывается на уменьшении относительной длины шеи, ее ориентировка все более приближается к вертикали. При уменьшении черепа происходит обратное явление - удлинение шеи и ее ориентировка по пологой сигмоиде.
Спинные позвонки формируют ограниченно подвижный ряд, определявший постановку тела теропод, которое не было наклонено назад и вниз с упором на хвост, как принято при традиционных реконструкциях бипедальных динозавров. У большинства теропод наиболее активная часть хвоста занимала меньше передней половины его длины. В определенной мере это также противоречит тому, чтобы считать хвост преимущественно опорным органом. Вместе с тем у овирапторид "активность" хвоста чрезвычайно усилена практически на всем его протяжении, что отвечает функции как плавательного органа. У дромеозаврид наблюдается резкое усиление жесткости и упругости хвостового ряда позвонков, что превышает его в совершенный динамический балансир.
В определении функции передней конечности немалая роль принадлежит шкивному или нешкивному характеру кисти, с чем связано усиление или ослабление передней конечности в целом и степень совершенства ее "хватательности".
Структура задней конечности отражает единство ее основы в приспособлении к бипедальной локомоции, совершавшейся при нерасставленной постановке задних конечностей, обычно свойственной птицам и млекопитающим.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'