
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Линии развития теропод
Изучая пути эволюции теропод, необходимо различать общие тенденции, осуществлявшиеся на разной основе, от общих же, развивавшихся на единой основе. Именно с этой точки зрения разделение палеопод и теропод, равно как и выделение различных линий внутри теропод, приобретают большую обоснованность. Общее направление в эволюции теропод, обусловленное их генетическим родством, было более перспективным, чем у палеопод, не перешедших границы триаса. Тероподы же в течение большей части мезозоя, развиваясь по ряду параллельных линий, оказались способными к осуществлению множества узких специализаций, приведших на поздних стадиях эволюции к обширному многообразию. Поток тероподовой эволюции имел отклонения, обусловленные сдвигом в особые адаптивные зоны, примером чему служат сегнозавры. Тем не менее именно благодаря этому многообразию все явственней обрисовывается единство всех линий тероподового ствола, рано обособившегося от палеопод (рис. 30).
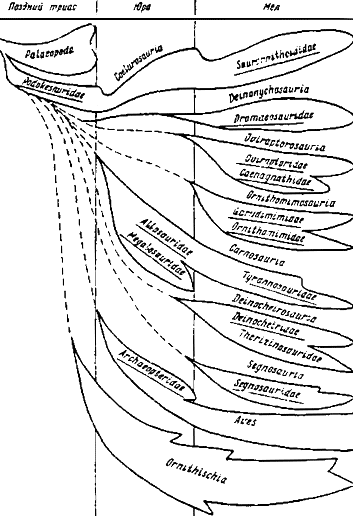
Рис.30. Филогения основных групп хищных динозавров и их предполагаемые связи с орнитисхиями и птицами
Ветви целурозавров (подокезавриды, компсогнатиды, целуриды, заурорнитоидиды, элмизавриды, авимимиды) составляют единственную линию из известных ныне среди теропод, перешедшую границу триаса и юры и продолжавшую свое дальнейшее восхождение до конца мезозоя в направлении приобретения все более узких специализаций. Эволюция целурозавров после триаса происходила скорее всего в рамках той же довольно обширной адаптивной зоны, в которой она началась. Это подтверждается тем, что послетриасовые целурозавры, в общем продолжающие одну и ту же эволюционную линию, что и ранние ветви, лишь глубже осуществили те морфологические тенденции, которые уже были заложены на более примитивном уровне еще у подокезаврид. Так, целуриды, а после них и элмизавриды, оставались мелкими хищниками с более глубокой редукцией боковых пальцев стопы и кисти, чем подокезавриды, у которых это явление лишь намечалось. В позднем мезозое линия целурозавров испытывает радиацию, наметившую основу для дальнейшей дивергенции на новые ветви и линии более узко специализированных форм. К началу мела целуриды вымирают, и на эволюционной сцене их место занимают другие группы, развивавшиеся в направлении параллельных специализаций с приобретением и углублением ряда усовершенствований. Так, авимимиды представляют линию с еще большим приближением строения плюсны к птичьему типу. Заурорнитоидиды формируют другую линию с полостью среднего уха (как у птиц) и отгибавшимся пальцем стопы с хищническим когтем. Вероятно, это связано с выходом заурорнитоидид в новую адаптивную зону, где они имели преимущество перед менее специализированными поздними целурозаврами. Вместе с тем, если не принимать в расчет указанных усовершенствований, то заурорнитоидиды отличаются от целурид разве только лишь усилением ранее отчетливо намеченных специализаций, характерных для всех целурозавров.
Линия дейнонихозавров известна пока в мелу; она характеризовалась рядом особенностей, обусловивших ее сходство с целурозаврами. Однако глубокие отличия в строении опистопубического таза свидетельствуют о ее более раннем заложении. Вместе с тем дейнонихозавры с приобретением параллельных целурозавровым специализаций, наиболее ярким свидетельством чему служит присутствие отгибавшегося второго пальца стопы с хищническим когтем. В совокупности с тазом, в котором имело место обращение лобковой кости, эти существенные преобразования отражают развитие дейнонихозавров в адаптивной зоне, немало отличной от целурозавровой. Среди дейнонихозавров на наиболее поздних стадиях развития наблюдается сильная редукция хищнического когтя стопы до размеров обычных, хотя специфическая структура фаланг полностью сохраняется. Дейнонихозавры дивергировали на несколько ветвей, различавшихся деталями в строении стопы, а также относительной высотой черепа. Все это показывает расщепление дейнонихозавров по близким параллельным направлениям в конце мела.
Овирапторозавры - следующая линия высокоспециализированных мелких теропод, генетически предположительно связанных с целурозаврами. Наряду с чертами, свидетельствующими о достаточно раннем эволюционном сдвиге, овирапторозавры несут в посткраниальном скелете немало признаков целурозавровой организации, подвергшихся значительной модификации. Овирапторозавры, как и рассмотренные выше дейнонихозавры, демонстрируют известное явление "мозаичной" эволюции, когда с несомненно прогрессивными признаками соседствуют многие черты, отражающие более примитивный характер. У овирапторов наибольшие изменения, в противовес дейнонихозаврам, затронули череп. Отсутствие зубов, присутствие массивного клюва, как у цератопсий и значительные модификации в строении небного комплекса и нижней челюсти отражают переход этих своеобразных хищников в адаптивную зону, вероятно, не свойственную для большинства теропод. Первично "хватательная" тероподовая трехпалая кисть, возможно, преобразовывалась в плавательный орган. По-видимому, овирапторозавры смогли осуществить прорыв в действительно новую адаптивную зону и пройти в число первых амфибионтов среди теропод. Вместе с тем не совсем ясно положение с ценагнатидами, вероятно, они были ветвью, параллельной овирапторидам и развивавшейся в несколько иной адаптивной зоне.
Перейдем к рассмотрению развития крупных теропод. Уже отмечалось, что в эволюции собственно карнозавров (мегалозаврид, цератозаврид, аллозаврид, тираннозаврид) явственно прослеживаются тенденции как общего тероподового, так и крайне специфического характера. Однако общие черты, связанные с усовершенствованием бипедализма и плотоядностью, совсем не обязательно должны свидетельствовать об их прямой филогенетической связи с собственно целурозаврами. Как показано ранее, на фоне осуществления "тероподовых" тенденций собственно карнозавры выделялись столь существенными специфическими чертами, как длительно прослеживаемая редукция передних конечностей, присутствие нешкивного типа кисти, определенное увеличение черепа, укорочение шеи и т. д. Все это предполагает довольно раннюю дивергенцию карнозавров вследствие эволюционного сдвига и заметного отхода от линий мелких теропод. С позиций осуществления эволюционных тенденций собственно карнозавров кажется вполне обоснованным рассматривать мегалозаврид в широком смысле (т. е. включая цератозаврид и аллозаврид) и тираннозаврид как два этапа в эволюции единой линии, направление развития которой вполне сохранялось в течение всего позднего мезозоя. В этом отношении сменявшие друг друга ветви карнозавров находятся в одинаковом положении с линией собственно целурозавров, также составлявших крупный ствол с приобретением внутри параллельных специализаций. Карнозавровые особенности получают у тираннозаврид, главным образом, лишь более глубокое выражение, чем у мегалозаврид. Нараставшая редукция передних конечностей и все предположительно связанные с этим явлением корреляции в структурных комплексах карнозавров отражают развитие этих хищников в направлении гигантизма, проявлявшегося уже на ранних стадиях их эволюции.
Орнитомимозавры - вторая группа беззубых теропод, которых ранее относили к целурозаврам в широком смысле, т. е. к мелким тероподам. Представленные известным семейством орнитомимид, а также гарудимимидами эти беззубые хищники в своем строении отражают как бы соединение "целурозавровых" и отчасти "карнозавровых" тенденций: с одной стороны, небольшой череп, удлиненная шея, нередуцированные передние конечности, с другой - таз, построенный по "карнозавровому" типу, примитивное нешкивное запястье. Доминирующим признаком, вероятно, следует считать все же строение препубического долихоилического таза, поскольку другие структурные комплексы, достаточно жёстко сцепленные друг с другом, могли подвергаться значительной модификации, как это демонстрируют все тероподы позднего этапа эволюции. Поэтому предполагается, что орнитомимозавры могли иметь более близкие связи с крупными тероподами, нежели с мелкими. Направление столь своеобразной специализации должно было установиться достаточно рано, и развитие орнитомимозавров шло по пути все большего ее усиления, о чем свидетельствует полная утрата первого пальца стопы у наиболее продвинутых из них, тогда как у примитивных он еще присутствует. Явление сходной специализации, отличающейся лишь глубиной выражения, показывает эволюцию двух параллельных ветвей орнитомимозавров, существовавших одновременно, в позднем мезозое Монголии. Мнения о систематическом положении орнитомимид были противоречивыми - их издавна относили к мелким тероподам (Matthew, 1915), и эта точка зрения считается общепринятой. Однако были и попытки отнести их к крупным тероподам (Abel, 1919). По-видимому, орнитомимозавры представляли рано обособленную линию, в большей мере отвечавшую условиям адаптивной зоны, развитие в которой способствовало гигантизму. Поздние представители специализированных монгольских орнитомимид по величине почти не уступали своим современникам - тираннозавридам. С другой стороны, несмотря на нередуцированность (в смысле уменьшения в размерах) передних конечностей, их функциональное ослабление ставит орнитомимозавров ближе к карнозаврам, чем к мелким тероподам. Другие яркие признаки, свойственные орнитомимозаврам, свидетельствуют в пользу их развития в ином направлении, чем собственно карнозавры. Допустимо считать, что эти структурные особенности - малый череп, удлиненная шея и передние конечности и т. д. развивались параллельно таковым у разных групп мелких теропод при сходных требованиях среды и, естественно, на основе генетической общности тероподового ствола.
Рассмотрение следующей линии крупных теропод - дейнохейрозавров, условно включающих дейнохейрид и теризинозаврид, наталкивается на ряд трудностей из-за отсутствия более полных морфологических данных. Однако можно с уверенностью утверждать, что они как крупные хищники развивались без редукции передних конечностей, осуществляя общую тероподовую тенденцию формирования трехпалой кисти. У теризинозаврид усиленная передняя конечность построена на основе шкивного механизма запястья, тогда как у дейнохейрид структура кости отвечала более примитивному нешкивному типу, сходному с орнитомимидным, и характеризовалась функциональной ослабленностью. Таким образом, передние конечности дейнохейрозавров вполне соответствовали двум структурным модификациям тероподового характера, вместе с тем существенно выделяясь развитием крупных или даже гигантских когтей, отражающих их крайне узкую специализацию. Рассматриваемые признаки дейнохейрозавров заставляют предполагать, что они развивались более или менее параллельно линиям теропод с нередуцированными передними конечностями. Однако дейнохейрозавры осваивали специфическую адаптивную зону, значительно отличающуюся от собственно карнозавровых, равно как и от зон мелких теропод. Можно предположить, что дейнохейрозавры должны были рано встать на путь самостоятельного развития. Среди крупных теропод они далеко отстоят от собственно карнозавров и орнитомимозавров, хотя дополнительные данные в будущем могут привести к совсем другим заключениям.
Линия сегнозавров ныне представляется, пожалуй, наиболее специфической. Ее представители плохо поддаются даже условному отнесению к крупным или мелким тероподам хотя бы на основании общих размеров. В рамках общетероподовых тенденций сегнозаврам свойственны черты наибольшего отклонения, лучше всего выраженные в альтиилии тазового пояса. Вместе с тем обращенность лобковой кости свидетельствует о частично сходных перестройках, произошедших в тазу у дейнонихозавров и сегнозавров, однако на этом их сходство заканчивается. Если у первых переориентировка произошла в рамках долихоилии, то у вторых таз претерпевает коренные изменения. По-видимому, это означает не только довольно раннее обособление сегнозавров от общего потока тероподовых линий, но и существенный сдвиг в сторону от основных направлений их эволюции. Сегнозавры развивались в условиях крайне узкой адаптивной зоны, возможно, не характерной в целом для теропод. Модификации в строении сегнозаврового таза показывают выделение, по крайней мере, двух линий, вероятно, осуществлявших параллельные специализации.
Подведем основные итоги. Накопившиеся данные достаточно ясно свидетельствуют против "двухлинейной" - только карнозавры и целурозавры - позиции в представлениях об эволюции теропод. Эти рамки кажутся уже слишком тесными и пересмотр можно сделать, базируясь преимущественно на материале по поздним стадиям развития теропод. Нет оснований сомневаться в том, что дальнейшее изучение раннего этапа принесет еще немало нового. Пока же ясно одно - взаимоотношения внутри теропод в эволюционном плане были гораздо более сложными и многообразными, чем представлялось в рамках традиционных понятий о двух "линиях". Новые данные позволяют обосновывать концепцию о многолинейном параллельном развитии хищных динозавров.
С самого начала появления заурисхий тероподы встали на путь усовершенствования бипедализма. Хищные палеоподы в этом направлении не имели таких перспектив вследствие "инадаптивности" существенно важных структур, определявших бипедальную локомоцию. Морфологическая эволюция палеопод совершалась в определенной мере параллельно развитию теропод. Единый ствол теропод расщеплялся на ряд линий, осуществлявших в течение позднего мезозоя параллельные специализации, обусловленные сравнительно ранним выходом представителей этих линий в узкие адаптивные зоны. Некоторые из них отличались крайним своеобразием, приведшим к существенному отклонению таких групп, как сегнозавры. Несмотря на специфичность, приобретаемые адаптации совершались на единой структурной основе, не заслоняемой всем многообразием модификаций у поздних линий. Последний этап развития теропод характеризуется ускорением эволюции, наблюдаемым на примерах дейнонихозавров, орнитомимозавров и овирапторозавров при одновременном параллельном развитии внутри каждой из групп более близкородственных ветвей. Наряду с параллелизмами более общего тероподового характера, они подчеркивают многоярусность явлений параллельной эволюции. Специализации на позднем этапе отличались большей углубленностью, соответствовавшей условиям новых адаптивных зон, по которым широким фронтом распределялись достигшие относительно высокого прогресса поздние тероподовые линии.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'