
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Подкласс Polymera. Полимеры
К подклассу полимер, или многочленистых (гр. poly - много), относятся все остальные трилобиты с числом туловищных сегментов от 5 и выше (трилобиты с 4 туловищными сегментами неизвестны). Подкласс объединяет свыше 1400 родов. На основании особенностей строения головного и хвостового отделов, характера лицевых швов подкласс разделен на 7 отрядов: Olenellida, Redlichiida, Corynexochida, Ptychopariida, Phacopida, Lichida, Odontopleurida.
Отряд Olenellida. Оленеллиды (Olenus - один из героев греческой мифологии, превращенный в камень) - наиболее примитивные многочленистые трилобиты. Тело длинное, уплощенное; головной щит относительно крупный; щеки снабжены щечными шипами; глабелла с ясно выраженными бороздами. Глаза, как правило, крупные, с глазными крышками; лицевые швы метапариевого типа; передняя ветвь направлена к щечному углу. Туловище состоит из непостоянного, но обычно большого числа (12-44) сегментов, несущих сильные борозды и шипы, а третий сегмент часто увеличен и несет особенно крупный шип. Хвостовой отдел очень маленький, нерасчлененный, почти целиком занят осевой частью или шиповидным тельсоном. Ранний кембрий. Около 20 родов (рис. 122, 1).
Отряд Redlichiida. Редлихииды (названы в честь К. Редлиха, описавшего фауну кембрийских трилобитов Индии в 1901 г.) близки к предыдущему отряду, отличаются крупными глазами с заднещечными лицевыми швами. Глабелла четко отделена от щек; подвижные щеки с длинными щечными шипами. Туловище состоит из многочисленных сегментов (15-25); хвостовой отдел маленький. Ранний и средний кембрий. Около 90 родов (рис. 122, 2).
Отряд Corynexochida. Коринексохиды (гр. koryne - булава, exochos - высокий, выдающийся). Тело удлиненное, овальное, головной щит округленный, с хорошо развитыми щечными шипами; глабелла длинная, с четкими бороздами, расширяется кпереди. Глаза меньших размеров, чем у предыдущих отрядов, узкие, с глазными валиками. Лицевые швы заднещечные. Ростральная пластинка соединена с гипостомой или рудиментарна. Туловище состоит из небольшого числа (5-12) сегментов. Хвостовой щит крупных (макропиги) или средних размеров с краевыми шипами. Ранний - поздний кембрий. Около 75 родов (рис. 122, 3).
Отряд Ptychopariida. Птихопарииды (гр. ptyche - складка, pareia - щека). Глабелла разнообразной формы; глаза от средних до маленьких, иногда отсутствуют; лицевые швы преимущественно опистопариевые, реже пропариевые; у некоторых занимают краевое положение. Гипостома отделена от цефалона гипостомальным швом. Туловище состоит из 6-25 сегментов, у многих групп их число постоянно. Хвостовой щит хорошо обособлен и состоит из небольшого числа сегментов. Появились в раннем кембрии от общих предков с редлихиидами, достигли, расцвета в позднем кембрии и ордовике и вымерли в перми. Отряд объединяет до 800 родов, разделенных на пять подотрядов: Ptychopariina, Asaphina, Illaenina, Harpina, Trinucleina.
У подотряда Ptychopariina простая, суживающаяся вперед глабелла, разделена простыми параллельными бороздами (3-4); глаза средние или маленькие, лицевые швы заднещечные; относительно большое туловище и маленький хвостовой отдел. Имеется фронтальный лимб. Кембрий - ордовик. Около 500 родов (рис. 123, 1,2).
У подотряда Asaphina (гр. asaphus - неясный) головной и хвостовой отделы почти одинаковых размеров (изопиги), туловище состоит из 6-9 сегментов. Глабелла слабо очерченная, почти без борозд; глаза на стебельках или полулунные, иногда очень крупные; лицевые швы заднещечные. Обладали способностью сворачиваться. Конец среднего кембрия - поздний ордовик, расцвет в раннем и среднем ордовике. Около - 110 родов (рис. 123, 3).
У подотряда Illaenina (лат. illaino - косоглазый) головной и хвостовой щиты почти равных размеров (изопиги); глаза полулунной формы; лицевые швы заднещечные; глабелла выражена слабо; имеется крупная ростральная табличка. Туловище из 6-10 сегментов. Возникнув в ордовике, этот подотряд пережил всех своих конкурентов и вымер в перми. Около 150 родов (рис. 123, 4, 5).
У подотряда Harpina (гр. harpe - серп) головной щит округленный с большими удлинениями щек (щечными рогами); глабелла суживается вперед, с 1-3 парами борозд; глаза развиты слабо или отсутствуют; лицевые швы краевые, реже заднещечные; туловище с 12-29 сегментами; хвостовой щит короткий. Поздний кембрий - поздний девон. Около 20 родов (рис. 123, 7).
У подотряда Trinucleina (гр. tri - три, nucleus - ядро) головной щит от округлой до треугольной формы, в несколько раз больше хвостового, окружен широким лимбом или краевой каймой. Глабелла и щеки выпуклые, с длинными щечными углами. Глаза имеются или отсутствуют; лицевые швы заднещечные или краевые. Туловище из 5-7 сегментов, редко до 30. Рахис узкий, плевры широкие. Хвостовой щит треугольный, узкий, рахис достигает конца щита; иногда сохраняются следы мест прикрепления мускулов. Ранний ордовик - поздний силур. Около 50 родов (рис. 123, 6).
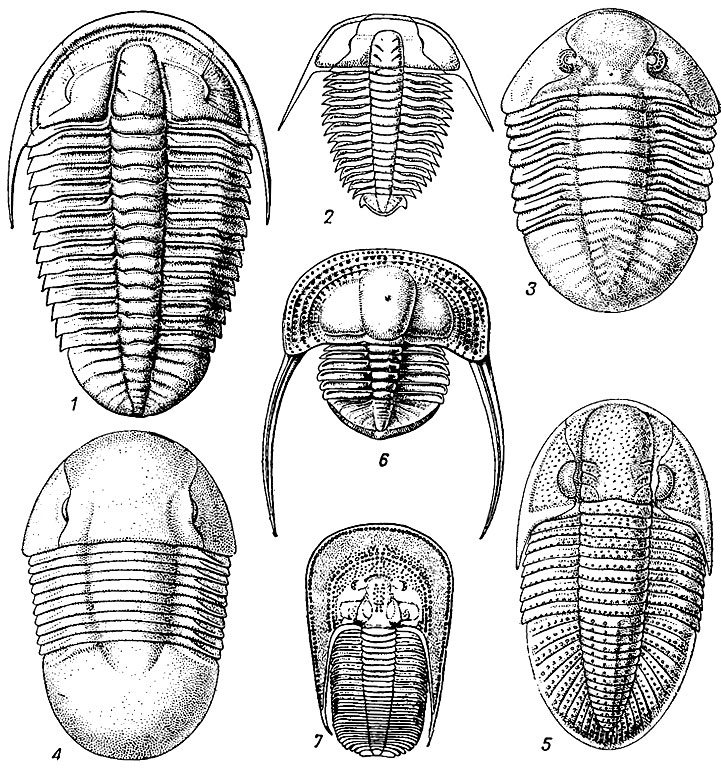
Рис. 123. Отряд Ptychopariida; 1-2 - подотряд Ptychopariina: 1 - Ptychoparia (средний кембрий), 2 - Olenus (поздний кембрий); 3 - подотряд Asaphina, Asaphus (ранний - средний ордовик); 4-5 - подотряд Illaenina: 4 - Illaenus (ордовик), 5 - Phillipsia (карбон); 6 - подотряд Trinucleina, Onnia (ордовик); 7 - подотряд Harpina, Harpes (средний девон)
Отряд Phacopida. Факопиды (гр. phakos - чечевица, ops, род. opos - глаз) - послекембрийские трилобиты мелких и средних размеров. Головной щит с крупной расширяющейся кпереди глабеллой; глаза шизохроические. Лицевые швы пропариевые или гонатопариевые, реже опистопариевые; туловище состоит из 8-19 сегментов. Хвостовой щит различных очертаний, чаще средних размеров, несет срединные или краевые шипы. Ордовик - девон. Около 180 родов (рис. 124).
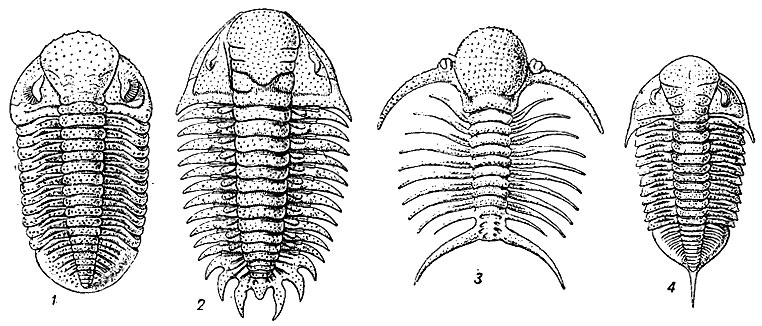
Рис. 124. Отряд Phacopida: - 1 - Phacops (силур - девон); 2 - Cheirurus (поздний ордовик - средний девон); 3 - Deiphon (силур); 4 - Dalmanites (силур - ранний средний девон)
Отряд Lichida. Лихиды (гр. Lichas - имя слуги Геракла, доставившего ему отравленный плащ) - заднещечные послекембрийские трилобиты от средних до очень крупных размеров. Головной и хвостовой щиты крупные и очень своеобразные. Глабелла широкая; лицевые швы опистопариевые. Хвостовой щит крупный, его плевральные части листовидной формы или шиповидные. Дорзальная поверхность бугорчатая. Ранний ордовик - поздний силур. 25 родов (рис. 125, 1, 2).
Отряд Odontopleurida. Одонтоплевриды (гр. odous, род odontos - зуб, pleura - бок). Мелкие трилобиты с выпуклым головным щитом, глабелла конической формы, с расширенной частью у затылочного кольца и часто с 2-3 парами боковых борозд. Глаза расположены посредине щек; лицевые швы опистопариевые. Туловище из 8-10 сегментов, причем осевая часть вздутая, выпуклая; плевральные части уплощенные. Хвостовой щит короткий, его рахис с 2-3 бороздами. Панцирь этих трилобитов имеет большое количество шипов на окончаниях почти всех сегментов (отсюда название отряда). Конец среднего кембрия - поздний девон, расцвет в конце силура - начале девона. Около 25 родов (рис. 125, 3).
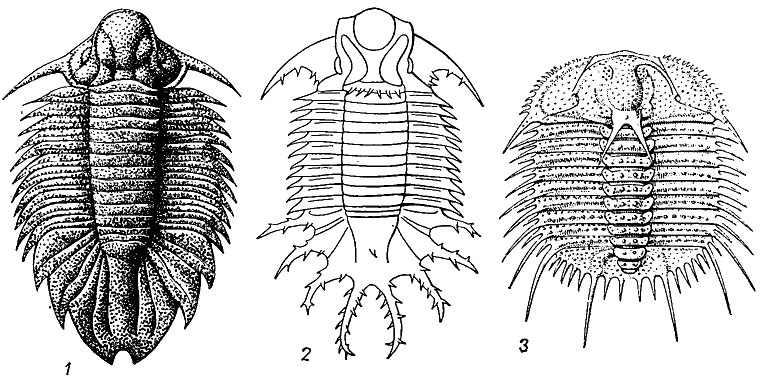
Рис. 125. Отряды Lichida (1-2) и Odontopleurida (3): 1 - Dicranopeltis (поздний ордовик - силур); 2 - Terataspis (девон); 3 - Odontopleura (средний силур)
Образ жизни и условия захоронения. Все трилобиты были морскими организмами, так как их остатки встречаются только в отложениях морского происхождения совместно с кораллами, брахиоподами, моллюсками, мшанками, граптолитами. Личинки трилобитов протаспис вели, скорее всего, планктонный образ жизни, что обеспечивало им широкое географическое расселение и адаптацию к разным экологическим нишам. После превращения в личинку типа мераспис трилобиты, по-видимому, переходили к бентосному образу жизни (рис. 126). Находки трилобитов в осадках, характерных для мелководных участков моря, - в тонких песках, глинах, глинистых и известковистых илах - позволяют предполагать, что трилобиты ползали по дну с помощью своих телоподитов - внутренних ветвей конечностей - и, возможно, плавали у дна, используя наружные ветви конечностей - преэпиподиты. Часть трилобитов зарывалась в ил, выставляя наружу глаза на стебельках, а формы, вторично утратившие зрение, всю свою жизнь проводили в илу, питаясь органическим детритом и мелкими организмами, населявшими верхние части илистого осадка.
О приспособлений к жизни на дне свидетельствует уплощенное, широкое тело, глаза на верхней стороне головного отдела, положение рта на нижней стороне, тонкая мембрана, прикрывавшая брюшную сторону, отсутствие специальных органов управления, возникающих у всех быстро плавающих форм. Бентосный образ жизни подтверждается также приуроченностью трилобитов только к определенным фациям. Дополнительными приспособлениями для жизни на илистом дне служили уплощения края головного щита - лимб и длинные шиповидные отростки щек, а также различные шиповидные отростки различных сегментов. Наличие шиповидного тельсона, подобного таковому мечехвостов, помогало этим трилобитам зарываться в осадок в поисках пищи. Мелкие трилобиты с большим количеством шипов и различных выростов, возможно, могли вести пелагический образ жизни. Об этом свидетельствуют также гипертрофированные глаза, иногда целиком занимающие поверхность щек (рис. 126, 3).
Для защиты вентральной стороны у многих трилобитов выработалась способность к свертыванию тела подобно хитонам или наземным ежам. Различают три основных способа свертывания (рис. 126, 8-10):
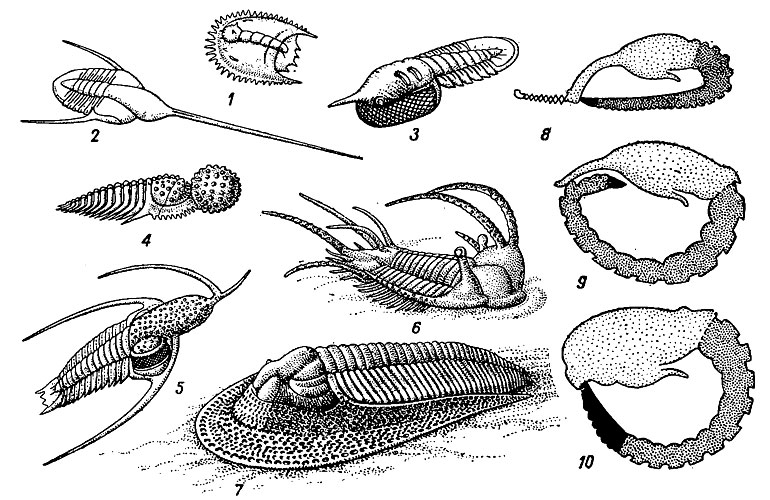
Рис. 126. Предполагаемый образ жизни трилобитов (1-5 - пелагический, 6-7 - бентосный, 8-10 - типы свертывания): 1 - личинка Acantholoma; 2 - Lonchodomas; 3 - Symphysops; 4 - Staurocephalus; 5 - Ceratarges; 6 - Teratorhynchus; 7 - Paraharpes. Типы свертывания: 8 - дискоидальный, 9 - двойной, 10 - сфероидальный; головной Щит обозначен редкими точками, туловище - частыми, хвостовой щит - зачернен
1) при сфероидальном свертывании, наиболее распространенном у трилобитов, происходило смыкание; хвостового и головного отделов, причем плевры туловища накладывались друг на друга и закрывали тело с боков, защищая конечности. Приспособлением для плотного смыкания служили так называемые "пандеровы органы". Для прочного смыкания на дублюре переднего края головного щита развивалась либо борозда (факопиды), либо система зубчиков. В борозду при свертывании упирался задний конец пигидия. "Пандеровы органы" в действительности представляют структуры наружного скелета: бугорки или валики, расположенные на дублюре щечных углов и на дублюре каждой плевры туловища. Они служили упорами для плевр при свертывании тела трилобита. Отверстия, расположенные за бугорками, относимые также к категории "пандеровых органов", служили, по-видимому, для просачивания воды к жаберным ножкам, лежащим внутри, свернутого панциря;
2) при двойном свертывании хвостовой отдел и несколько туловищных сегментов подгибались и затем происходило свертывание, при котором передний край головы соединялся с одним из тех туловищных сегментов, где происходил изгиб; пигидий и часть туловищных сегментов оказывались внутри свернувшегося трилобита;
3) при дискоидальном свертывании передний край головы соприкасался с задним краем хвостового отдела и трилобит оказывался согнутым, как книга.
Способность к свертыванию возникала у разных групп трилобитов независимо, как защитная реакция от врагов, которых в морях палеозоя у трилобитов было более чем достаточно. К числу врагов могли относиться остракодермы и рыбы, другие группы членистоногих (некоторые водные хелицеровые), головоногие моллюски: наутилоидеи, актиноцератиты, эндоцератиты. О нападении врагов свидетельствуют различные виды травм на наружном скелете, залеченные при жизни.
В ископаемом состоянии сохраняются как скелеты погибших от разных причин трилобитов, так и скелеты, сброшенные при линьке. Большинство остатков трилобитов, найденных в ископаемом состоянии, принадлежит скелетам, сброшенным при линьке. Различные швы, в первую очередь лицевые швы, затем швы между головой и туловищем, облегчали трилобитам линьку. У трилобитов без лицевых швов панцирь разрывался между головой и туловищем и в ископаемом состоянии иногда) встречаются туловище с хвостом и лежащий рядом головной щит, часто перевернутый. При наличии лицевых швов чаще всего сохраняется кранидий без подвижных щек. В бургасских сланцах среднего кембрия: Британской Колумбии (Канада) найдены остатки разнообразных членистоногих, в том числе трилобитид и трилобитов с сохранившимися остатками конечностей. Хорошо сохранились трилобиты в ордовикских известняках окрестностей Нью-Йорка и восточной части Канады.
История развития трилобитов (рис. 127). Первые трилобиты известны из алданского яруса нижнего кембрия. В отложениях самого нижнего яруса кембрия - томмотского - остатки трилобитов отсутствуют. Для расчленения кембрийских отложений используются почти исключительно трилобиты, достигавшие в кембрии большого разнообразия и изобилия. Трилобиты используются для выделения биогеографических областей и позволяют среди отложений кембрийской системы выделить отделы, ярусы и даже зоны. В раннем кембрии известны представители обоих подклассов: миомеры и полимеры. Среди первых главную роль играли Eodiscida, среди вторых Olenellida, Redlichiida, Corynexochida и первые Ptychopariida. На рубеже раннего и среднего кембрия полностью вымирают Olenellida и появляются Odontopleurida и в конце среднего кембрия первые Asaphina. В среднем кембрии становятся многочисленными Agnostida, Corynexochida, Paradoxidacea из Redlichiida, ряд семейств - из подотряда Ptychopariina.
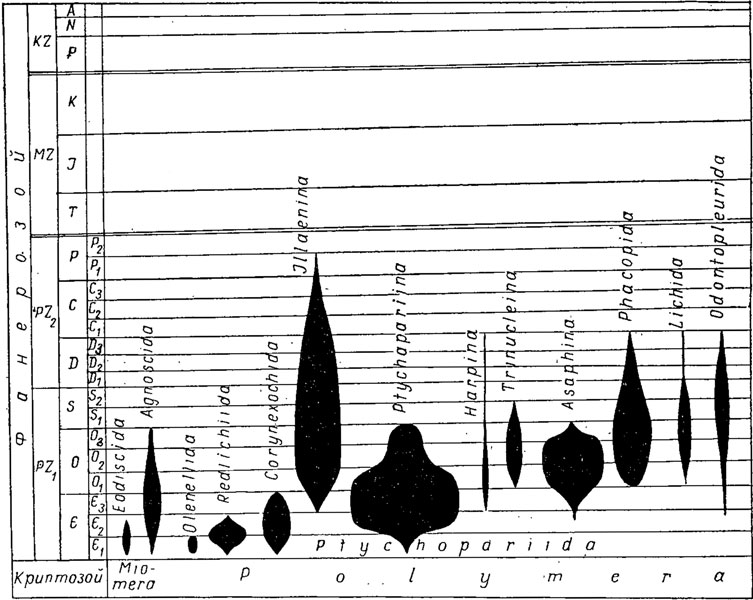
Рис. 127. Геохронологическое распространение трилобитов
На рубеже среднего и позднего кембрия вымирают Eodiscida, Redlichiida, появляются Harpina, в позднем кембрии становятся разнообразными Agnostida, Ptychopariina.
На рубеже кембрия и ордовика весьма существенно изменился систематический состав трилобитов, их значение для стратиграфии несколько уменьшилось и ведущую роль стали играть граптолиты. Вымирают Corynexochida. От птихопариид возникают два новых специализированных отряда: Phacopida и Lichida. В пределах отряда птихопариид обособляются два новых подотряда: Illaenina и Trinucleina, испытывают расцвет Asaphina. В конце ордовика вымирают Agnostida и подотряды Asaphina и Ptychopariina и резко сокращается общее число родов всех трилобитов.
В силуре значение трилобитов для стратиграфии еще уменьшилось; тем не менее среди трилобитов были распространены представители Phacopida, Harpina, Odontopleurida. В конце силура вымирают Trinucleina.
В девоне продолжали свое существование Harpina, Phacopida, Lichida и Odontopleurida, вымирающие в конце периода. Только последние представители подотряда Illaenina продолжали существовать в морях карбона и перми, причем до перми дожили только два семейства этого подотряда. В середине пермского периода, насколько можно судить по находкам, исчезают последние представители трилобитов.
В кембрийское время были обособлены три зоогеографические области: Атлантическая (Прибалтика, Скандинавия, Англия, Марокко, восточное побережье Канады), Тихоокеанская (вдоль западного побережья Северной Америки от Аляски до Мексики), Восточно-Азиатская, или краевая Тихоокеанская (от Кореи до Австралии, захватывая Индию). В каждой из областей были распространены свои характерные трилобиты.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'