
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Надотряд Autobranchia. Аутобранхии
К надотряду аутобранхий (гр. autos - самостоятельный) отнесена основная масса бивальвий, у которых жаберный аппарат преобразован в фильтр-сито, т. е. выполняет не только функцию газообмена, но и отбор мелких пищевых частиц и доставку их к ротовому отверстию. В желудке развиты сортирующие аппараты, в него глубоко вдается тифлозоль с сопровождающей его кишечной бороздой; развит кристаллический стебелек; печень открывается большим числом самостоятельных отверстий и имеет несколько дивертикул; выделяются различные типы желудков, среди которых наиболее примитивным считается униоидный, имеющий большой тифлозоль и общие протоки печени; такой тип желудка, вероятно, был характерен для всех актинодонтид. Митилоидный тип желудка имеет сложный тифлозоль; он характерен для устриц, митилин, арцин, птериин и позволяет объединять их в один отряд циртодонтид. Для остальных групп аутобранхий характерен свой тип желудка, что позволяет выделять несколько естественных групп, которым придается ранг отрядов. В пределах всего надотряда встречаются все типы раковин, замков и лигаментов, рассмотренные в общей части. По строению замка, желудка и жабр надотряд Autobranchia, распадается на восемь отрядов: Actinodontida, Pectinida, Astartida, Carditida, Venerida, Hippiiritida, Cyrtodontida, Pholadomyida.
Отряд Actinodontida. Актинодонтиды (гр. actis, род. actinos - луч, радиус) объединяют как морских, так и пресноводных бивальвий, имеющих равностворчатую раковину с перламутровым или перекрещенно-пластинчатым слоем, лиродесмоидный, актинодонтный, схизодонтный или дизодонтный замки. Жабры образованы свободными или сросшимися нитями - филаментами. Желудок униоидного типа. Большой тифлозоль имеет вид дуги, проходящей через весь желудок. Представители отряда известны с ордовика и существуют до наших дней, причем если морские формы в настоящее время немногочисленны, то пресноводные представляют процветающую группу. Отряд объединяет три подотряда: Trigoniina, Unidfliina, Actinodontina (рис. 176).
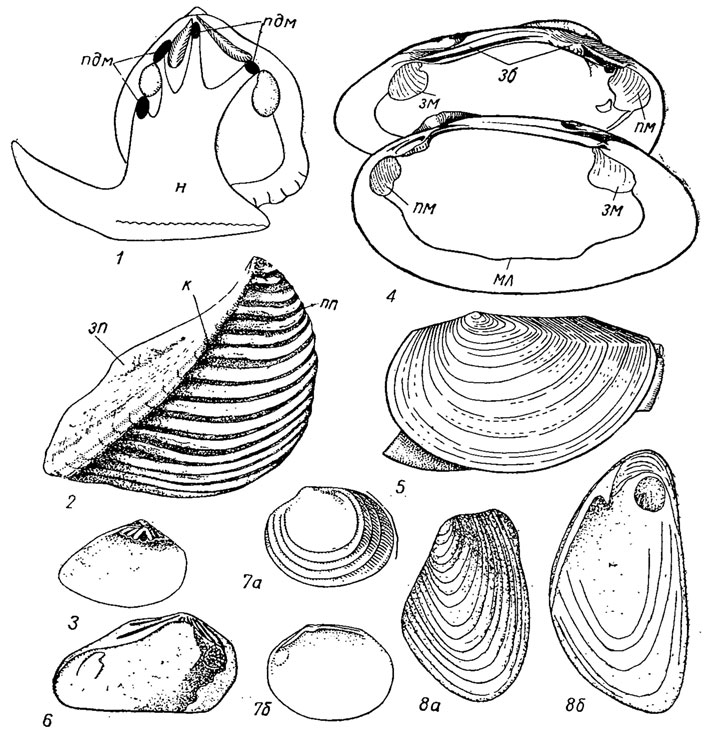
Рис. 176. Отряд Actinodontida; подотряды Trigoniina (1-3), Unioniina (4-5) и Actinodontina (6-8): 1 - Neotrigoriia (совр.), схема строения; 2 - Trigonia (средний триас - мел); 3 - Lyrodesma (ордовик); 4 - Utiio (триас - ныне); 5 - Anodonta (поздний мел - ныне); 6 - Actinodonta (ордовик); 7а, б - Cycloconcha (ордовик); 8а, б - Modiomorpha (силур - пермь); зб - зубы, зм - задний мускул, зп - заднее поле, к - киль, н - нога, пдм - педальный мускул, пм - передний мускул, пп - переднее поле, мл - мантийная линия
Подотряд Trigoniina. Тригониины (гр. triasr - три, gonia - угол) объединяют большое семейство тригониид, распространенное широко в мезозое и представленное сейчас реликтовым родом Neotrigoriia, обитающим у берегов Австралии, и два палеозойских семейства. Раковина имеет характерный схизодонтный замок, хорошо развитую скульптуру, состоящую из радиальных ребер или бугорков и киля, идущего от макушки к заднему краю и отделяющего большое переднее поле от небольшого заднего. К подотряду отнесены также семейство лиродесматид (1 род), развитое в среднем и позднем ордовике, имевшее раковину с лиродесмоидным замком, состоящим из радиально расходящихся от макушки нескольких зубов, и семейство миофорид (13 родов), развитое от девона до конца триаса (рис. 176, 1-3). Подотряд существует с ордовика.
Подотряд Unioniina. Униониины (лат. unio - объединение, а также одна жемчужина) объединяют пресноводных бивальвий, имеющих равностворчатую раковину до 30 см длиной, с хорошо развитым периостраком, опистодетным лигаментом, двумя псевдокардинальными и одним боковым зубами в левой створке (иногда полностью редуцированными). У некоторых представителей имеется псевдотаксодонтный замок. Не исключена возможность, что в подотряд объединены неродственные группы, возникшие от разных предков и в разное время приспособившиеся к жизни в пресных водах. У современных унионин в онтогенезе имеются личинки - глохидий, лазидий или гаусториальная. Подотряд появился в конце девона, дал довольно много представителей в карбоне и перми (надсемейство Anthracosiacea, около 20 родов) и получил широкое распространение начиная с триаса (надсемейство Unioniacea свыше 150 родов) (рис. 176, 4-5).
Подотряд Actinodontina. Актинодонтины объединяют морских, преимущественно палеозойских, бивальвий, раковина которых имеет актинодонтный замок, несколько напоминающий лиродесмоидный, на отличающийся от него развитием зубов, идущих параллельно спинному краю раковины и укороченных под макушкой. Актинодонтины появляются в среднем кембрии и представляют собой наиболее древних бивальвий, от которых возникли все остальные группы аутобранхий. Они объединяют около 40 родов; основные представители вымирают к концу перми, а их потомки, если включить в этот подотряд современный род Gastrochaena, существуют до наших дней (рис. 176, 6-8).
Отряд Pectinida. Пектиниды (лат. pecten - гребешок). Раковина округленная или вытянутоовальная, иногда неправильной формы, равно- и неравностворчатая, вдоль спинного края развиты ушки треугольной формы; преобладает-радиальная скульптура, ребра от слабых до очень сильных; концентрическая скульптура иногда сопровождается выростами, шипами; структура внутреннего слоя перекрещенно-пластинчатая или листоватая. Замок изодонтный или с кардинальными крурами; иногда отсутствует. Лигамент наружный и внутренний. Мантия: с несросшимися утолщенными краями, которые загнуты внутрь, образуя своеобразный "парус". Вдоль края мантии развиты фоторецепторы-глазки (иногда до 100), короткие щупальца и другие органы чувств. Имеется один большой мускул, занимающий почти центральное положение, разделенный на переднюю часть, состоящую из поперечных мышц, способную к быстрым сокращениям, и заднюю с гладкими мышцами, приспособленную для удержания створок в закрытом состоянии. Жабры состоят из свободных филаментов (нитей), образующих две ветви: нисходящую и после коленообразного изгиба - восходящую. Желудок пектиноидного типа, большой тифлозоль сравнительно короткий. Отверстия дивертикул многочисленные. Пектиниды-фильтраторы, обитают во всех морях и океанах, на самых разнообразных глубинах, от мелководья до ультраабиссали (до 8000 м). На ранних стадиях они прикрепляются с помощью биссуса; одни всю жизнь остаются прикрепленными к субстрату при помощи биссусного лучка, превращенного в особый известковый стебелек, выходящий через отверстие в нижней створке (Anomia), или прочно цементируются нижней створкой к подводным предметам (Spondylus); другие из биссусных нитей строят себе гнездо, скрепляя биссусом мелкие камешки, обломки раковин и другой строительный материал, выстилая "гнездо" мягкими биссусными нитями (Lima). Преобладающее же число свободно лежит на дне. и способно в случае опасности передвигаться скачками, хлопая створками (Pecten, Lima); в этом случае при захлопывании створок края "паруса" плотно смыкаются и вода с силой выталкивается через отверстия в основании ушек. К лучшим "пловцам" относятся формы с гладкой раковиной.
Пектиниды, объединяющие свыше 100 родов, появились в ордовике, но широкое развитие получили начиная с мезозоя (рис. 177). Отряд Astartida. Астартиды (гр. Astarte - финикийская богиня любви и плодородия). Раковина равностворчатая, иногда зияющая или окруженная со всех сторон мантией (становится внутренней с редукцией аддукторов); преобладает концентрическая скульптура, имеются луночка и щиток. Замок астартоидный и люциноидный, иногда полностью редуцированный; два аддуктора, лигамент преимущественно опистодетный. Структура внутреннего слоя перекрещенно-пластинчатая, реже гомогенная или призматическая. Жабры пластинчатые, состоят из филаментов, соединенных перемычками; мантия с несросшимися краями или срастается, оставляя отверстия для ноги и сзади для сифонов.
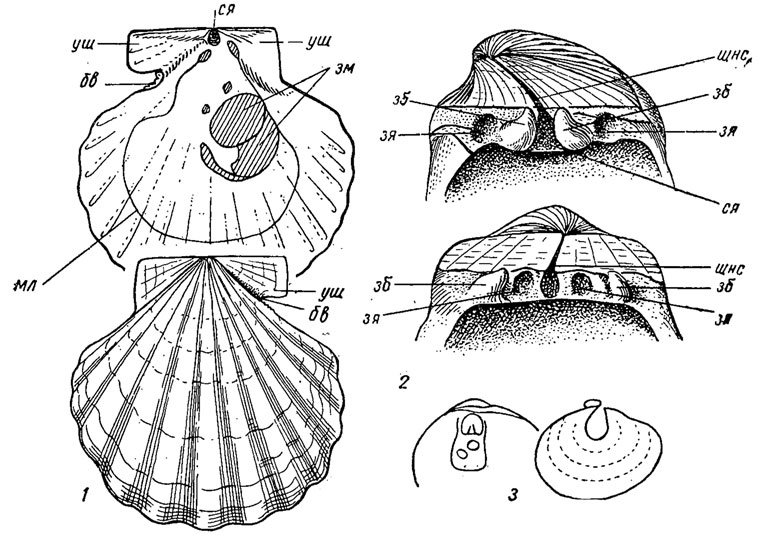
Рис. 177. Отряд Pectinida: 1 - Chlamys (триас - ныне), правая створка снаружи и изнутри; 2 - Spondylus (юра - ныне), схема строения замка правой (вверху) и левой (внизу) створок; 3 - Anomia (мел - ныне); oбв - биссусный вырез, зб - зубы, зм - задний мускул, зя - зубные ямки, мл - мантийная линия, ся - ямка для внутренней связки, уш - ушки, щнс - щель для наружной связки
Астартиды - обитатели морей, преимущественно умеренных и приполярных; живут в зоне мелководья, обычно на поверхности грунта; немногие зарывающиеся обладают вытянутыми сифонами и соответственно имеют мантийный синус. Первые астартиды появляются в ордовике, но многочисленными и разнообразными становятся начиная с мела. Они объединяют свыше 270 родов (рис. 178).
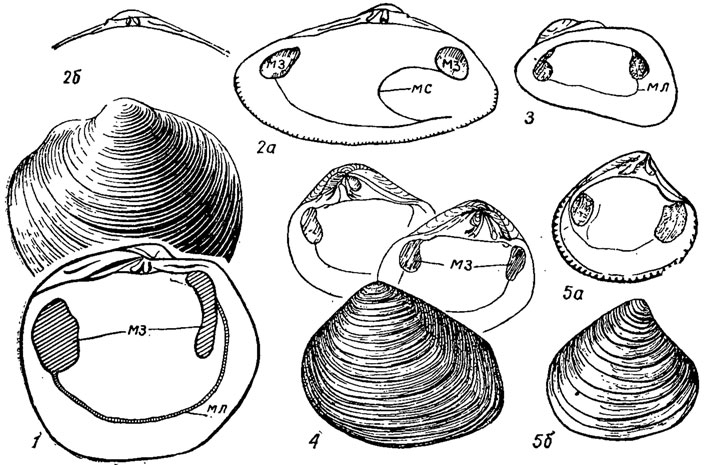
Рис. 178, Отряд Astartida: 1 - Lucina (поздний мел - ныне); 2 - Dotiax (эоцен - ныне); 3 - Hiatella (поздняя юра - ныне); 4 - Crassatella (поздний мел - неоген); 5 - Astarte (юра - ныне); мз - мускул - замыкатель, мл - мантийная линия, мс - мантийный синус
Отряд Carditida. Кардитиды (гр. cardia - сердце). Раковина равностворчатая, сердцевидная, тригональная, трапециевидная; замок кардитоидный, сходный с астартоидным по числу кардинальных зубов (2-3), но отличающийся от него сильной скошенностью зубов; имеется по одному латеральному зубу с каждой стороны. Преобладает радиальная скульптура; макушка прозогирная. Лигамент наружный, поддерживается нимфами, иногда внутренний; имеются два мускула-замыкателя; мантийная линия интегрипаллиатная. Жабры пластинчатые. Желудок кардитоидного типа, большой тифлозоль очень длинный, закрученный на конце в многооборотную спираль; дивертикулы печени открываются тремя - четырьмя группами отверстий. По типу питания-инфаунные фильтраторы. Первые кардитиды появляются в девоне, становятся более разнообразными начиная с мела; современные кардитиды живут преимущественно в тропических и субтропических морях, немногие встречены в мелководье умеренных морей. Известно около 80 родов.
Отряд Venerida. Венериды (лат. Venus - богиня любви). Один из наиболее крупных отрядов аутобранхий. Объединяет свыше 400 родов, из них большинство ранее относимых к отряду гетеродонт (без астартид и кардитид) и почти все роды, относимые к отряду десмодонт. Жабры пластинчатые, желудок венероидного типа, с одним или двумя слепыми впячиваниями; большой тифлозоль напоминает цифру 7. Раковина равностворчатая у большинства; у прирастающих (подотряд хамина) резко неравностворчатая; замок циреноидный, люциноидный или десмодонтный. Обитатели морей, солоноватоводных и пресноводных бассейнов, по типу питания - инфаунные фильтраторы или собиратели-грунтоеды; среди них: живущие на поверхности осадка, частично и полностью зарывающиеся, сверлильщики известковых пород (фоладины) и древесины (тередины), прикрепленные биссусом. Венериды, вероятно, возникли в позднем триасе от астартид. Основной ствол, выделенный в подотряд венерина, - обитатели дна моря, ползающие, частично погруженные в осадок или зарывающиеся, с хорошо развитыми сифонами; в юре от венерин обособилась группа сверлильщиков известковых пород, плотных илов и глин, а начиная с палеоцена или конца мела - группа сверлильщиков древесины, выделенная в подотряд фоладина. В палеоцене часть венерид перешла к прикрепленному образу жизни, цементируясь одной из створок, и в связи с этим претерпела резкие изменения внешней формы раковины (подотряд хамина).
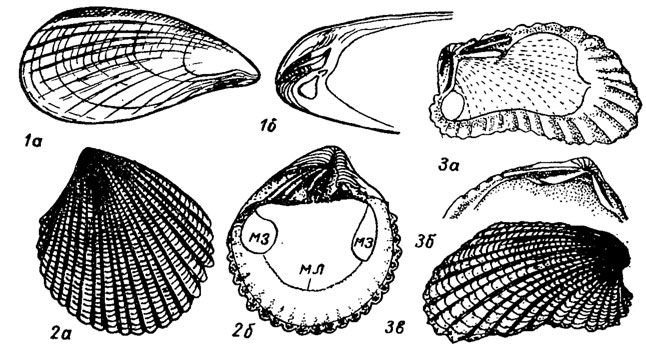
Рис. 179. Отряд Carditida: 1 - Myoconcha (юра - мел); 2 - Vertericardia (палеоцен - ныне); 3 - Cardita (палеоцен - ныне); мз - мускул-замыкатель, мл - мантийная линия
Подотряд Venerina. Венерины объединяют равностворчатых бивальвии с люциноидным и циреноидным замками или без зубов, с концентрической и радиальной скульптурой, от слабо выраженной до очень сильной; имеют раковины от маленьких до гигантских размеров, достигающих длины 1,4 м и веса до 200 кг (Tridacna). Структура внутреннего слоя перекрещенно-пластинчатая или гомогенная. Имеют два мускула-аддуктора, паллиальную линию с синусом и без синуса; среди зарывающихся форм обособились две группы: с длинной, почти цилиндрической раковиной и короткими сифонами (сем. Solenidae) и с овальной, зияющей на конце раковиной и очень длинными сифонами (сем. Myidae). Обитатели морей, солоноватоводных и пресноводных бассейнов. К жизни в пресных водах перешли дрейссенсиды, прикрепляющиеся биссусом, с двумя неравными мускулами, внешне напоминающие митилид из отряда циртодонтид. Венерины известны с конца триаса и существуют до наших дней. Объединяют около 300 родов (рис. 180).
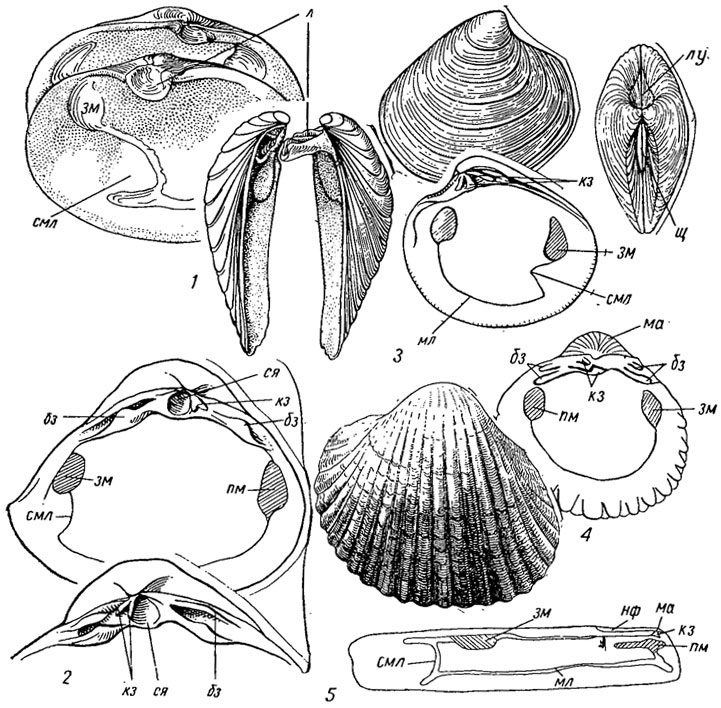
Рис. 180. Отряд Venerida; подотряд Venerina: 1 - Mya (олигоцен - ныне); 2 - Mactra (эоцен - ныне); 3 - Venus (олигоцен - ныне); 4 - Cardium (неоген - ныне); 5 - Solen (миоцен - ныне); бз - боковые зубы, зм - задний мускул, кз - кардинальный зуб, л - ложечка, лу - лунка, ма - макушка, мл - мантийная линия, нф - нимфа, пм - передний мускул, смл - синус мантийной линии, ся - ямка для внутренней связки, щ - щиток
Подотряд Chamina. Хамины (гр. chamai - живущие на грунте) объединяют три рода, представители которых прикрепляются к подводным камням путем цементации; створки неравные, неправильных очертаний, с прозогирной макушкой, концентрической или радиальной скульптурой с шипами; замок редуцирован; сохранилось по одному кардинальному зубу в каждой створке и у некоторых по два кардинальных и одному латеральному; по внешней форме зубы несколько напоминают зубы гиппуритид. Хамины появились в конце мела (датском ярусе) и в настоящее время являются обитателями теплых морей (рис. 181, 1).
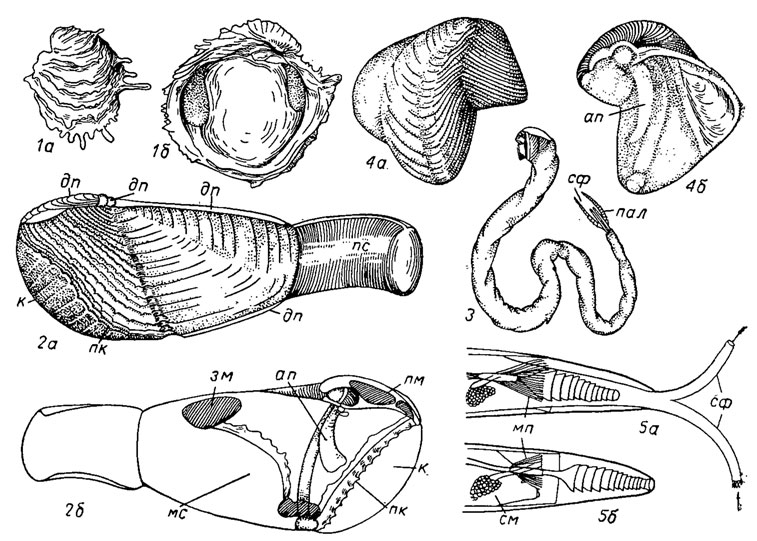
Рис. 181. Отряд Venerida; подотряды Chamina (1) и Pholadina (2-5): 1а, б - Chama (палеоцен - ныне); 2 - схема строения раковины сем. Pholadidae: 2а - вид снаружи, 2б - изнутри; 3-4 - сем. Teredinidae, Bankia: 3 - общий вид, 4а, б - раковина, увел.; 5 - схема взаимоотношения сифона и палетки: 5а - сифон выставлен, 5б - сифон втянут; ап - апофиз, дп - дополнительные пластинки, зм - задний мускул, к - каллум, мп - мускулы палеток, мс - мантийный синус, пал - палетки, пк - передний край раковины, пм - передний мускул, пс - пластинки, покрывающие сифон, см - сифональный мускул, сф - сифон
Подотряд Pholadina. Фоладины (гр. pholas - моллюски, сверлящие породы) - труппа, приспособившаяся к сверлению известковых пород или древесины. В соответствии с подобной адаптацией выделяют морских сверлильщиков - фоладид и древоточцев - терединид; первые объединяют 28 родов, вторые - 21. Сверлящий образ: жизни привел к существенным изменениям раковины и отдельных ее структур.
Раковина у фоладид равностворчатая, без лигамента и замка; передний край образует макушечную складку - отворот, к которому прикрепляется передний аддуктор; задний аддуктор крупный; существует еще один дополнительный аддуктор на брюшной стороне; мускулы передний и задний действуют поочередно. Передняя часть раковины вооружена резкими шиповатыми ребрами, играющими основную роль при сверлении. Каждая створка имеет внутри раковины ножевидный апофиз, служащий для прикрепления мускулов ноги. Сифоны сильные, сросшиеся, покрытые хитинизированным периостраком; нога с подошвой служит для прикрепления к стенкам норки. Фоладиды сверлят известковые породы, сжимая поочередно передний и задний аддукторы и частично поворачивая раковину, проделывают ходы в плотном иле, глине; один род (Xylophaga), сверлящий древесину, недавно был встречен на глубине около 7 тыс. м. Раковины фоладид снабжены дополнительными пластинками, покрывающими раковину на спинной стороне спереди и сзади и с брюшной стороны. У некоторых форм (мартезий) передний зияющий край бывает закрыт полностью или частично особой известковой оболочкой - каллумом (рис. 181, 26).
Терединиды, приспособленные к сверлению древесины, имеют очень маленькую раковину, зияющую спереди и сзади, с зазубренной передней частью; тело длинное, червеобразное, с двумя сифонами на заднем конце, расположено вне раковины; у основания сифонов имеется пара известковых пластинок - палеток, которые при втягивании сифонов внутрь норки закрывают входное отверстие; у одних палетки простые, ложкообразные, у других напоминают колосок, состоят из мелких, входящих друг в друга воронковидных зазубренных конусов (рис. 181, 5). Сверление древесины, как и у фоладид, происходит с помощью переднего зазубренного края раковины при поочередном сокращении то переднего, то заднего мускулов и легкого поворачивания тела при помощи ноги, играющей роль присоски. Ходы древоточцев изнутри выстланы тонким слоем известковых отложений. Тередины питаются мелкими планктонными организмами и дополнительно древесными опилками, которые перевариваются внутриклеточно в дивертикулах печени. В отличие от фоладид они гермафродиты.
В ископаемом состоянии представители подотряда встречаются начиная с палеоцена. От фоладин кроме раковин сохраняются просверленные ходы, дополнительные пластинки, палетки.
Отряд Hippuritida. Гиппуритиды (названы по роду Hippurites, имеющему сходство с рогом) объединяют свыше 130 родов крупных двустворчатых моллюсков, впервые появившихся в позднем силуре (надсем. Megalodontacea). Палеозойские гиппуритиды имели равностворчатую гладкую раковину, с прозогирной макушкой, толстыми кардинальными зубами (от одного до нескольких), с наружным, опистодетным лигаментом и с глубоко сидящим передним аддуктором, расположенным в передней части дорзального края. В начале поздней юры (оксфорде) от мегалодонтаций возникли рудисты.
У рудистов две створки неравные; различают три основных варианта: 1) прикрепленная створка - спирально закрученная, свободная - в форме крышечки; 2) прикрепленная створка длинная, большая, коническая, свободная - плоская, коническая или закрученная; 3) прикрепленная - маленькая, коническая, свободная - значительно крупнее первой, закрученная. Прикрепление осуществлялось обычно левой или инверсно-правой створкой. У позднеюрских и раннемеловых форм створки отличались закрученностью (Diceras), у позднемеловых преобладали формы с конической раковиной, внешне напоминающей скелеты одиночных кораллов (Radiolites, Hippurites). Конические раковины некоторых рудистов достигали 2 м в высоту и до 0,6 м в диаметре. Обычно имелись два крупных зуба в крышке и один в прикрепленной створке, или наоборот. Лигамент наружный у ранних форм, внутренний - у поздних и, возможно, у некоторых отсутствовал.
У рудистов стенка обычно толстая, до 10 см толщиной, ячеистопризматическая, с толстым призматическим наружным слоем, состоящим из полых призм; у ранних рудистов стенка ячеистая. В стенке наблюдаются различные каналы, вертикальные и радиальные, простые и разветвленные. У ряда форм отмечаются поперечные днища, отделявшие тело от нижней нежилой части раковины. Кроме основной внутри раковины имелись еще дополнительные полости. Створки соединялись двумя мускулами-аддукторами, которые прикреплялись к особым выступам - миофорам. Наличие мускулов к лигамента позволяем предполагать, что створки у основных групп рудистов открывались и закрывались, так же как у всех ординарных бивальвий. Однако у радиолитов и гиппуритов с плоской крышкой и вертикально стоящими зубами и соответствующими зубными ямками верхняя свободная створка могла, скорее всего, двигаться только вертикально. Возможно, движение осуществлялось вследствие изменения объема ноги и тела. Для обеспечения питанием и газообмена существовало два сифона, структуры которых выражены углублениями, складками в наружной стенке, особыми столбиками; один сифон был вводным, другой - выводным (рис. 182). Первые рудисты, как уже было отмечено, появились в начале поздней юры, ответвившись от протодицераса (надсем. Megalodontacea). Рудисты были обитателями тропических морей, главным образом коралловых рифов, жившими большими поселениями; в раннемеловое время они были сравнительно немногочисленны (около 25 родов), но стали очень разнообразными и многочисленными в позднемеловое время (до 80 родов) (рис. 182). Однако в конце Маастрихта все рудисты вымерли. Невесская и др. (1971) присоединяет к гиппуритидам одно современное семейство Arcinellidae, которое большинство малакологов относит к хаминам.
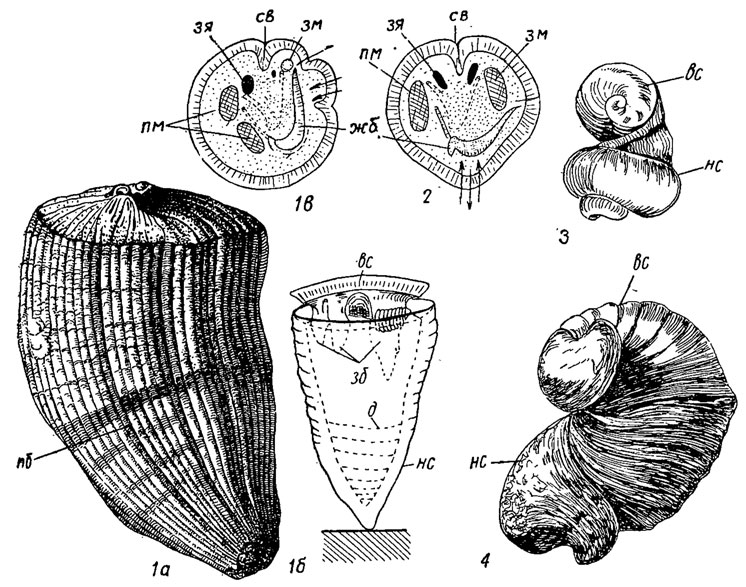
Рис. 182. Отряд Hippuritida: 1 - Hippurites (поздний мел): 1а - внешний вид, 1б - продольный разрез, 1в - поперечный разрез; 2 - Radiolites (поздний мел), поперечный разрез; 3 - Requienia (мел); 4 - Diceras (поздняя юра); вс - верхняя створка, д - днища, жб - жабры, зб - зубы, зм - задний мускул, зя - зубные ямки, не - нижняя створка, пб - продольные борозды, пм - передний мускул, св - связка; стрелки указывают направление тока воды
Отряд Cyrtodontida. Циртодонтиды (гр. kyrtos - изогнутый) имеют нитчатые жабры со свободными филаментами, желудок митилоидного типа; большой тифлозоль S-образно изогнут, дивертикулы печени открываются рядом отверстий. Отряд возник от актинодонтид в начале ордовика и сразу же дал ветви, выделенные в подотряды Arcina и Pteriina, отличающиеся строением замка и раковины. В девоне возник подотряд устриц, а позднее, в триасе, ответвился подотряд митилин.
Подотряд Arcina. Арцины (лат. arca - коробка). Раковины овальные, трапецеидальные; замок циртодонтный и псевдоктенодонтный, состоящий из ряда зубов, расположенных в один ряд вдоль замочного края. Под макушкой развита арея, на которой расположена сложная связка в шевронах. Нога развита слабо; имеются биссусная железа, два мускула-аддуктора; появляются в ордовике и в настоящее время живут преимущественно в тропических морях. Объединяют до 80 родов (рис. 183, 1,2).
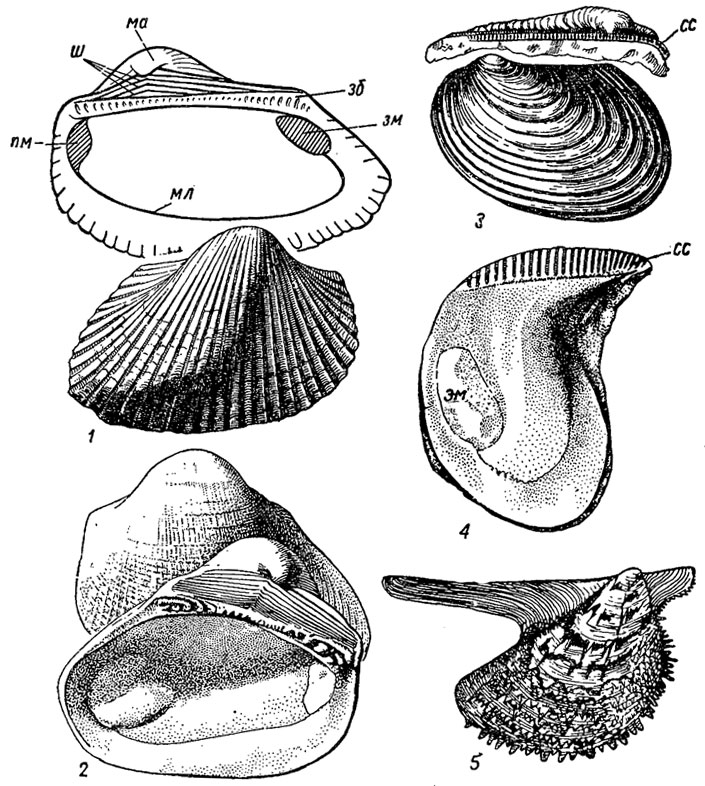
Рис. 183. Отряд Cyrtodontida; подотряды Arcina (1-2) и Pteriina (3-5): 1 - Area (средняя юра - ныне); 2 - Cucullaea (юра - ныне); 3 - Inoceramus (юра - мел); 4 - Isognomon (трас - ныне); 5 - Pteria (триас - ныне); зб - зубы, зм - задний мускул, ма - макушка, мл - мантийная линия, пм - передний мускул, сс - сложная связка, ш - шевроны
Подотряд Pteriina. Птериины (гр. pteron - крыло). Раковины равно- и неравностворчатые, с крыловидными ушками, тригональные квадратные или трапецеидальные; нога с биссусом; спинной край прямой и несет многочисленные ямки для сложной связки. Скульптура концентрическая и радиальная. Замок птериноидный, с задними краевыми зубами неравной длины или лишенный зубов. Птериины появляются в ордовике, в карбоне приспосабливаются к жизни в солоноватоводных бассейнах. В юрское и меловое время получает широкое распространение семейство иноцерамид, имевших разнообразной формы раковину, сложную связку и хорошо развитый призматический слой. Современные птериины - обитатели мелководья, преимущественно теплых морей. Объединяют до 150 родов (рис. 183, 3-5).
Подотряд Mytilina. Митилины (гр. mitos - нить). Раковины треугольной формы, суженные спереди и расширенные сзади, образуют значительные поселения в прибрежной части моря; мускулы неравные: передний - маленький, задний - большой; нога пальцевидная, у молодых служит для переползания, у взрослых выделяет биссусные нити: для прочного прикрепления к подводным скалам или к песчинкам при частичном погружении в осадок; связка опистодетная; хороша развиты сифоны; замок без зубов или со слабо развитыми зубчиками в примакушечной части. Первые митилины появляются в девоне, в конце перми часть из них приспосабливается к сверлению известковых пород, однако сверление в отличие от фоладид осуществляется кислотой, выделяемой особой железой и растворяющей известковое вещество (лигофаги, или морские финики); поэтому раковина у них тонкая, более или менее удлиненная, почти цилиндрическая. Подотряд объединяет около 60 родов (рис. 184).
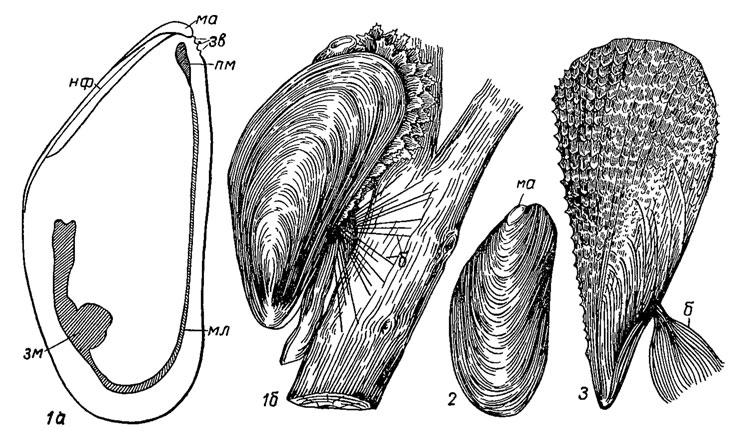
Рис. 184. Отряд Cyrtodontida; подотряд Mytilina: 1 - Mytilus (поздняя юра - ныне): 1а - левая створка изнутри, 1б - прижизненное положение; 2 - Modiolus (девон - ныне); 3 - Pinna (ранний карбон - ныне); б - биссус, зв - зубовидные выступы, зм - задний мускул, ма - макушка, мл - мантийная линия, нф - нимфа, пм - передний мускул
Подотряд Ostreina. Остреины, или устрицы (лат. ostrea, гр. ostreon - устрица); ведут прирастающий образ жизни, имеют неравностворчатую раковину: левая створка блюдцевидная, правая - плоская, в форме крышечки. Макушка прирастающей створки может быть клювовидной, нависающей (род Gryphaea) или, завернута в форме плоской спирали (Exogyra). После цементации нога и биссус атрофируются, структура внутреннего слоя листоватая. Имеют открытую мантию и свободные нитчатые жабры. Замок без зубов или с рудиментами таксодонтных зубчиков. Лигамент внутри ямки на примакушечной арее. Обитатели мелководья, живут на плотных грунтах и скалах на глубине до 50-70 м, образуя устричные банки. Могут выносить значительное опреснение. Появляются в триасе и существуют до наших дней преимущественно в теплых морях. Объединяют свыше 50 родов (рис. 185).
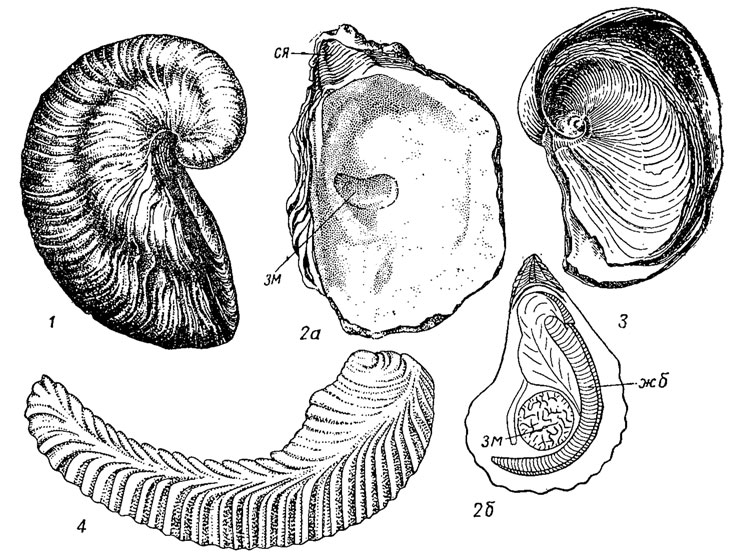
Рис. 185. Отряд Cyrtodontida; подотряд Ostreina: 1 - Gryphaea (юра); 2 - Ostrea (мел - ныне): 2а - левая створка изнутри, 2б - схема внутреннего строения; 3 - Exogyra (мел); 4 - Lopha (триас - ныне); жб - жабры, зм - задний мускул, ся - ямка для связки
Отряд Pholadomyida. Фоладомииды. Раковины равностворчатые с двумя мускулами, опистодетным лигаментом, у некоторых (Pandora) лигамент с литодесмой. Скульптура концентрическая и радиальная; замок десмодонтный, внутренний слой перламутровый; жабры пластинчатые, желудок пандороидного типа - большой тифлозоль сравнительно короткий; ведут зарывающийся образ жизни; хорошо развиты сифоны. Фоладомииды возникли в девоне, по-видимому, от отряда актинодонтид. В связи с зарывающимся образом жизни у одной группы фоладомиид (Clavagella) раковины полностью или частично редуцировались и тело заключено в известковую трубку (обособлены в подотряд клавагеллин). Объединяет около 90 родов (рис. 186).
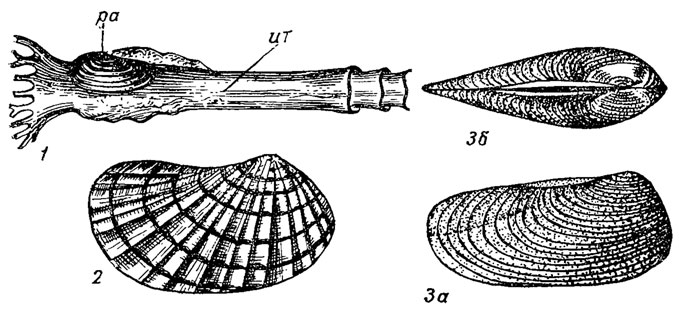
Рис. 186. Отряд Pholadomyida: 1 - Clavagella (поздний мел - ныне); 2 - Pholadomya (поздний триас - ныне); 3 - Allorisma (девон - пермь): 3а - вид сбоку, 3б - вид сверху; ит - известковая трубка, ра - раковина
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'