
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Класс Crinoidea. Криноидеи, или морские лилии
Общая характеристика. Криноидеи (гр. krinon - лилия), или морские лилии, - наиболее крупный класс кринозой, тело которых состоит из чашечки, заключающей внутренние органы, пяти обычно хорошо развитых рук, служащих для сбора пищи, и стебля или системы усиков, предназначенных для прикрепления к подводным предметам. Чашечка радиально-симметричная, построена из пояска радиальных табличек и одного или двух поясков основных табличек. Чашечка сверху прикрыта крышечкой, или тегменом, в которой находятся амбулакральные желобки, переходящие на руки и далее на пиннулы. Ордовик - ныне.
Строение тела. Внутренние органы морской лилии заключены в чашечку, в центре которой на верхней стороне расположено ротовое отверстие (рис. 263). Рот ведет в пищеварительный тракт, делающий один или несколько петлеобразных изгибов и открывающийся анальным отверстием в заднем интеррадиусе. Пищеварительный тракт лежит во вторичной полости тела и подвешен к стенкам тела при помощи мезентериальных перепонок. Ют чашечки отходят пять неразветвленных или ветвящихся рук. Чашечка вместе с руками образует крону. Вокруг пищеварительного тракта расположен кольцевой канал амбулакральной системы; от него в руки отходят, пять радиальных каналов, вдоль которых расположены амбулакральные ножки; у морских лилий они остроконечны, лишены ампул, присасывательных дисков и выполняют пищесборную, дыхательную и чувствительную функции. Пищей криноидеям служат планктонные организмы и мелкие частицы детрита. Пища доставляется ко рту по желобкам на руках с помощью амбулакральных ножек и ресничек покровного эпителия. Количество пищи, добываемой морской лилией, зависит от степени разветвления рук и соответственно длины бороздок, или желобков. У одной тропической лилии с 68 ветвями рук общая длина пищевых борозд достигает 100 м. Вокруг рта расположено нервное кольцо, от которого по пяти радиусам в руки отходят нервные стволы, обеспечивающие их движение.
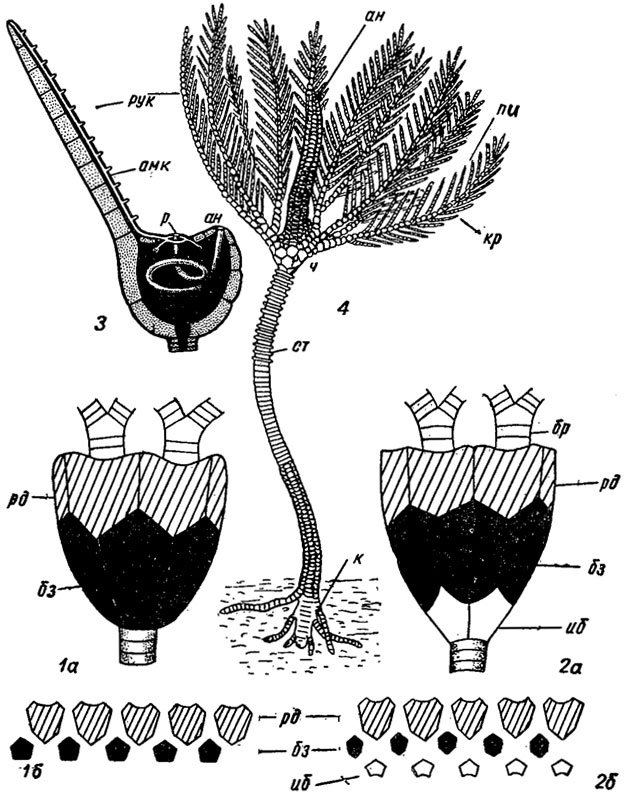
Рис. 263. Схема строения морской лилии: 1а, б - моноциклическая чашечка; 2а, б - дициклическая чашечка; 3 - схематический разрез через чашечку; 4 - общий вид прикрепленной морской лилии; амк - амбулакральный канал, ан - анальное отверстие, к - 'корни', кр - крона, пи - пиннулы, р - рот, рук - руки, ст - стебель, ч - чашечка, таблички: бз - базальные, бр - брахиальные, иб - инфрабазальные, рд - радиальные
Скелет чашечки. Чашечка, или тека, разнообразной формы, коническая, бокаловидная, дисковидная или шаровидная (рис. 263). Часть чашечки ниже мест прикрепления рук называется спинной, или дорзальной, а верхняя часть - крышечкой, или тегменом. Дорзальная часть теки образована двумя или тремя поясками табличек. Различают пояски: радиальных (RR), базальных (BB) и инфрабазальных (IB) табличек; каждый из поясков состоит из пяти табличек. От основания чашечки отходит стебель или у бесстебельчатых форм - усики, или цирри; к радиальным табличкам прикрепляются руки. Чашечка, дорзальная часть которой кроме пояска радиальных табличек имеет поясок базальных, называется моноциклической; если она имеет поясок базальных и инфрабазальных табличек - дициклической. Дорзальная часть теки иногда бывает построена исключительно из радиальных табличек, реже только из базальных. Часто в строении дорзальной части принимает участие ряд других табличек, среди которых выделяются анальная (одна или несколько), расположенная в заднем интеррадиусе, радианальная и др. В процессе эволюции у морских лилий наблюдается увеличение размеров чашечки. Это увеличение происходит за счет включения в состав чашечки члеников нижних частей рук и развития новых, так называемых интеррадиальных и интербрахиальных табличек (см. рис. 271, 5-8).
Скелет руки. От радиальных табличек чашечки отходят руки. Они редко остаются простыми, большей частью раздваиваются один или несколько раз. Руки состоят из отдельных члеников, напоминающих по форме позвонки, которые соединяются друг с другом с помощью мышц или эластичной связки. Как правило, они снабжены короткими членистыми придатками - пиннулами. Членики рук сочленяются также при помощи особых площадок, часто имеющих один или два валика. Руки обладают гибкостью и большой подвижностью. При неблагоприятных условиях (высокой температуре, недостатке кислорода, нападении врагов) морские лилии способны обламывать свои руки, при этом утраченные части позднее восстанавливаются. Руки и пиннулы снабжены довольно глубокими желобками, выстланными у современных форм ресничным эпителием. Вдоль желобка проходит радиальный амбулакральный канал, от которого отходят пучками (по 3) остроконечные амбулакральные ножки без ампул; они выполняют функцию осязания и дыхания. Боковые ветви радиальных каналов заходят также в пиннулы.
Руки предназначены для сбора пищи. В руки продолжаются вторичная полость тела, нервные стволы, сосуды кровеносной системы. По пищевым желобкам рук пища попадает в ротовое отверстие, расположенное в центре тегмена. В процессе эволюции увеличивается длина и степень разветвленности рук. Однорядная рука у примитивных форм заменяется двухрядной (рис. 264, 2); двухрядная рука позволяет морской лилии собирать больше пищи. Увеличение длины рук происходит при их дихотомическом ветвлении или при образовании перистой руки (рис. 264, 1). Однако в процессе эволюции возникали морские лилии, у которых руки частично или полностью редуцировались. При редукции рук могли исчезать и поддерживающие их радиальные таблички чашечки.
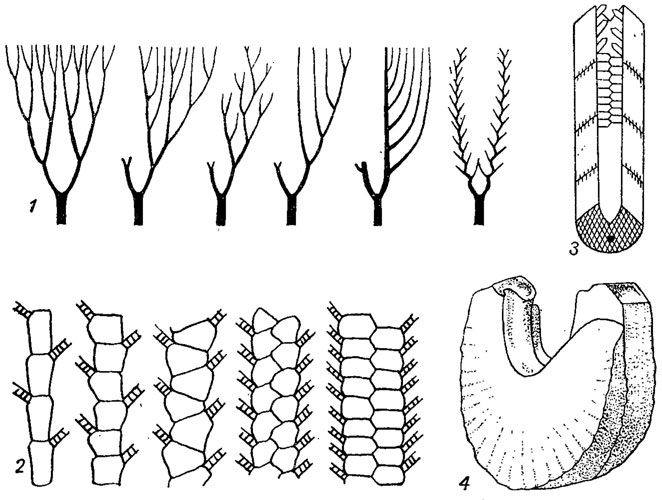
Рис. 264. Схема строения рук: 1 - развитие перистой руки из дихотомически разветвленной; 2 - развитие двухрядной руки из однорядной; 3 - часть руки (4 членика) с амбулакральным каналом и кроющими табличками; 4 - два членика руки Diamenocrinus (девон) рук могли исчезать и, поддерживающие их радиальные таблички чашечки
Тегмен у большинства современных форм почти совершенно лишен крупных скелетных элементов. Он пронизан многочисленными порами, ведущими в полость тела; через , поры амбулакральная система пополняется водой. Амбулакральные ножки, расположенные около рта, бывают видоизменены в околоротовые щупальца. У древних морских лилий тегмен был прикрыт пятью оральными, или ротовыми, табличками, расположенными интеррадиально (рис. 265). Оральные таблички развиты в разной степени: у одних форм они известны только в личиночной стадии и отсутствуют у взрослых; у других - хорошо развиты и плотно соединяются друг с другом; у третьих крышка состоит из многочисленных мелких табличек, среди которых различают таблички, покрывающие пищевые желобки, и расположенные между ними интерамбулакральные таблички. Эти таблички, соединяясь друг с другом, образуют своеобразный свод над чашечкой; под таким сводом расположен рот, и пища поступает по пищевым желобкам, лежащим под крышкой.
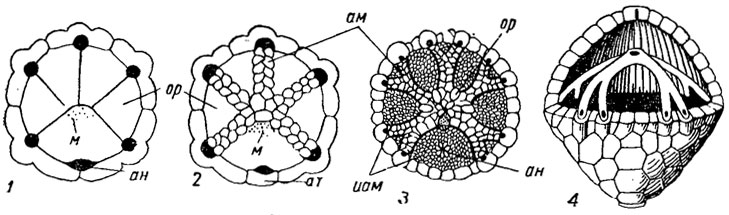
Рис. 265. Типы строения крышки (тегмена): 1 - развиты только оральные таблички; 2 - крышка с амбулакральными полями; 3 - оральные таблички редуцированы (сохранились около рта); 4 - пищевые желобки расположены под прочной крышкой; ан - анальное отверстие, м - мадрепорит, таблички: ам - амбулакральные, ат - анальная, иам - интерамбулакральные, ор - оральные
Анальное отверстие размещается на верхней стороне орального диска чашечки интеррадиально, ближе к ее краю. У морских лилий, живших в спокойных, малоподвижных водах, возникала анальная трубка, покрытая мелкими табличками. Подобная трубка позволяла животному удалять экскременты на значительном расстоянии от рта.
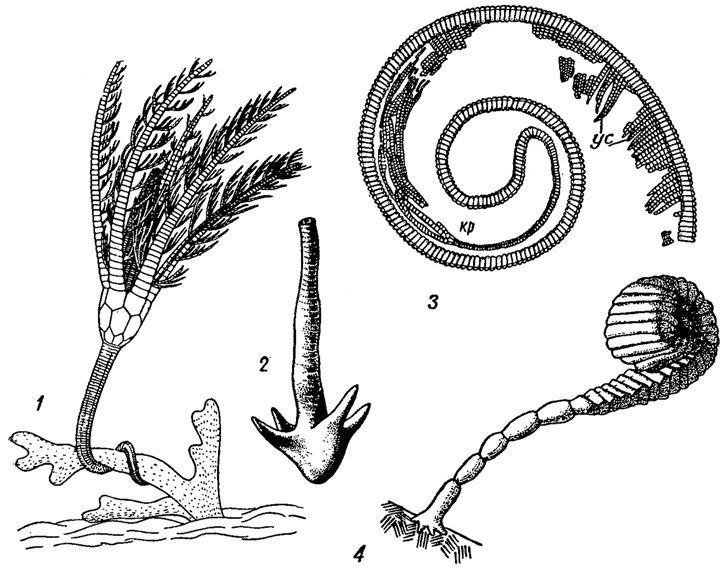
Рис. 266. Типы стеблей морских лилий: 1 - стебель Eifelocrinus, прикрепленный к колонии мшанок (реконструкция); 2 - 'якорь' у Aticyrocrinus; 3 - двусторонне-симметричный стебель с усиками (ус) у Myelodactylus, окружающий крону (кр); 4 - стебель, спирально свернутый вокруг чашечки у Ammonicrinus
Стебель. К нижней стороне чашечки, к ее центральной табличке, прикреплен гибкий стебель, состоящий из члеников различной формы: круглой, эллиптической, четырехугольной, пятиугольной и очень редко треугольной и шестиугольной (рис. 266). У одних родов стебель достигает длины нескольких метров, у других остается коротким или совершенно атрофируется. У некоторых форм чашечка прирастала своим основанием. Через весь стебель проходит осевой канал, имеющий различное поперечное сечение. У древних морских лилий стебель состоял из пяти рядов табличек, расположенных в чередующемся порядке. В процессе эволюции наблюдается переход к циклическому расположению и к слиянию при этом каждых пяти соседних табличек в один членик стебля (рис. 267). Нередко среди одинаковых члеников встречаются более крупные так называемые узловые членики, несущие усики. Морские лилии прикрепляются к субстрату разными путями: прирастанием стебля к каменистому дну путем выделения вокруг основных члеников значительного количества извести и образования прикрепительного диска, развитием на конце стебля разветвленных корневидных ветвей, наличием своеобразного якоря, предназначенного для закрепления У некоторых морских лилий длинный тонкий стебель обвивал водоросли или полипняки кораллов и служил для временного прикрепления, у других - закручивался вокруг чашечки в плоскую спираль и, возможно, служил для медленного и недалекого передвижения по дну с помощью двурядно расположенных усиков (см. рис. 266, 5). Известно также развитие на нижнем конце стебля шаровидного вздутия, разделенного перегородками на отдельные камеры и служившего, по-видимому, плавательным пузырем при планктонном образе жизни. Наконец, у ряда форм стебель отсутствовал и отсутствует на взрослой стадии у многих современных криноидей. У таких бесстебельчатых морских лилий стебель существует только на первых стадиях развития в течение месяца-полутора, после чего их чашечка самопроизвольно отламывается от стебля и молодая морская лилия переходит к свободному образу жизни. В основании чашечки развиваются усики, или цирри. Движение таких лилий происходит при помощи рук, однако за один прием они проплывают небольшое расстояние (до 3 м), совершая при этом до 100 ударов в минуту. Число, размеры, длина и внешний вид усиков зависят от условий обитания: морские лилии, живущие на мягких илах, имеют тонкие длинные, почти прямые усики; лилии, обитающие на камнях, снабжены короткими изогнутыми усиками.
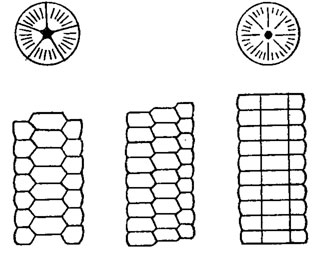
Рис. 267. Эволюция стебля от пятирядного к циклическому
Размножение и развитие. Наиболее подробно изучено размножение и развитие современной бесстебельчатой морской лилии, принадлежащей к роду Antedon (рис. 268). Морские лилии раздельнополые. Половые клетки созревают в пиннулах рук; выброс половых продуктов происходит обычно одновременно, и оплодотворение яиц осуществляется в воде. Оплодотворенные яйца заключены в оболочку, часто снабженную различными шипами, иглами. В этих оболочках происходит развитие яиц до стадии личинки. Вначале личинка лишена рта и питается только желтком. На брюшной стороне у нее имеется прикрепительная присоска. Поплавав некоторое время в воде, личинка опускается на дно, прикрепляется к субстрату передней частью тела. Узкая передняя часть превращается в стебель, а широкая задняя - в чашечку. Реснички, покрывающие тело личинки, исчезают, происходит вращение комплекса внутренних органов на 90°. Возникают пять оральных табличек, образующих на верхней стороне пирамиду, снизу развиваются пять базальных табличек. Между ними и началом стебля появляются 3-5 инфрабазальных табличек. В это время скелет молодой морской лилии несколько напоминает скелет некоторых палеозойских цистоидей. Вскоре между базальными и оральными табличками развивается поясок, состоящий из пяти радиальных табличек, и возникают руки. На границе между чашечкой и стеблем формируются новые членики стебля. Через пять недель после оседания личинки на стебельке раскачивается миниатюрная морская лилия высотой около 4 мм. В дальнейшем руки постепенно удлиняются, каждая рука делится на две ветви; вдоль руки возникают пиннулы, которые чередуются друг с другом. На этой стадии морская лилия похожа на представителей стебельчатой морской лилии рода Pentacrinus. Спустя некоторое время оральные таблички редуцируются, и на верхней стороне развивается кожный покров - тегмен. Базальные таблички также редуцируются. Затем чашечка самопроизвольно отламывается от стебля, и молодая лилия, превратившись в бесстебельчатую, начинает вести подвижный образ жизни, передвигаясь с помощью рук. Для временного прикрепления в основании чашечки развиваются цирри. Изучение онтогенеза современных морских лилий указывает на возникновение бесстебельчатых представителей от прикрепленных.
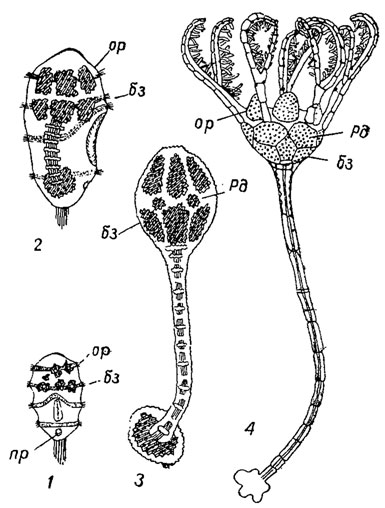
Рис. 268. Онтогенез Antedon bipida: 1-2 - свободноплавающие личинки (теменная пластинка обращена вниз); 3 - прикрепленная стадия (напоминающая цистоидей); 4 - пентакринусовая стадия с обособленными руками; пр - присоска, таблички: бз - базальные, ор - оральные, рд - радиальные
Основы систематики и классификация. Систематика морских лилий основывается на строении чашечки в целом, на строении ее дорзальной части, крышечки (тегмена), рук и стебля, на количестве и характере расположения анальных, интеррадиальных и интербрахиальных табличек. В состав класса включены четыре подкласса: Camerata, Inadunata, Flexibilia, Articulata, из которых первые три существовали от ордовика до перми, а представители четвертого, появившись в начале триаса, продолжают свое существование в современных морях (рис. 269-272).
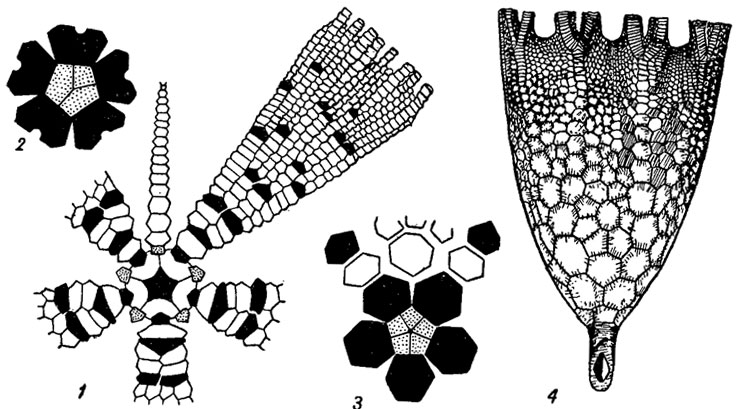
Рис. 269. Подкласс Camerata. Схема строения (1-3 - в плане, 4 - сбоку): 1 - Cleiocrinus (средний ордовик); 2 - Glyptocrinus (поздний ордовик); 3 - Platycrinites (девон - пермь); 4 - Acrocrinus (карбон)
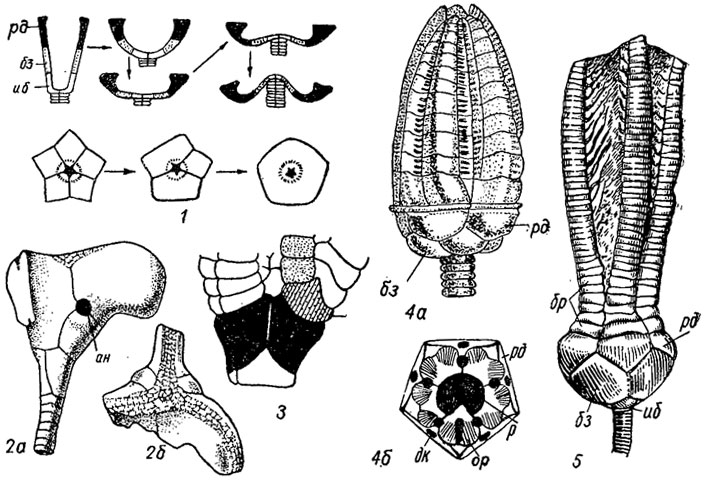
Рис. 270. Подкласс Inadunata. Схема строения: 1 - эволюция чашечки у инадунатных криноидей; 2 - Cornucrinus (ордовик): 2а - вид сзади, 2б - вид сверху; 3 - locrinus (средний - поздний ордовик); 4 - Cupressocrinites (средний девон): 4а - чашечка с руками, 4б - вид чашечки сверху; 5 - Cromyocrinus (карбон); дк - дорзальный канал, таблички: ан - анальная, ор - оральная (остальные обозначения см. рис. 263)
Экология и тафономия. Морские лилии в палеозое и мезозое были обитателями преимущественно мелководных зон моря. Часть из них жила под защитой коралловых рифов, часть опускалась на небольшие глубины. Современные морские лилии живут на всех глубинах: от сублиторали до абиссали (до 9700 м), от тропиков до полярных широт. Иногда они образуют большие поселения - "луга", состоящие из одного вида. Вероятно, такие "луга" морских лилий существовали в прошлые геологические времена, так как известны слои криноидных известняков, состоящих из члеников стеблей, реже рук и остатков чашечек. Глубоководные морские лилии имеют длинные ветвистые руки, тонкие стебли и небольших размеров чашечку; конце стебля развиты различной длины корешки. У морских лилий, живущих в теплых водах, чашечки более массивные, иногда снабженные шипами; стебель, как правило, укороченный, в некоторых случаях полностью отсутствует, и морская, лилия прикрепляется к субстрату непосредственно чашечкой. Большинство современных морских лилий ведет свободный образ жизни медленно плавая около дна при помощи рук. Подобные свободноплавающие криноидеи встречались уже в палеозое (силуре, перми), небольшая часть морских лилий, по-видимому, вела планктонный образ жизни. К ним относится силурийская форма (Scyphocrinites), у которой на конце стебля возник шаровидный плавательный аппарат (пневматофор). Меловая бесстебельчатая морская лилия (Saccocoma) плавала благодаря наличию маленькой чашечки и длинных рук. Малая подвижность морских лилий делает и делала их раньше доступными для поселения на них других организмов, для различного рода сожительства,, вплоть до настоящего паразитизма. Черви семейства Myzostomidae часто поселяются на морских лилиях, одни из них ползают по стеблю и чашечке, другие образуют на пиннулах для своего жилья особые вздутия, третьи паразитируют внутри тела. Некоторые брюхоногие моллюски, вероятно, были комменсалистами морских лилий. На брюшной стороне чашечки некоторых палеозойских криноидей находят раковины брюхоногих моллюсков (платинодератид), расположенные таким образом, что устьевой край был приближен к анальному отверстию морской лилии; не исключена возможность, что эти брюхоногие моллюски питались экскрементами лилии.
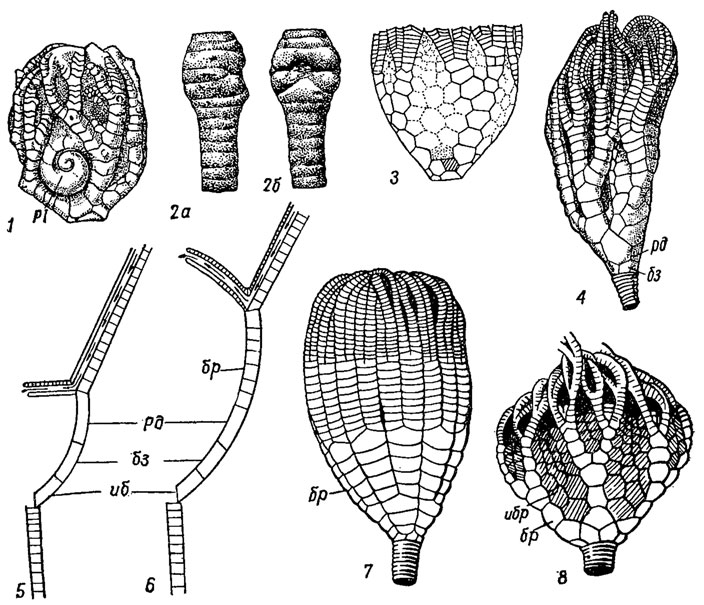
Рис. 271. Подкласс Flexibilia: 1 - чашечка Taxocrinus из девона с раковиной брюхоногого моллюска Platyceras (Pl); 2а, б - часть стебля из карбона, поврежденного паразитирующими мизостомидами (Annelides); 3,8 - Sagenocrinites (силур), в состав чашечки входят брахиальные (бр) и интербрахиальные (ибр) таблички; 4 - Protaxocri- nus (средний ордовик - силур); 5-6 - увеличение размеров чашечки за счет брахиальных табличек; 7 - Ichthyocrinus (силур - ранний девон), в состав чашечки входят брахиальные таблички; (обозначения см. рис. 263)
История развития криноидей. Происхождение криноидей до сих пор не выяснено. Предполагают, что они обособились в кембрии от общего предка с дистоидеями и их развитие было связано с возникновением радиальных выростов тела - рук, предназначенных для сбора пищи. Руки не гомологичны брахиолам цистоидей и бластоидей. В раннем ордовике становятся известными представители двух подклассов: камерат и инадунат, а начиная со среднего ордовика - подкласс флексибилий. Если первые два подкласса образуют дивергирующие группы, то подкласс флексибилий остается на протяжении палеозоя небольшой группой, вымирающей в середине перми. Особенно многочисленны и разнообразны были камераты и инадунаты в девоне и раннем карбоне. Число камерат резко сокращается к концу карбона, и последние представители этого подкласса вымирают в середине перми. Инадунаты, наоборот, в перми дают новую вспышку и характеризуются довольно широким распространением. Один из подотрядов инадунат (энкринина) сохраняется в триасовое время, но и он к концу триаса вымирает. Первые представители подкласса артикулят появляются в триасе; в юре и мелу они становятся многочисленными; среди них наряду с прикрепленными стебельчатыми формами появляются бесстебельчатые подвижные криноидеи. В современных морях артикуляты стебельчатые (75 видов) и бесстебельчатые (свыше 500 видов) являются единственными представителями некогда обширного не только класса криноидей, но и всего подтипа кринозой.
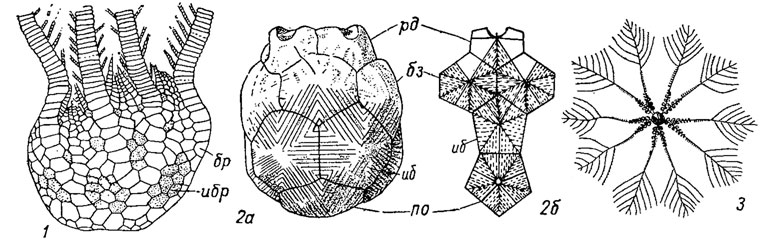
Рис. 272. Подкласс Articulata: 1 - Uintacrinus (поздний мел); 2а, б - Marsupites (поздний мел); 3 - Saccocoma (поздняя юра - мел); таблички: ибр - интербрахиальные, по - пятиугольная табличка основания (остальные обозначения см. рис. 263)
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'