
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Часть вторая. Принципы и направления исследований тонкой структуры и вещественного состава ископаемых скелетных образований
2.1. Основные вопросы биоминерализации (С. Н. Голубев)
Известно, что кости, раковины и другие скелетные образования состоят из природных композиционных материалов, являются комплексом биополимеров и ионных кристаллов (в подавляющем большинстве случаев карбонатов или фосфатов кальция). В составе минеральной части скелета низших организмов широко распространен аморфный диоксид кремния. В минерализованных структурах разных организмов (от одноклеточных до млекопитающих) состав органического компонента с точностью до групп соединений одинаков. Это белковомукополисахаридный комплекс, куда входят фибриллярные структурные белки (коллаген или сходные с ним белки). Мукополисахариды иногда представлены хитином, чаще это другие соединения, иногда их называют хитиноидными. Сам термин "хитин" имеет два значения: 1) в узком смысле - индивидуальное вещество из группы мукополисахаридов; 2) в широком смысле - материал, из которого состоит панцирь членистоногих (фактически это комплекс, включающий собственно хитин и структурные белки). Материал панциря членистоногих является неплохой иллюстрацией того, что такое органический компонент любого скелета*.
* (Здесь и далее для краткости понятие "скелета" включает минерализованные трубки червей, минерализованные выделения прокариот и другие аналогичные образования.)
Пространственные соотношения между органической и минеральной фазами скелета. Тонкая структура органических матриц подобна трехмерной сетке: в одном направлении вытянуты молекулы белка, двум другим соответствует субдвумерная мукополисахаридная сетка [1087]. Полисахаридная "сетка" состоит из дискретных пластин (см. 2.2). На определенной стадии внутри белковых фибрилл возникают кристаллические зародыши, причем взаимная ориентация кристаллографических осей хаотична, соответствующая упорядоченность возникает только на последующих стадиях процесса [1640]. Заметим, что такая же картина наблюдается и при типовой эпитаксии небиологических материалов. Эпитаксия - направленная кристаллизация одного вещества, определяемая частицами другого. Необходимым ее условием является определенное стереохимическое соответствие между компонентами.
Расположение кристаллических зародышей при биоминерализации подчиняется четкой упорядоченности. В случае коллагена считается установленным, что зародыши формируются, как показано на рис. 56 [1641, 2395]. В действительности соотношения, видимо, сложнее; во всяком случае интерпретация ряда моментов неоднозначна. Возникновение зародышей в зонах, которые, по существу, являются субмолекулярными "порами", обеспечивает важный момент: на ранней, самой уязвимой стадии процесса кристаллиты минимально деформируют тонкую структуру коллагена [2395], на последующих стадиях минерализации такая деформация неизбежна. Подчеркнем, что когда минерализация только начинается, внутри объема органической матрицы еще нет тех "пустот", в которых могли бы уместиться полностью кристаллиты. Процессы собственно кристаллизации и процессы "освобождения места" в органической матрице идут почти синхронно. Таким образом, биоминерализацию можно представить как результат "замещения" части вещества органической матрицы минеральными кристаллами.
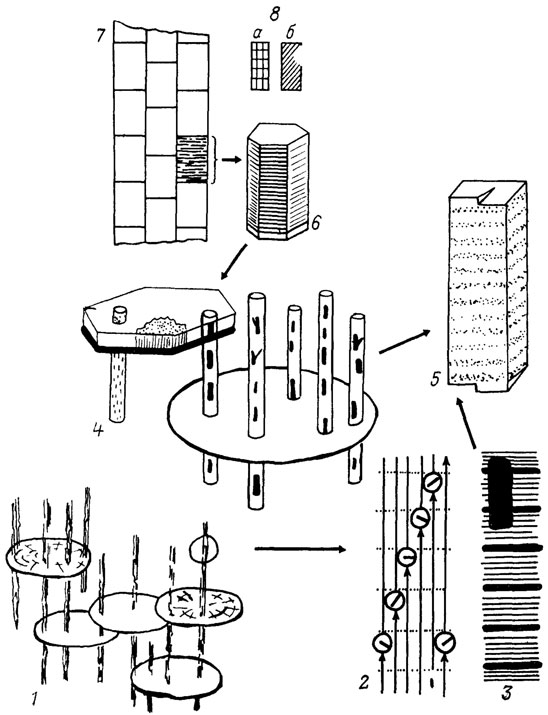
Рис. 56. Принципиальная схема биоминерализации. По С. Н. Голубеву. 1 - органическая матрица до начала минерализации, на двух пластинах показана вероятная ориентировка молекул, ув. 20 000; 2-3 - коллагеновая фибрилла, ув. 200000; (2 - один из классических вариантов интерпретации, длинные стрелки - молекулы тропоколлагена, в кружочках кристаллические зародыши; 3 - поперечная исчерченность, фактически наблюдаемая картина - один полностью сформированный элементарный кристаллит); 4 - справа кристаллиты (их ширина несколько преувеличена) внутри белковых фибрилл, раздвоенные кристаллиты - двойники, слева - средняя стадия минерализации; на базе гексагональной пластины возник поликристалл, ув. 50000; 5 - элементарный кристаллит, вдоль оси с чередуются зоны с повышенной и пониженной концентрациями дефектов; на торцах видна винтовая дислокация, спираль роста, ув. 500000; 6 - 'стопка' гексагональных пластинчатых поликристаллов, элементарные кристаллиты не различимы; 7 - фрагмент сечения условного скелета, тонкая штриховка не совпадает с направлением удлинения элементарных кристаллитов, ув. 100000; 8 - тот же фрагмент, что на позиции 7; ув. 2000; 8, а - неполяризованный свет; 8, б - скрещенные николи, объект в положении погасания
В подавляющем большинстве случаев рост кристаллов останавливается на той стадии, когда между соседними кристаллитами остаются органические мембраноподобные пленки. В результате возникает структура, свойственная большинству скелетов и аналогичная кладке кирпичей, скрепленных цементом: соседние кристаллиты не срастаются между собой непосредственно, а соединены в единое твердое тело (кость, раковину и т. д.) "цементирующей пленкой" органической матрицы (см. рис. 56). При этом, например, прочность кости определяется не только параметрами идеализированной решетки кристаллов и особенностями геометрии системы, но и энергией специфического взаимодействия между органической и минеральной частями скелета. Точная природа этого взаимодействия неизвестна, но представление о существовании специфического комплексообразования на границе раздела является общепризнанным.
Элементарные кристаллиты, т. е. минимальные структурные единицы, имеющие обволакивающую органическую пленку, геометрически являются сильно вытянутыми пластинками. Их размер по порядку величин одинаков у самых разных организмов: длина - около 100 нм, ширина - десятки нанометров, толщина - около 5 нм. Как правило, толщина органических пленок, разделяющих элементарные кристаллиты, достигает нескольких нанометров [2395].
Массовая доля органической части при таких соотношениях составляет ориентировочно 20-30%, что соответствует фактической ситуации для большинства скелетов. Однако известны минерализованные структуры и с гораздо меньшим содержанием органического вещества - например, скелеты некоторых иглокожих и эмаль зуба млекопитающих [2395]. Их минерализация на ранних стадиях идет в полном соответствии со сказанным выше, но потом рост кристаллов не останавливается до тех пор, пока органическая матрица практически полностью не будет "вытеснена". Так, в эмали сформированного зуба массовая доля органического вещества составляет 0,5-1,5% [2220]. В данном случае функциональный смысл явления понятен: органические пленочные прослойки недопустимо снижают твердость эмали. Однако за такое повышение твердости организму приходится "расплачиваться". Лишаясь органической части, эмаль становится структурой, гораздо более хрупкой, чем обычная кость. Помимо этого эмаль в прижизненном состоянии - практически мертвое образование, она не обновляется и не способна к регенерации. Однако большинство минерализованных объектов подчиняется принципу "обволакивающих пленок". Но надо четко понимать, что нет механизмов, которые бы индивидуально синтезировали органическую пленку вокруг каждого кристаллита. Онтогенетически пленки соответствуют той части органического вещества, которая не была замещена растущими минеральными кристаллами. По отношению к кристаллам эти пленки являются не новообразованием, а реликтом ранних стадий процесса.
Скелетам свойственно наличие нескольких соподчиненных уровней структуры (см. рис. 56). Например, раковина состоит из слоев, слои - из призм, призмы - из ламеллей, последние - из фибр и т. д. Конкретные соотношения, само количество иерархических уровней, а также исторически сложившаяся терминология для разных организмов неодинаковы. При формировании относительно крупных (микронных, миллиметровых, а иногда и сантиметровых) структурных единиц между кристаллографическими осями элементарных кристаллитов реализуются вполне определенные соотношения, в простейшем случае одноименные оси взаимнопараллельны. При этом, например, в поляризационном микроскопе такие крупные структуры могут проявлять свойства монокристаллов.
Реальные кристаллы и биоминерализация. В изучении кристаллов имеется, грубо говоря, две стадии. На первой стадии исследуется идеализированная решетка того или иного кристаллического вещества, на второй стадии изучаются разноплановые дефекты кристаллической решетки. Практически соответствующие работы ведутся разными специалистами по разным методикам. Так получилось, что автор впервые начал проводить изучение реальных кристаллов биологических скелетов, т. е. такие исследования, при которых конкретно рассматривалась неидеальность кристаллов. Для этого использовались специальные методы электронной микроскопии, принципиально неприменимые в обычной биологии мягких тканей. Работы проводились на примере кокколитов как из культуры водорослей, так и из осадков [253]. Ниже приведены некоторые выводы, причем без конкретного экспериментального обоснования, т. е. без объяснения того, каким способом соответствующие детали структуры можно сделать видимыми в электронном микроскопе (это совсем не те методы, которые получили широкое распространение в современной палеонтологии).
Субигольчатые кальцитовые кристаллы кокколитов характеризуются специфической периодичностью концентрации дефектов. Вдоль длинной оси кристалла (ось "с", совпадающая с направлением оси белковой фибриллы) чередуются блоки, примерно по 50-60 нм каждый, с относительно пониженной и повышенной концентрациями дефектов. Практически несомненно, что в такой форме кристалл наследует тонкую поперечную исчерченность коллагена, которая сама по себе, вероятно, тоже связана с чередованием участков разной степени упорядоченности [1107]. В костях человека вдоль того же направления существуют блоки (размером в те же 50 нм); их природа неизвестна [2220]. По-видимому, природа этой структурированности у кокколитофирид и у человека одна и га же, а совпадение размеров блоков просто отражает сходство коллагенов у столь разных организмов. В рамках ранее применявшихся методов эти блоки выявлялись в костях тогда, когда при ультрамикротомировании костей возникали трещины, косвенно отражающие реальную структуру кристаллов.
У кокколитофорид изучена разновидность скелетной патологии, развитие которой связано с нарушением периодичности в 100 нм и с появлением биологически аномальных кольцевых дислокаций. Непосредственно наблюдались различные степени развития такой патологии. Современное состояние проблемы парадоксально: известно не менее 100 видов патологии костной ткани человека, а исследования такого типа пока проводились только на одноклеточных водорослях. На самом деле это совершенно естественно: микронные кокколиты наиболее пригодны для электронно-микроскопических исследований реальной структуры кристаллов примерно так же, как почти каждое направление в молекулярной биологии начинается с изучения бактерий.
Широко известно, что рост обычных кристаллов происходит по спирали (винтовой дислокации). Определенные данные говорят о том, что спираль коллагена предопределяет возникновение главной винтовой дислокации кристаллов - спирали роста. Если это так, то становится понятным давно известный факт: усовидные кристаллиты вытянуты вдоль направления белковых фибрилл. Другими словами, кристалл растет по спирали, унаследованной от коллагена; растет вдоль оси коллагеновой фибриллы и становится субигольчатым, вытянутым вдоль этой оси.
Двойникование биогенных кристаллов. Кристаллы многих веществ образуют особые сростки, так называемые двойники. Более сложные сростки называются тройниками, еще более сложные - полисинтетическими двойниками. Двойникование наблюдается во всех скелетах. Вероятно, раньше всего это явление изучалось на моллюсках [1994], а с наибольшей детальностью - на примере некоторых кокколитов [253]. Кроме кокколитов двойникование изучалось нами на скелетах червей, литотамниевых водорослях, на костях человека и на ряде других объектов [254].
Структурная роль двойникования при биоминерализации чрезвычайно велика: двойникование выступает как фактор сопряжения относительно правильной кристаллографической формы с формой биологических образований, всякого рода овалов, субсферических структур и т. д. (рис. 57). Примечательно, что на примере дискоастеров (ископаемых остатков, сближаемых с кокколитами) нам удалось показать, что благодаря полисинтетическому двойникованию возникают сростки с пятилучевой симметрией. Как известно, ни в каких кристаллах невозможна ось симметрии пятого порядка; к сросткам это ограничение не относится, но соответствующие структуры маловероятны для небиологических систем и практически не встречаются. Добавим, что имеются теоретические расчеты, объясняющие появление округленных контуров в системах геометрически сложного двойникования микроскопических реальных кристаллов и небиологической природы [513]. Следовательно, организмы при биоминерализации осуществляют сопряжение кристаллографически правильной формы с криволинейными контурами типичных биологических структур.
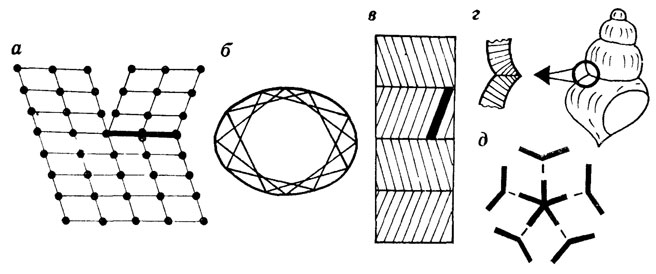
Рис. 57. Двойникование кристаллов. По С. Н. Голубеву. а - один из вариантов двойникования кристаллов кальцита, точки - катионы кальция анионы не показаны, б-д - двойникование как фактор сопряжения относительно правильной кристаллографической формы со специфической формой скелетных структур б - фрагмент определенного кокколита, сдвойникованные ромбоэдрические пластины обеспечивают появление эллиптических контуров, в действительности имеются еще и более мелкие 'вспомогательные' элементы, в - принципиальная схема возникновения перекрещенно пластинчатого слоя, г - двойникование кристаллов на стыке оборотов позволяет построить спирально коническую раковину, д - 'наложение' пяти сдвоиникованных структур приводит к появлению скелетных элементов с пятилучевой симметрией
Потенциальные формообразующие возможности скелетного образования, в котором одноименные кристаллографические оси всех кристаллитов параллельны друг другу, весьма ограничены. Кроме того, прочностные характеристики такого скелета несовершенны материал неизбежно получится волокнистый, легкорасщепляющийся. Природа преодолевает эти сложности, используя явление двойникования. Из-за двойникования в трехмерной структуре скелета всегда есть усовидные кристаллиты, расположенные под углом к основному "пучку" (см. рис. 56, 4; идеализированный вариант - см. рис. 56, 7). В результате формируется скелет с весьма совершенными прочностными характеристиками.
Поскольку углы взаимного поворота кристаллитов не произвольны, а соответствуют законам двойникования, то суммарная структура строго упорядочена и энергетически весьма выгодна. Последнее не надо понимать только в смысле экономии энергии при биоминерализации. Существует фундаментальный физико-химический принцип: любая система стремится занять положение с возможным минимумом потенциальной энергии. Любой спонтанный природный процесс направлен в сторону понижения общей потенциальной энергии; соответствующим образом ведут себя механические пружины, магниты, химические процессы и т. д.
Энергетическая выгодность микроструктурной системы скелета означает: 1) построение скелета в конечном итоге приводит к минимизации потенциальной энергии, при прочих равных условиях такой процесс легче "автоматизировать"; 2) если потенциальная энергия скелета минимальна, то автоматически исключается возможность биологически нецелесообразных перестроек: такая перестройка приводила бы к повышению потенциальной энергии и как спонтанный процесс невозможна. Если организму понадобится перестроить скелет, то он сможет это сделать, используя внешние источники энергии [253]. Состояние, энергетически выгодное при одних условиях, может стать невыгодным при других, поэтому непосредственно распространять характеристики прижизненной устойчивости скелета на процессы фоссилизации нельзя, но определенная взаимосвязь соответствующих закономерностей существует.
Кристаллы одного и того же вещества, в частности кальцита, могут двойниковаться несколькими геометрическими способами. Соответственно трехмерная структура полисинтетического двойника может быть очень сложной. Для определенных кокколитов весьма экзотической формы, состоящих из огромного числа кристаллитов, нам удалось показать, что весь кокколит является единым полисинтетическим двойником. Сейчас мы подошли вплотную к пониманию того, что вся суммарная структура макроскопического скелетного образования (например, типа раковины гастроподы) может быть представлена как единый сложный полисинтетический двойник. Если это так, то появится возможность конкретизировать параметры рациональности микроструктур различных скелетов. В этом случае несомненно, что в некоторых группах изменятся представления о признаках прогрессивности. В частности, общепринятая точка зрения о примитивности микрогранулярной структуры восходит к доэлектронно-микроскопическим временам, когда эта структура казалась хаотичной. В свете сказанного микрогранулярную структуру в том виде, в каком она предстает при детальных электронно-микроскопических исследованиях, надо признать одной из самых высокоупорядоченных.
В то же время двойникование биогенных кристаллов - явление специфическое, лишь частично аналогичное двойникованию "обычных" кристаллов. При двойниковании небиогенных кристаллов границами двойников являются если и не атомные плоскости, то области соизмеримой толщины. В скелетах же сдвойникованные кристаллы разделены органической пленкой. Энергетическая выгодность такого необычного двойникования обусловлена наличием дальнодействующей упорядоченности; нечто похожее реализуется в системах жидких кристаллов. Геометрическая и размерная специфика кокколитов позволила прямым наблюдением показать фактическое существование дальнодействующей упорядоченности (на уровне дислокаций решетки) в системе несоприкасающихся кристаллитов, показать ощутимость соответствующих физических сил и реальность того вклада, который привносится в энергетическую выгодность системы существованием дальнего порядка [253].
Физико-химический аспект типовой фоссилизации. На материале культуры водорослей и из океанических осадков изучались кокколиты одного биологического вида. При переходе этих скелетов в ископаемое состояние происходит диффузионное выравнивание концентрации дефектов по объему кристаллитов, исходная периодичность в 100 нм реальной структуры исчезает. При этом возрастает устойчивость скелета к растворению: при исходной структуре собственно растворение кокколита сопровождается его распадом на субколлоидные частицы, в роли которых выступают блоки с пониженной концентрацией дефектов. Эти блоки растворяются медленнее и остаются после того, как кокколит перестает существовать в качестве единого твердого тела. Фактическая устойчивость скелета как морфологически опознаваемой структуры контролируется блоками с повышенной концентрацией дефектов, а они при фоссилизации исчезают. Подчеркнем, что такое повышение устойчивости к растворению является чисто твердофазным процессом и не имеет ничего общего с другим процессом - иногда наблюдавшимся химическим замещением кальцита. Все сказанное продемонстрировано как в прямых опытах, так и в модельных экспериментах [253].
Другая важная закономерность, сопровождающая переход скелета в ископаемое состояние, связана с полным или частичным (в зависимости от места нахождения) разрушением органической части скелета. В тех случаях когда матрица разрушается полностью, происходит нарушение исходной структуры и соседние кристаллиты непосредственно срастаются; идет медленный процесс, который в некотором роде аналогичен методу спекания порошков в высокотемпературной технологии. При частичном разрушении матриц тонкая молекулярная структура меняется, происходит обугливание органики, ее дубление. Правда, процесс является статистическим, некоторая доля молекул может сохраняться и в первичной форме [1422]. Ясно, что в прижизненном состоянии система органических структур скелета, система "обволакивающих пленок" должна функционировать как своеобразная система подводящих каналов; в противном случае кристаллы просто не смогли бы вырасти. При переходе скелета в ископаемое состояние такая "система каналов" исчезает или теряет работоспособность, из-за чего во много раз снижается скорость обменных химических реакций (как следствие изотопно-обменных процессов). Причем, как показали прямые эксперименты, эффект проявляется во много раз сильнее там, где органические структуры сохраняются. Они сохраняются в обугленном виде, а в такой форме соответствующие пленки служат дополнительными границами раздела, снижающими скорость диффузии в твердом теле, по сравнению с прижизненной ситуацией все зеркально меняется с исключительной точностью.
Две указанные закономерности фоссилизации имеют как бы внутреннюю по отношению к скелету физическую природу, а потому реализуются в очень разных геологических условиях. К этому может добавляться много частных процессов, специфичных для условий того или иного местонахождения. Достаточно ясно, что рассмотренные закономерности имеют значение не только для кокколитов. Но очевидно и обратное: на этом уровне между скелетами разных организмов должны существовать тонкие различия. Становится хотя бы в принципе понятным, почему в одних и тех же условиях скелеты разных организмов сохраняются в химическом смысле неодинаково - сам факт этого общеизвестен (разумеется, речь идет о скелетах с одинаковым составом минеральной части).
Биоминерализация у разных организмов - единство основных закономерностей. Вероятно, что при первом ознакомлении с принципом "обволакивающих пленок" трудно преодолеть впечатление, что это надуманная абстракция. Ведь речь идет уже не только о детальной морфологии как таковой, а о характере взаимодействия кристаллитов, о способе их соединения в твердое тело.
Если кристаллиты "скреплены" обволакивающей пленкой, то при разрушении матрицы скелет должен просто рассыпаться на отдельные кристаллиты. Почему же этого не происходит при фоссилизации скелета? Дело в кинетических соотношениях: при фоссилизации матрица разрушается медленно и кристаллиты успевают срастись между собой. Если же осуществить быстрое химическое разрушение матриц, то скелеты действительно рассыпаются, что прямо доказывает реальность принципа обволакивающих пленок [253]. Этот принцип отражает одну из самых общих и важных закономерностей биоминерализации. Он применим к скелетам с разным составом минеральной части. По нашим данным, этому принципу подчиняются минерализованные структуры самих организмов - от прокариот до млекопитающих. Известные исключения (типа эмали зуба) формируются также в соответствии с принципом обволакивающих пленок и перестают ему подчиняться только на последних стадиях процесса.
Распад скелета при разрушении органических матриц использован нами как основа нового электронно-микроскопического подхода к исследованию скелетов. Естественно, обособленные фрагменты, на которые рассыпаются минерализованные структуры, были изучены разными методами электронной микроскопии [254]. Изучены 15 объектов, включая минеральные выделения сине-зеленых водорослей, скелеты литотамниевых водорослей, червей, брахиопод, моллюсков, членистоногих, кости человека, панцирь черепахи и др. Малой выборкой, на уровне отдельных представителей охвачены минерализованные структуры с разным составом, принципиально разной морфологией и таксономической принадлежностью. Во всех объектах выявились субигольчатые кристаллиты и их агрегаты. Специфика метода позволяет исключительно четко обнаруживать кристаллические двойники, однозначно регистрируются случаи агглютинации. Литотамниевые водоросли включают в свой скелет посторонние кокколиты, черви - панцири диатомей; в обоих случаях массовая доля таких включений ничтожна, другими методами включения обнаруживаются с трудом. Наиболее интересным и неожиданным результатом оказалось обнаружение в составе органических матриц дискретных полисахаридных пластин, устойчивых к реактивам, применявшимся для разрушения скелетов (реактивы разрушают в основном белковый компонент матрицы). Пластины обнаружены во всех минерализованных структурах организмов (от прокариот до человека), и на весь этот набор приходятся только два морфологических варианта пластин. Причем пластины каждого варианта неотличимы друг от друга, хотя и извлечены из минерализованных структур организмов, относящихся к разным типам и даже более высоким таксонам. Установлены следующие морфологические варианты.
1. Эллиптические пластины длиной около 1,5 мкм, эксцентриситет эллипсов с высокой точностью равен 1/√2. В центре располагается структура в форме рельефной "нашлепки", в плане это тоже эллипс, но с эксцентриситетом 1/2, причем этот эллипс проходит через фокальные точки предыдущего (структурная роль пластин показана на рис. 56). Иногда пластины извлекаются упорядочено перфорированными, остается как бы след от белковых фибрилл и кристаллов. Пластины получены из минеральных выделений сине-зеленых водорослей ("современные строматолиты") - культура Института микробиологии АН СССР, из скелетов красных литотамниевых водорослей, из трубки полихеты, из раковины замковой брахиоподы Pictothyris sp., из чешуи костистой рыбы Hemichrompis bimaculatus, из панциря степной черепахи и из эмали зуба человека. В двух последних объектах эллиптические пластины сосуществуют с пластинами другого типа, описываемого ниже. Эллиптические пластины электронографически аморфны.
2. Пластины от отчетливо гексагональных до субокруглых размером около 1 мкм. Резко отличаются от вышеописанных кристалличностью (по данным электронной микродифракции). Выделены из "губчатого" слоя раковины сепии, раздельно из перламутра и призматического слоя митилуса, из панциря балянуса, из крышки гастроподы Astrea rugosa, из панциря черепахи (как и в случае эллиптических пластин - маргинальные элементы), из эмали зуба человека (распознаваемые фрагменты получены из недоминерализованного зуба), из разных костей человека и некоторых других объектов. Пластины этого типа в полностью деминерализованной форме дают отчетливые электронно-графические рефлексы, т. е., будучи органическими, обладают собственной кристалличностью.
Определение межплоскостных расстояний показало, что между веществом пластин и минеральными кристаллами реализуются условия эпитаксии: вдоль определенных кристаллографических направлений межплоскостные расстояния совпадают. В кальцитовых скелетах такое совпадение реализуется для плоскостей {202̄5} с расстоянием 0,158 нм, в арагонитовых - для плоскостей {311} с расстоянием 0,156 нм. Цифры получены прямым измерением по матричным пластинам, но находятся на грани точности метода, поэтому не будем забывать о белковых фибриллах, разрушенных при извлечении пластин. Там, вероятно, реализуются и другие эпитаксиальные соотношения. В оксиапатитовых скелетах пластины имеют межплоскостные расстояния 0,089 и 0,135 нм, что соответствует половинным расстояниям апатитовых плоскостей {325̄1} и {303̄0}. Исследование идентичными методами костной саркомы человека показало, что в пораженной области имеются гексагональные пластины, внешне неотличимые от нормальных, но с аномальными электронно-графическими характеристиками. Условия эпитаксии нарушены, саркома характеризуется субхаотичностью в ориентации кристаллов.
Кремневые скелеты изучены на примере двух видов губок Euplectella aspergilina и Hialospongia sp. Обнаружились гексагональные пластины с теми же межплоскостными расстояниями, что и в случае апатитовой минерализации. Но по каким-то причинам организмы поставляют на пластины другие растворы, осаждают диоксид кремния. При этом оказывается, что те самые межплоскостные расстояния, по которым реализуется эпитаксия при апатитовой минерализации, есть и у кристобалита: 0,178 нм для {325̄1} апатита и 0,178 нм для {004} кристобалита; 0,270 нм для {303̄0} апатита и 0,133 нм (практически половина) для {105} кристобалита. Расстояния совпадают, но настоящая эпитаксия невозможна - ориентация плоскостей резко различна; более того, апатит гексагонален, кристобалит тетрагонален. Указанное минералогическое совпадение обеспечивает биологические механизмы управляемого формирования аморфного тела. При посмертной кристаллизации таких скелетов должен возникать именно кристобалит, а не кварц, что и происходит в океанических осадках.
Видимо, никто не подозревал, что дискретные эллиптические пластины можно извлечь, скажем, из человеческого зубы. Но у водорослей они известны. Пластины формируются в цистернах комплекса Гольджи и являются дискретными фрагментами клеточной стенки. Если стенка минерализуется (как у кокколитофорид), то пластины входят в состав скелетных матриц. Такой механизм формирования клеточной стенки свойствен водорослям, грибам и высшим растениям. Данные по животным ограничены клетками пищеварительной системы крыс.
В целом эволюционные корни биоминерализации прослеживаются вплоть до механизмов формирования клеточной стенки.
Пластины разных организмов химически заведомо нетождественны: иногда это хитин, чаще другие полисахариды. Вместе с тем ясно, что обнаружена закономерность, общая для организмов, от прокариот до человека. Общность закономерности есть определенный принцип работы биофизической "машины", он допускает наличие вариантов состава как матриц, так и минерального компонента. В сфере биоминерализации, как и в других биологических системах, есть барьер минимальной сложности, ниже которого процесс невозможен. При биоминерализации этот барьер определяется необходимостью присутствия механизмов, останавливающих рост кристаллов в нужный момент. Достаточно упомянуть разнообразные патологические "отложения солей" у человека, чтобы представить, что произошло бы с организмом бактериального уровня, если бы у него не было механизмов тормозящего действия. На этой несколько парадоксальной стороне биоминерализации, являющейся развитием более общей идеи Н. В. Белова о биологических механизмах защиты от "кристаллической смерти", мы уже останавливались, причем с привлечением экспериментального доказательства существования такого тормозного механизма у кокколитофорид [254].
Мы рассмотрели лишь некоторые аспекты проблемы биоминерализации. В виде единой системы представлений удалось проанализировать формирование скелета, его прижизненное состояние, возникновение определенной патологии, общие закономерности фоссилизации. На этой же базе были созданы новые экспериментальные подходы и неоднократно осуществлялось правильное предсказание результатов их применения. Данные о полисахаридных пластинах - первые сведения морфологического характера о существовании серьезного сходства между скелетами и скелетоподобными структурами организмов разных типов и даже царств. Для классической палеонтологии между "скелетами" литотамниевых водорослей и, например, раковинами брахиопод нет ничего общего. Фактически из-за этого изучение древних и современных скелетов мало что дает для выяснения эволюционных соотношений на уровне высших таксонов. При фрагментарности современных данных делать обобщения нельзя, но более чем вероятно, что на основании большого количества фактических данных по скелетным материалам могут быть получены сведения о эволюционных соотношениях высших таксонов. Причем эти сведения послужат совершенно независимой проверкой существующих представлений.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'