
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
1.8. Палеонтологические проблематики (В. Н. Шиманский)
Геологу и палеонтологу довольно часто приходится иметь дело с объектами, систематическая принадлежность которых к той или иной группе организмов неясна и более того - даже не всегда доказуема вообще принадлежность к остаткам организмов. Все подобные остатки принято называть проблематикой. Существует, однако, несколько разных пониманий этого термина. Большинство палеонтологов понимает под проблематикой любые остатки организмов, для которых нельзя указать точной принадлежности к тому или иному классу и даже типу. Это могут быть единичные, совершенно непонятные формы, иногда части скелета, не дающие полного представления о целом скелете; отдельные роды, иногда группы родов, условно объединяемые в семейства и даже отряды неизвестного систематического положения. При таком понимании проблематики в нее входят также различные следы жизнедеятельности организмов (следы ползания, норки и т. д.). Это понимание столь широко вошло в науку, что выпускаются даже целые сборники, содержащие описание различных групп организмов, в названии которых стоит слово "проблематика" - например, "Проблематики палеозоя и мезозоя" и "Проблематики фанерозоя" [843, 844].
Существует и иная трактовка этого понятия. Так, в "Палеонтологическом словаре" [799, с. 263] сказано, что "проблематика" является синонимом понятия "проблематические остатки", а последним дается следующее объяснение: "Проблематические остатки (от греч. problematicos - затруднительный, спорный) - ископаемые, природа и происхождение которых неясны". К ним относятся большинство следов жизнедеятельности организмов, а также скелетные остатки неопределенного происхождения. В некоторых случаях сомнительно даже органическое происхождение некоторых таких "остатков". При таком понимании, очевидно, нельзя считать проблематикой целые группы организмов, для которых неясно только их место в системе органического мира. В качестве примера можно указать хиолиты, археоциаты, ринхолиты (в принадлежности которых к головоногим моллюскам никто не сомневается) и т. д.
Еще более строгое определение мы находим в "Геологическом словаре" [236, с. 184], где сказано: "Проблематические остатки (образования) ... ископаемые, природа которых и происхождение неизвестны. Относятся частью к фитоморфозам и фукоидам. Известны из всех осадочных отложений. Многие проблематические остатки после изучения были отнесены к ископаемым растениям или животным".
Наконец, в "Толковом словаре английских геологических терминов" практически отрицается возможность называть "проблематиками" группы организмов и даже вообще организмы, для которых неясно их систематическое положение. В словаре имеются два термина - "problematic fossil" и "problematicum". Сущность первого сформулирована так [1068, с. 562]: "Природные объекты, осадочные текстуры или знаки в породах, похожие на окаменелости, но органическая природа которых очень сомнительна". Под "problematicum" (там же) понимаются: "Следы, объекты, осадочные текстуры или другие образования (в породах), природа которых проблематична, напр. сомнительные "окаменелости", которые, возможно, имеют неорганическое происхождение или органическая природа которых не доказана, особ, органические остатки (такие, как следы жизнедеятельности) более или менее неясной природы".
Правда, в англоязычных палеонтологических работах последних лет термин "проблематика" применяется также в очень широком понимании, т. е. под проблематикой понимаются и "непонятные" группы организмов.
В связи с двойственностью понимания термина автором настоящей главы еще в 50-х годах была сделана попытка подразделять остатки неясного систематического положения на "проблематические" и "полупроблематические" [1166]. К последним, в частности, были отнесены ринхолиты, т. е. организмы, в принадлежности которых к головоногим моллюскам никто не сомневался, но данные о точной их принадлежности к определенной группе (к наутилоидеям, аммоноидеям и т. д.) отсутствовали.
Видимо, следует различать несколько понятий. Термин "проблематика" желательно сохранить для любых образований, природа и происхождение которых неизвестны. Это могут быть неорганические объекты, напоминающие образования животного и растительного происхождения, несомненные остатки организмов, о которых, однако (в силу их фрагментарности, очень плохой сохранности и т. п.), нельзя сказать ничего определенного ("нельзя их диагностировать"), а также различные следы жизнедеятельности организмов.
Как видим, в этом комплексе могут оказаться совершенно различные объекты. Довольно часто среди них окажутся "псевдофоссилии", т. е. неорганические образования, очень похожие на остатки животных и растений (иногда, например, небольшие сталактиты, по внешней форме и на пришлифовках весьма сильно напоминающие ростры белемнитов). Иногда в руки исследователя попадают разрозненные элементы скелетных остатков или отдельных частей неизвестных организмов (разрозненные небольшие таблички, капсулы, возможно, яйцевые). Как правило, такому остатку не дается специального видового названия и в дальнейшем сведения о подобной находке могут полностью потеряться в огромном потоке информации. Вероятно, так было "потеряно" достаточно много уникальных объектов.
Особой категорией являются многочисленные следы жизнедеятельности организмов. Они представляют собой предмет исследований ихнологии - особого раздела палеонтологии. Для таких образований предложена даже особая классификация, не сопоставимая с классификацией, принятой для собственно животных организмов [208]. Подробнее эта проблема рассмотрена в специальном разделе (см. 3.2).
Следует ввести термин "загадочные организмы"* для остатков, в растительном или животном происхождении которых никто не сомневается и которые даже рассматриваются в определенном систематическом ранге (род, семейство, отряд и др.), но положение их в системе остается неизвестным. Правда, возникает вопрос о границах понимания "загадочных" групп, т. е. до какого же уровня группу следует считать "загадочной". Очевидно, следует различать несколько уровней "загадочности". К одному отнести формы, для которых установлен вид и род, но от дальнейших выводов исследователи отказываются. В качестве примера можно указать совершенно невероятное животное Tullimonstrum, имеющее довольно крупное овальное тело с двумя клешнями на переднем конце и короткими, симметричными выростами вдоль "боков" (рис. 51). Интересно, что его остатки, притом в довольно значительном количестве, были найдены сравнительно недавно в каменноугольных отложениях Северной Америки. Даже после детальных исследований специалисты не могут отнести его к какому-либо из известных типов.
* (С. В. Мейен предложил для "загадочных" групп термин "энигматы", или "энигматики" (от лат. aenigma - необъясненное, загадочное).)
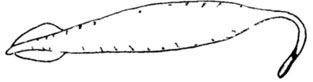
Рис. 51. Одна из возможных реконструкций Tullimonstrum gregarium Rich. Ув. 0,5. По М. У. Фостеру
К другому уровню должны быть отнесены формы семейственной и отрядной групп, как правило, включающих по нескольку таксонов подчиненного ранга. В данном случае уже имеются какие-то представления об иерархии таксонов, о более важных особенностях организмов, объединяющих их в группу, и о менее важных - характерных для родов и видов. Однако для отнесения таких групп к тем или другим классам и типам материала недостаточно или близких форм в известных уже классах и типах нет. В качестве примеров таких групп укажем Cornulitidae или Cornulida (семейство или отряд "червей", тентакулитов или кишечнополостных из ордовика - карбона), Coleolidae или Coleolida (семейство или отряд "червей", моллюсков из кембрия - карбона), Hyolithelmintes (отряд предположительно "червей" из кембрия), Moravimminida (отряд микроорганизмов из триаса). К этим двум группам практически относятся почти все остатки из докембрия, так как отнесение их к определенным типам животных весьма условно, а чаще же не делается совершенно. Видимо, это особый мир, который еще предстоит изучать и систематизировать.
Достаточно близка к этому уровню категория групп, для которых установлены даже классы, а иногда и типы, но положение которых в системе органического мира остается неясным. К таковым относятся Rangeomorpha, Pteridiomorpha, Erniettomorpha (классы в гипотетическом типе Petaloname; самые верхи докембрия или низы кембрия); Archaeocyatha (тип; кембрий), Aphrosalpingoida (класс; силур), Cribricyatha (класс: кембрия), Radiocyatha (класс, кембрий), Receptaculita (в последние годы рассматривается в ранге класса или типа; ордовик - пермь), Soanitida (в последнее время ранг поднят до класса; ордовик).
Относительно петалонам существует два диаметрально противоположных суждения. По мнению исследователя, установившего классы и тип [2083], это совершенно особая группа, включающая довольно сложноорганизованные существа, являющиеся предками ряда более поздних ветвей животного мира. По мнению других, формы, относимые к петалонамам, по развитию стоят не выше кишечнополостных и вполне вероятно близки к кораллам.
Очень интересен комплекс групп, помещенный за петалонамами. Наиболее крупная из них - тип археоциат, установленный А. Г. Вологдиным в конце 30-х годов и с тех пор достаточно прочно сохраняющий свой ранг, что довольно редко для "загадочных" групп. Достаточно хорошо разработана систематика этого типа, установлены классы, отряды и значительное число систематических единиц более низкого ранга. Выявлены гомологические ряды развития отдельных таксонов, значительно облегчающие работу систематиков (см. 4.1). И тем не менее археоциаты остаются загадкой. До настоящего времени не решен вопрос о принадлежности их к многоклеточным или одноклеточным, к животным или растениям. Не более достоверны знания и о других перечисленных группах, близких к археоциатам. Даже право на существование некоторых из них подвергается сомнению.
Поучительна "судьба" крибрициат. Первоначально они были известны в качестве "личинок" археоциат. Позже А. Г. Вологдин [181, 182] выделил эти организмы в самостоятельный класс (крибрициаты), а некоторые исследователи считали их даже типом. Однако в последние годы крупные специалисты по древнейшим организмам высказали мысль, что крибрициаты - не самостоятельные организмы, а лишь одна из форм существования археоциат: своеобразные выросты на кубках, получившие название "крибры" [374].
"Загадочность" всех названных групп, сходство между разными группами, как бы далеки они ни казались по внешней форме (в частности, сходство микроструктуры скелета), и явное несходство со всеми остальными организмами были причиной объединения их всех в особое царство археата [373]. С этим предложением согласны не все специалисты. Существует мнение, что рецептакулитов, археоциат и сходные с ними группы (афросальпингоид и соанит) следует сближать с зелеными водорослями, так как ультраструктура вещества археоциат сравнима с ультраструктурой водорослей. Однако и с этим согласны далеко не все исследователи. По их мнению, на ранних стадиях развития у наиболее примитивных представителей органического мира ультраструктуры скелетных образований могли быть достаточно сходными.
Достаточно "загадочны" еще две очень древние группы: Angustiochreida (отряд; верхи докембрия, низы кембрия) и Hydroconozoa (класс; кембрий возможно существовал и далее - до конца мезозоя). Первая группа объединяет очень странные организмы с трубчатым скелетом, часто с трехлучевой симметрией (рис. 52). Первоначально данная группа была выделена в качестве отряда неизвестной систематической принадлежности. В последнее время высказывается мнение, что это или самостоятельный класс кишечнополостных, или, возможно, особый подкласс в классе сцифомедуз [135]. Все же эта группа остается пока полной загадкой, так как трехлучевая симметрия, действительно хорошо выраженная у многих относящихся к ней форм, нехарактерна для кишечнополостных. Нет единого мнения и о втором из названных таксонов. Гидроконозои, первоначально описанные из кембрия [508], достаточно сильно отличаются от форм из мела - палеоцена, отнесенных к ним позже тем же автором. Не исключено, что это только сходные формы и класс нуждается в серьезной ревизии. Исключительно интересно некоторое сходство его представителей с современными стауромедузами. Трудно сказать, говорит ли это о родстве или о сходных путях эволюции в разных группах.
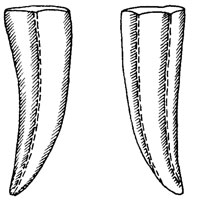
Рис. 52. Angustiochrea lata Valk et Sysoiev. По А. К. Валькову, В. А. Сысоеву. Северо-восток Сибирской платформы Ув. 20. р. Малая Куонамка, кембрий, аядакский ярус, вид раковины с двух сторон, видны борозды
Совершенно загадочны пока очень своеобразные кембрийские организмы, выделенные в особый класс Stenothecoida [2509]. Наличие двустворчатой раковины внешне роднит их с моллюсками, поэтому (и что самое интересное - почти одновременно) класс был выделен также И. А. Аксариной [10] под именем Probivalvia.
Однако, как выяснилось, эти створки являются не правой и левой, а брюшной и спинной. Исключительно своеобразная скульптура на наружной стороне створок отражается в виде валиков на внутренней их поверхности (рис. 53). На внутренней поверхности створок установлены более или менее метамерно расположенные вогнутости, трактуемые как следы прикрепления мускулов и отпечатки петель кишечника. Вместе с тем остается и много неизвестного, в частности, неясен механизм смыкания и размыкания створок. Таким образом, в настоящее время нельзя говорить о близости стенотекоидей (пробивальвий) к той или другой определенной группе организмов, что и дало С. Н. Розову [879] основание для выделения их в особый тип.
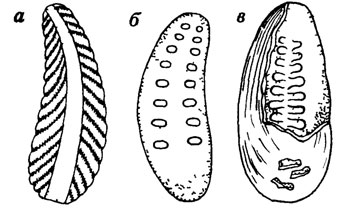
Рис. 53. Стенотекоидеи (сильно увеличено). По С. Н. Розову. а - внутреннее ядро вентральной створки Bagenoriella pectinata Aksar; б - внутреннее ядро дорсальной створки и мускульные отпечатки Stenothecoides kunatensis Aksar; в - внутреннее ядро дорсальной створки и отпечатки (?) кишечника Stenothecella sibirica Aksar
Достаточно загадочной группой являются хиолиты, получившие наибольшее развитие уже в кембрии, но продолжавшие существовать в течение всего палеозоя. Большинство исследователей считало хиолитов моллюсками, хотя твердой уверенности в этом не было, так как некоторые особенности хиолитов (в частности, своеобразные приустьевые придатки) совершенно неизвестны в других группах моллюсков. Недавно появились работы, в которых на основании детального анализа всех признаков хиолитов делается вывод о целесообразности выделения этой группы в самостоятельный тип [1026, 1336].
Иногда "загадочными" оказываются таксоны, основанные на фрагментарных остатках. В качестве примера можно привести класс Mattheva, установленный на основании анализа рода Matthevia Walcott из кембрия Северной Америки [2508]. От животного сохраняется только небольшая коническая раковинка, подразделенная продольной перегородкой на две довольно глубоких полости (рис. 54). Род Matthevia относили то к птераподам, то к ихнолитам, хотя он с трудом вписывается в рамки и тех и других. Известный специалист по древнейшим организмам Э. Йохельсон высказал предположение, что раковина является только одной из частей скелета, состоящего из двух таких раковинок (передней и задней), а, возможно, еще и ряда вставленных между ними по перечных пластинок. При такой реконструкции получается организм, несколько напоминающий моллюска из класса лорикат (рис. 55). Этим-то гипотетическим животным и было дано название Mattheva. Вне всякого сомнения, подобные реконструкции очень интересны, но все же класс остается пока гипотетическим и только дальнейшие находки принадлежащих к нему окаменелостей могут подтвердить его реальность или показать возможность иного решения проблемы.
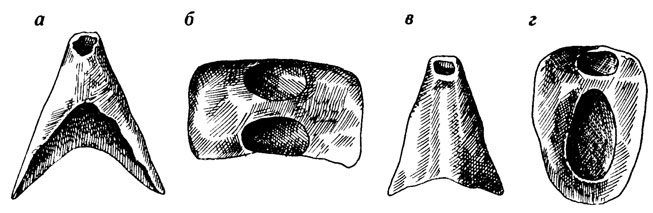
Рис. 54. Раковина Matthevia variabilis Walcott, передний конец обломан Ув. 1,5. По Э. Л. Иохельсону. а, б - вентральный и внутренний вид передней части, в, г - дорсальный и внутренний вид задней части
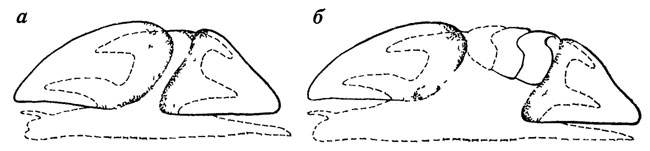
Рис. 55. Возможная реконструкция Matthevia. По Э. Л. Иохельсону. а - предпочтительная ориентация твердых частей, б - та же ориентация, но с гипотетически вставными промежуточными пластинами
Известны случаи, когда группа, казалось бы, совершенно точно нашедшая свое место в системе органического мира, вновь оказывается в числе "загадочных". Одним из наиболее ярких примеров такого рода является Volborthella, долгие годы считавшаяся одним из наиболее древних головоногих моллюсков, возможно, даже предком всех головоногих (правда, в последние десятилетия отряд Volborthellida считался только боковой своеобразной ветвью головоногих, но все же твердо оставался в этом классе). Исследователей всегда несколько затрудняла сохранность фольбортелл, от которых сохранялись только крошечные ядра. Детальные исследования последних лет показали, что это животное, видимо, вообще нельзя относить к моллюскам [874]. Оказалось, что так называемые "камеры" являются слоями, сложенными одной из форм кремнезема, "перегородки" - линиями соприкосновения этих слоев, а "сифон" - внутренней полостью, в которой, вероятно, обитало животное, строившее вокруг себя столь своеобразный и мощный (по сравнению с диаметром "сифона") защитный чехол. Природа и систематическое положение этого существа загадочны.
Известен еще ряд групп, которые остаются предметом дискуссий, хотя никто не сомневается в их реальности и достаточно высоком систематическом ранге: Stromatoporata (подкласс в одном из классов кишечнополостных, губки, самостоятельный класс беспозвоночных, водоросли), Chaetetida (коралловые, полипы, гидроиды, губки), Tentaculita (класс моллюсков, подкласс головоногих моллюсков); менее известны Machaeridia (черви), Xenoconchia (самостоятельный класс моллюсков, подкласс соленоконхов, гастроподы, хиолиты), Rostroconcha (самостоятельный класс моллюсков, отряд двустворчатых), Calcichordata (одна из групп примитивных иглокожих, или особый подтип, предковый для низших хордат).
В некоторых случаях столь разная трактовка "загадочных" таксонов не имеет существенного значения для понимания общего хода эволюции органического мира, в других - имеет. В качестве примера можно указать ростроконхов, которые, по мнению одних авторов, являются самостоятельным классом моллюсков с двустворчатой раковиной, предковым для ряда других классов, а с точки зрения других - только отрядом двустворок, предком иных больших групп не являющимся [1009]. Причина этого - совершенно различное понимание ориентировки раковины (передний конец, по мнению одних, является задним с точки зрения других). Вполне понятно, что такая переориентировка ведет и к совершенно иному пониманию остальных элементов скелета, а следовательно, и тела животного. Проблема ориентировки, безусловно, одна из важных. У кальцихордат, например, причина разного понимания их систематического положения тоже состоит в определении истинной ориентировки верхнего и нижнего концов скелетного остатка.
Совершенно особая категория - таксоны, включающие не целые организмы, а заведомо какие-то их части или стадии развития. Из наиболее известных назовем палеозойских хитинозой и кальцисферулид, достаточно хорошо известных из юрских и меловых отложений.
Перечислить все "загадочные" группы сложно. Во-первых, число их непрерывно увеличивается по мере накопления наших знаний, во-вторых, оно зависит от изменения наших взглядов: одна и та же группа то "обретает" свое место в системе, то вновь "теряет" его. Во всяком случае, "загадочных" групп, даже начиная с "семейства" и кончая "классом", несколько десятков [844, 1168, 1582]. Число же "загадочных" родов трудно поддается учету.
Хотя "проблематика" и "загадочные" группы известны из отложений разного возраста, подавляющее большинство их относится к докембрию и кембрию. Предполагают, что большинство докембрийских форм - совершенно особые ветви органического мира, исчезнувшие на рубеже криптозоя и фанерозоя, столь они своеобразны [1097]. Сложнее решить вопрос о кембрийских и более поздних группах неясного систематического положения. Совершенно очевидно, что в истории органического мира не могло быть форм "вне системы", не связанных родством с теми или иными достаточно хорошо известными группами. Любая группа, как бы мала она ни была, должна найти свое место. Выяснение этого - одна из важнейших задач исследователя, так как, во-первых, это помогает нам понять эволюцию органического мира в целом и выявить закономерности этого процесса, во-вторых, помогает точно выяснить значение "загадочных" групп, а иногда и проблематики для геологической практики.
При решении вопроса о систематическом положении таких групп теоретически возможны два решения: 1) группы являются частью других, хорошо известных таксонов, хотя и сильно уклоняющимися их ветвями; 2) группы принадлежат к самостоятельным высшим таксонам, как бы малы они ни были по объему. К выбору следует подходить осторожно. Присоединяя исследуемые группы к хорошо известным таксонам, приходится в большей или меньшей степени изменять характеристики последних, делая их более расплывчатыми. Кроме того, "слияние" небольших групп с другими неизбежно поведет к опасности "обеднения" истории эволюции органического мира, к представлению о том, что почти изначально появились все те высшие таксоны, которые продолжали существовать далее сотни миллионов лет. Тем самым мы как бы снимаем возможность "проб и ошибок" в развитии органического мира. Выделяя же "загадочные группы" в самостоятельные классы и типы, мы рискуем потерять значение высокого ранга таксонов как очень крупных ветвей, эволюция которых отражала специфику биосферы на разных этапах ее существования.
При изучении проблематик и "загадочных групп", видимо, не может быть единого подхода, так как слишком различны материалы, с которыми приходится иметь дело исследователю [995].
Для "настоящих" проблематик необходимо в первую очередь попытаться решить вопрос о их природе - являются ли они неорганическим образованием, остатком организма, отпечатком какой-то части организма или результатом его жизнедеятельности. Вероятно, одно из первых мест в этом анализе должно занимать выявление закономерности или хаотичности строения остатка, наличие определенных элементов, которые можно было бы считать какими-то особенностями скелета, пусть даже неясного назначения, выяснение микро- и ультраструктуры скелета, а также его химического состава. Состав скелета, сколь это ни странно, приходится ставить на последнее место, так как известны самые разные случаи его замещения.
Для "загадочных" групп, в разряд которых может перейти и проблематика в случае успешной ее расшифровки в качестве остатков скелета, также важно решить целый ряд вопросов. В первую очередь приходится думать над тем, являются ли изучаемые остатки скелетом взрослого организма или только какой-то стадии его развития. В качестве примера можно указать Chitinozoa, которые, возможно, являются цистами каких-то одноклеточных организмов. Не решен вопрос о природе кальцисферулид и некоторых других групп. Иногда сразу очевидно, что исследователь имеет дело только с частью скелета организма. Таков, например, риихолит. В подобных случаях следует стремиться к поискам сходных форм в мире современных организмов. Только наличие ринхолитов у современного наутилуса позволило безошибочно от нести все группы ископаемых ринхолитов к головоногим моллюскам. Без такого сопоставления они долгое время продолжали бы оставаться загадочными остатками, так как совместные находки ринхолитов с их "хозяевами" очень редки и сделаны только в последние годы. В данном случае помогло сравнение не с наиболее обычными и распространенными группами современных головоногих моллюсков, а с наиболее древней и, видимо, примитивной группой - наутилусом.
Вероятно, при сравнениях всегда следует обращать внимание не только на доминирующие в данном типе или классе формы, но и на реликтовые формы, наиболее уклоняющиеся группы и т. п. В ряде случаев полезные сведения можно получить также при детальном изучении современных тератологических форм. У них могут возникать особенности, утерянные в процессе эволюции подавляющим большинством особей данного таксона.
Совместные усилия неонтологов и палеонтологов очень важны для решения вопроса о принадлежности многих "загадочных групп". Так, в последние годы стали известны современные организмы, сходные по строению скелета с строматопорами, хететидами и даже с такой "загадочной группой" как сфинктозоа. По-видимому, нельзя утверждать, что вопрос о систематической принадлежности названных групп ископаемых решен окончательно, так как не исключена возможность, что они относятся к полностью вымершим группам. Известно, что явление сходного развития отдельных элементов скелета у разных групп организмов в истории органического мира очень распространено, но даже наличие аналогий значительно облегчает задачу палеонтологов.
При изучении "загадочных" форм, представленных более или менее полными скелетами, на первое место выходит выяснение их плана строения. По В. Н. Беклемишеву [61], план строения характерен для типа. Поэтому любой анализ следует начинать именно с этого вопроса, хотя, конечно, он далеко не всегда приведет к окончательному ответу о принадлежности исследуемого организма к определенному типу. Следует выяснить тип симметрии, монолитность или членистость скелета, расположение частей скелета и их предположительное значение, а также микро- и ультраструктуру скелета. Ультраструктуре в последнее время уделяется особое внимание. По ультраструктуре скелета далеко не всегда можно установить тип организма, так как данных накоплено мало. Однако в некоторых случаях, например, при решении вопроса - принадлежит остаток "червям" или моллюскам, этот метод дает совершенно определенное заключение.
Следующим шагом по пути выяснения систематического положения изучаемых остатков должны быть поиски аналогий среди современных и вымерших групп. Так, двустворчатая раковина характерна для бивальвий (пелеципод) и брахиопод, но встречается также у гастропод и некоторых ракообразных. Раковина с поперечными перегородками характерна для головоногих (хотя в этом же классе значительное число форм имеют сильно редуцированную раковину или ее лишены), но перегородки имеются также у тентакулитов, хиолитов, моноплакофор и гастропод. Необходим дальнейший морфофункциональный анализ, позволяющий определить истинную природу сходных образований и выяснить возможную принадлежность изучаемого организма к той или иной группе.
В качестве примера достаточно сложного анализа остановимся на истории выяснения систематического положения хиолитов [1026]. В течение долгого времени хиолиты рассматривались в составе типа моллюсков. При этом вполне естественно коническая раковина сопоставлялась с раковиной гастропод, а крышечка - с крышечкой гастропод. Предполагалось наличие хорошо развитой головы, ноги и мантийной полости, расположенной над ногой. Однако крышечка брюхоногих имеет спиральное строение, у хиолитов же - двустороннесимметричное. На крышечке хиолитов имеется серия различных образований, что говорит о довольно сложной мускулатуре, прикреплявшейся к крышечке, и о том, что крышечка выполняла несколько функций, а не только прикрывала устье раковины. Расположение предполагаемых мускулов заставляет сомневаться в возможности существования ноги такого типа, как у брюхоногих моллюсков; скорее всего никакого образования, гомологичного ноге моллюсков, у хиолитов вообще не было. Подобное расположение мускулов также затрудняло функционирование жабр в мантийной полости, располагавшейся (при условии принятия моллюскоидной модели хиолитов) над предполагаемой головой на топографически спинной стороне. Не остается места и для желудка. Рассматривается также попытка сравнения мускульных отпечатков хиолитов и головоногих моллюсков и моноплакофор. Оказывается, что одни мускулы соединяют раковину и крышечку, а другие - раковину и мягкое тело. Как видно, возможное строение хиолитов трудно совместимо со строением моллюсков (не только головоногих, но и других). В работе [1026] говорится также о попытке сравнения хиолитов с сипункулидами, но при тщательном анализе и в этом случае выявляется ряд отличительных признаков хиолитов, не позволяющих сближать их с сипункулидами. Таким образом, методом сравнения с разными известными группами исследователи приходят к необходимости создания своей модели для хиолитов и выделения их в особый тип животного мира.
Наконец, необходимо обратить внимание на важность максимально тщательного сбора различных проблематик и всяких уклоняющихся (аномальных) экземпляров, принадлежащих даже к уже известным загадочным группам. Иногда такие находки заставляют пересмотреть уже сложившееся мнение о природе тех или иных групп. Так произошло, в частности, с крибрициатами, когда были найдены их экземпляры, непосредственно "сращенные" с типичными археоциатами. Так же была установлена принадлежность ринхолитов не только наутилоидеям, но и аммоноидеям и т. д. Однако и такие находки требуют самого тщательного анализа. В частности, не до конца ясен вопрос с крибрициатами - не исключено, что они и археоциаты все же разные организмы, но по каким-то причинам иногда сращенные вместе. Выяснение вопроса о принадлежности ринхолитов мезозойским аммоноидеям и наутилоидеям ставит вопрос о необходимости тщательного поиска подобных образований в палеозое, где аммоноидеи широко распространены. Отдельные указания на находки палеозойских ринхолитов имеются, но все это требует дальнейших подтверждений. Если же окажется, что у палеозойских наружнораковинных ринхолитов не было - возникнет новый вопрос о причинах возникновения такого образования в мезозойских формах, причем нужно помнить, что внутреннераковинные прекрасно обходятся без ринхолитов.
Важность исключительно внимательного анализа различных "непонятных" остатков, находимых палеонтологами и геологами при полевых работах, а также необходимость периодического пересмотра "проблематик" в старых коллекциях особенно хорошо видна из истории изучения конодонтов. Эти достаточно широко распространенные ископаемые остатки, имеющие большое значение для стратиграфии, давно являлись предметом оживленных дискуссий. Всем было ясно, что находимые остатки представляют собой часть организма, но какого именно - оставалось неясным. Их рассматривали в качестве челюстей червей, элементов скелета рыбообразных представителей позвоночных, относили и к разным другим группам.
Исключительно счастливая находка образца с конодонтоносителем, хранившегося с 1925 г. в музее Геологического института в Эдинбурге, положила конец основной части дискуссии [26, 1379]. "Носителем" конодонтов оказалось небольшое животное, несколько напоминающее представителей из типа щетинкочелюстных (хетогнат, см. рис. 43, А). Важно, что тип Conodonta - название было предложено ранее для гипотетических обладателей конодонтов - теперь имеет свою характеристику (см. 1.4).
Можно смело сказать, что работа с "загадочными" группами - одна из наиболее увлекательных сторон палеонтологических исследований.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'