
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
2.3. Структура (палеогистология) скелета беспозвоночных животных (И. С. Барсков, В. П. Макридин)
Основная область структурных исследований в палеонтологии беспозвоночных организмов - изучение ископаемых скелетных остатков на уровнях органа (макроструктура) и ткани (микроструктура).
К настоящему времени наиболее изучена структура скелетных образований моллюсков и брахиопод, и именно на этих группах удобнее всего рассмотреть некоторые общие и частные проблемы и направления данных исследований [609]. В меньшей степени, часто лишь в виде литературных ссылок, будет рассмотрена структура других групп организмов, обладающих минерализованным скелетом (фораминифер, кишечнополостных, мшанок, иглокожих). В сферу структурных исследований входит изучение и неминерализованных скелетных образований у таких групп, как граптолиты и полихеты (сколекодонты). О них приведены здесь лишь общие сведения с ссылками на основные литературные источники.
Первые микроскопические исследования скелетной ткани (раковин) моллюсков были проведены в середине прошлого века [1408]. Важные сведения об особенностях строения перламутра были получены в 20-е годы нашего столетия [2228]. Основополагающую роль сыграла работа О. Б. Беггильда [1335], в которой на большом материале раковин современных и ископаемых моллюсков разных классов показано разнообразие их микроскопического строения, систематизированы микроструктурные типы, понимание которых в большей части сохранилось до настоящего времени.
Большим шагом вперед стали в конце 50-х и в 60-е годы исследования японских ученых, связанные с выращиванием "искусственного" жемчуга. С них взяло начало применение при исследовании структуры скелета электронной микроскопии и биохимических методов. Правда, эти работы были ограничены изучением небольшого числа групп двустворчатых моллюсков. Новый толчок к расширению микроструктурных исследований дало развитие в середине 60-х годов техники растровой электронной микроскопии. В результате изучения обширных коллекций Британского музея [2364, 2365] и музеев Японии [1838] были охарактеризованы микроструктурно более 100 (из 120-140 известных в настоящее время) семейств современных двустворчатых моллюсков. Проводились также работы по другим группам моллюсков, работы сравнительного направления, изучение онтогенеза структур и детальные исследования в пределах таксонов низкого ранга.
В результате проведенных исследований установлено, что моллюски обладают максимальным среди беспозвоночных разнообразием микроструктурных типов, слагающих скелетные элементы. Ниже приводятся типы микроструктур и текстуры скелета моллюсков [51]:
| Типы микроструктуры | Текстуры |
|---|---|
| Призматический | Сферулито-призматическая, призматическая, полупризматическая, субконическая, радиально-призматическая, фурциллятная |
| Сложный призматический, фиброидно-призматический, транспризматический, составной призматический, фиброидный, перекрещенно-пластинчатый | |
| Сложный перекрещенно-пластинчатый | Конусная, блоковая, спутанно-пластинчатая, правильная |
| Листоватый | Неправильная, перистая, субромбическая |
| Перламутровый | Кирпичная, столбчатая, ступенчатая, планочная |
| Гомогенный, мозаикостракум |
Для двустворчатых моллюсков разработана следующая классификация микроструктурных групп и их основных разновидностей [1411]:
Микроструктурные группы и их основные разновидности: I. Призматическая: А. Простая призматическая. Б. Фиброидная призматическая. В. Сферулитовая призматическая. Г. Сложная призматическая. II. Сферулитовая. III. Пластинчатая: А. Перламутровая. Б. Параллельно-листоватая. IV. Перекрещенная: А. Перекрещенно-пластинчатая. Б. Перекрещенно-игольчатая. В. Сложная перекрещенно-пластинчатая. Г. Перекрещенно-спутанная (линейная). V. Гомогенная: А. Гомогенная. Б. Зернистая (гранулярная). VI. Изолированные спикулы или остроконечники. VII. Изолированные кристаллические морфотипы. Микроструктурные типы и их образующие категории: I. Арагонитовая призматическая. II. Кальцитовая призматическая. III. Перламутровая (арагонитовая). IV. Фарфоровидная (арагонитовая): А. Арагонитовая перекрещенно-пластинчатая. Б. Арагонитовая перекрещенно-игольчатая. В. Арагонитовая сложная перекрещенно-пластинчатая. Г. Арагонитовая перекрещенно-спутанная (линейная). Д. Арагонитовая гомогенная. V. Листоватая (кальцитовая): А. Параллельно-листоватая. Б. Кальцитовая перекрещенно-пластинчатая (перекрещенно-листоватая). В. Кальцитовая сложная перекрещенно-пластинчатая (сложная перекрещенно-листоватая). Микроструктуры, редко встречающиеся в раковинах двустворчатых моллюсков: I. Сферулитовая. II. Изолированные спикулы или остроконечники. III. Изолированные кристаллические морфотипы.
Существуют разные личные подходы к классифицированию структур, хотя понимание главнейших из них более или менее однозначно и в целом соответствует исходным представлениям О. Б. Беггильда. В классификациях достаточно полно отражено разнообразие структур, но отсутствуют генетическая основа и единая методика их описания, отражающие общность происхождения структур в качестве продукта эпителиальной секреции.
Различия между крупными ветвями моллюсков выражаются в распространенности и в специфичности сочетания определенных микроструктурных типов в скелетных образованиях. Так, у цефалопод и моноплакофор присутствуют скелетные ткани лишь с перламутровым и призматическим типами микроструктуры. У скафопод известны перекрещенно-пластинчатый тип с его различными вариантами и призматический тип. Для двустворчатых моллюсков характерно большое разнообразие микроструктурных типов и вариантов их сочетаний (рис. 58-61). Одной из особенностей является отсутствие в одной раковине сочетания перламутровой и перекрещенно-пластинчатой структур. Почти так же разнообразны микроструктуры раковин гастропод. Интересно, что у некоторых трохид в одной раковине присутствуют слои и перламутровой, и перекрещенно-пластинчатой микроструктуры.

Рис. 58

Рис. 59. Перекрещенно-пластинчатая структура наружного слоя раковины двустворчатого моллюска Irus irus L. Ув. 3000. Керченский п-ов, чокрак. Палеонтологический институт АН СССР, кол. С. В. Попова

Рис. 60

Рис. 61. Составная призматическая структура наружного слоя раковины двустворчатого моллюска. Сев. Молдавии. Палеонтологический институт АН СССР, кол. С. В. Попова
"Наложение" данных по структуре раковины двустворчатых моллюсков на филогенетические схемы, построенные по конхологическим и сравнительно-анатомическим признакам, позволило составить представление об общих тенденциях в эволюции структур в этом классе. Исходным типом строения признается трехслойная раковина со средним перламутровым, наружным и внутренним простыми призматическими слоями [2363]. В историческом развитии происходит отчетливая смена таксонов с этим типом микроструктуры и группами, обладающими перекрещенно-пластинчатыми структурами, которые достоверно известны с мезозоя. У гетеродонтных двустворчатых моллюсков, преобладающих в мезозое и кайнозое, перекрещенно-пластинчатые структуры развиты наиболее полно. В этой группе известны многочисленные варианты (текстурные типы) перекрещенно-пластинчатых структур и прослеживаются их взаимные переходы. Эти переходы - появление у наиболее молодых и наиболее продвинутых представителей гомогенной (или гранулярной) микроструктуры - отражают, вероятно, некоторые общие тенденции в эволюции биоминерализации. Наиболее четко они демонстрируются в следующем ряду структур: перекрещенно-пластинчатая → сложная перекрещенно-пластинчатая → гомогенная.
На первом этапе элиминируется упорядоченность в расположении пластин первого порядка и исчезают разделяющие их матрицы. На втором этапе то же самое происходит с пластинами второго порядка, так что образующиеся неопределенной и сложной формы блоки часто изометричные, но не пластинчатые, составлены "пластинами" третьего порядка, точнее, тонкими "прутьями", имеющими диаметр около 2-5 мкм. Дальнейшее "распадение" этих кристаллитов на изометричные зерна (гранулы) размером 2-5 мкм со случайной ориентировкой оптических осей лишает структуру строгой упорядоченности. Если при становлении биоминерализации осуществлялся, по-видимому, процесс организованности структуры с появлением нескольких иерархических уровней, то здесь наблюдается обратное явление: уменьшается число органических матриц, структура становится однородной, сокращается количество иерархических уровней и снижается содержание органических компонентов.
Важным достижением микроструктурных исследовании явилось становление теории секреторных фаз в деятельности млн тинного эпителия у моллюсков [1991] пли аналогичной теории "стандартного режима секреции" у брахиопод [2493]. Основу этой теории составляет положение, что на постларвальной стадии онтогенеза эпителий мантии имеет определенную зону роста, локализованную на внутреннем краю ее наружной лопасти вблизи наружной мантийной борозды. В последние годы, однако, появились данные об отсутствии у мантийного эпителия локализованной зоны роста секретирующего ("раковинного") поля, что требует внесения корректив в концепцию секреторных фаз. Тем не менее ее приложения, наиболее последовательно развитые применительно к объяснению эволюционных изменений структуры в скелетных образованиях цефалопод, весьма успешны.
Исходя из теории последовательных секреторных фаз проведена гомологизация трех слоев стенки раковины со слоями перегородок фрагмокона у современного наутилуса, показано соответствие двух слоев соединительных колец сифона наружному (сферулито-призматическому) и среднему (перламутровому) слоям стенки раковины [1991]. В то же время оболочка сифона у аммоноидей, состоящая только из одного слоя, который соответствует лишь перламутровому слою кольца наутилуса [328, 344], не гомологична сифонной оболочке наутилуса. Развитие этих исследований в филогенетическом плане для других крупных групп цефалопод выявило существенное разнообразие в структуре оболочки сифона и позволило проследить различные пути их развития по этому структурному признаку [50].
Та же теория была положена в основу сопоставления скелета внутреннераковинных цефалопод (белемнитид и сепии) и его сравнения со скелетом предковых групп наружнораковинных [47, 49]. Сравнительное структурное изучение привело к возникновению понятия о гомоструктурности слоев и элементов скелета, потребовало конкретизации представлений о их гомологизации. Одинаковые и выполняющие одну и ту же функцию (соответственно топографически гомологичные и гомодинамные) части скелета могут существенно различаться по структуре и быть образованы слоями, сформированными в разные физиологические фазы секреции. Так, оболочки сифона у эндоцерид, аммоноидей и наутилид не гомологичны друг другу. Рассматриваемый тип гомологии было предложено называть физиологической гомологией [48]. Применение принципа физиологической гомологии в структурном изучении скелетных элементов и других беспозвоночных весьма перспективно.
Важное направление микроструктурных исследований у моллюсков - изучение закономерностей онтогенетических преобразований структур. На этом пути получен ряд важных сведений, позволяющих судить о наличии или отсутствии личиночной стадии, о времени и характере перестроек в онтогенезе секреторной функции мантийного эпителия. Работы выполнены на современных и ископаемых гастроподах [1300, 1781] и цефалоподах [330, 344, 404, 1854]. Хотя интерпретации хода онтогенеза у ископаемых форм, проводимые разными исследователями, не всегда одинаковы, они наполняют новым содержанием представления, основанные только на морфологическом изучении.
Существенное развитие в последние годы получили исследования скелетного вещества беспозвоночных на ультраструктурном уровне, предусматривающем изучение пространственных и размерных соотношений мельчайших кристаллических элементов, из которых состоят кажущиеся оптически однородными "элементарные" монокристаллы микроструктурного уровня. Исследования Х. Мутвея [718, 1995] показали, что элементарные компоненты микроструктуры перламутрового слоя - полигональные таблетки - могут иметь весьма различную ультраструктуру. Значительную роль во взаимной ориентировке ультраструктурных элементов играют процессы двойникования (см. 2.1).
Одно из перспективных направлений - изучение структуры скелета вымерших групп моллюсков и групп, сближаемых с этим типом. Если современные двустворчатые моллюски в ранге семейств изучены примерно на 90%, то среди 170 семейств ископаемых микроструктурными исследованиями охвачено не более 20%. Так что даже простые "иконографические" материалы по ископаемым моллюскам будут иметь важное значение.
Полноценное микроструктурное изучение древнейших (кембрийских и ордовикских) представителей типа моллюсков несет и общебиологические задачи. Практически одновременное появление в скелетной форме представителей различных классов моллюсков поможет, вероятно, выявить структурные и биохимические закономерности становления и ранних этапов развития процесса биоминерализации как важнейшего феномена в эволюции организмов, в частности, таких, как возрастание роли минеральной компоненты в скелетных образованиях, переход от одноклеточного типа секреции к тканевому, выявление примитивных и продвинутых типов микроструктур и т. д.
Важной и малоизученной стороной микроструктурных исследований ископаемых является изучение тех процессов, которые протекают при фоссилизации скелетных остатков, приводящих к развитию вторичных структур. Эти процессы, включающие диагенез, перекристаллизацию и другие явления, сопровождающие долгую "жизнь" скелетных остатков, применительно к изменению структур скелетной ткани могут быть названы общим термином "палеоизация" предложенным М. Флоркеном для обозначения изменений органической составляющей скелетов (см. 2.6). Это направление является одной из сторон тафономических исследований, тафономией на микроструктурном уровне. В настоящее время можно назвать лишь немного работ, в которых рассматриваются эти вопросы [56]. Здесь обращают на себя внимание многолетние экспериментальные исследования Ш. Грегуар по влиянию "диагенетических факторов" - пиролиз, воздействие воды и воздуха в различных условиях на сохранность структуры и состава органической и минеральной компоненты перламутрового слоя [1669].
Бытовавшие ранее представления о процессах перехода скелетных остатков в ископаемое состояние ("выгнивание" органического вещества скелета, заполнение образовавшихся пустот новыми минералами и т. д.) не соответствуют действительности. Новые подходы к выявлению закономерностей палеоизации [253] открывают представления о роли органических матриц скелета и твердофазных процессов, которые часто могут происходить еще при жизни организмов (см. 2 1).
Сходные проблемы стоят и решаются при микроструктурных исследованиях брахиопод - другой крупной группы "раковинных" беспозвоночных.
Раковины замковых брахиопод характеризуются исключительно кальцитовым, а не арагонитовым минеральным составом, и существенно меньшим набором микроструктурных типов, даже в сравнении с кальцитовыми раковинами моллюсков. В этом проявляется, несомненно, специфика биоминерализации у этих групп, что естественно, так как появление скелета у моллюсков и брахиопод происходило независимо и нельзя говорить о наследовании физиологического режима секреции. Вместе с тем, как показано [1411], у представителей обоих типов имеются одинаковые типы структур - гомоструктурность [1411]. Это свидетельствует о существовании общих закономерностей биоминерализации, что позволяет в принципе очертить возможные, реализуемые и запрещенные типы структурного проявления процессов. Для этого необходима выработка единой классификации типов микроструктур, основанной на общей теории биоминерализации, которая объясняла бы с единых позиций образование биоминералов и их морфологию от простейших до позвоночных.
Основополагающие исследования структуры раковин брахиопод проведены А. Вильямсом [2493], который ввел понятия секреторного режима в деятельности мантии, предложил первые схемы филогенеза наиболее распространенных групп на основе сходства структур и физиологическую интерпретацию их эволюционных изменений А. Вильямс выделил группы со "стандартным" режимом секреции и с отклоняющимся режимом, обосновал понимание фиброзного слоя как продукта одноклеточной секреции, среди форм со "стандартным" режимом секреции выделил группы с трехслойной и двуслойной раковиной.
Микроструктурные исследования большого числа представителей, в частности крупнейших вымерших таксонов замковых и беззамковых брахиопод с карбонатным скелетом, увеличили разнообразие микроструктурных типов в раковинных слоях, поставили ряд интересных проблем, касающихся методических вопросов и эволюции структур.
Большинство таксонов замковых брахиопод обладает относительно однотипной структурой раковины, в строении которой основную конструктивную роль играет фиброзный, так называемый "вторичный", слой (рис. 62). Вместе с тем среди групп со стандартным режимом секреции у значительного числа таксонов различного ранга обнаружена значительная гетерогенность в строении фиброзного слоя. Внутри слоя установлены участки, построенные фибрами различной формы и пространственной ориентировки, выделяемые в качестве подслоев [466, 834, 1072 и др.]. Остается до юнца невыясненной природа присутствующего у многих форм слоя, облекающего изнутри стенку раковины и внутренние морфологические элементы [каллотеста, по 2425]. Согласно В. П. Камышану [466], каллотест гомоструктурен вторичному фиброзному слою и мог бы считаться его модификацией, но локализация в раковине позволяет рассматривать его в качестве самостоятельного слоя. Тесно связано с этим вопросом и выяснение особенностей структуры элементов замочного и ручного аппарата. По микроструктурному типу (фиброзной структуре) они принадлежат к той же секретной фазе, в течение которой образуется и "вторичный" слой стенки раковины. Однако слои, образующие элементы внутреннего строения раковины, по ряду признаков заметно отличаются от слоев ее стенки. При этом разные элементы (круры, наружные замочные пластины, срединная дорсальная септа, а также дельтидиальные пластины и замочный отросток) могут иметь существенные структурные отличия. В их строении участвуют слои нескольких генераций [466], при этом некоторые из слоев (например, медиотест) выделяются как самостоятельные структурные элементы. Образование этих слоев связано, вероятно, с наличием отдельных генеративных зон в локальных складках эпителия, которые формируют внутренние скелетные элементы.
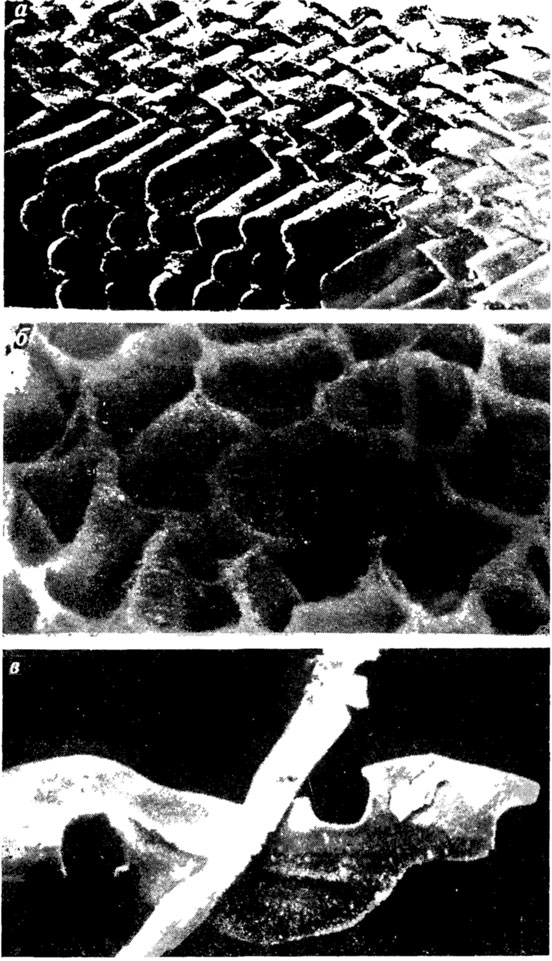
Рис. 62. Фиброзный слой раковин замковых брахиопод
По данным Т. Н. Грунт [277], у атиридид внутренние элементы скелета достаточно разнообразны в составе замочных пластин и срединной септы могут присутствовать слои разной микроструктуры, не только фиброзные, но и гомоструктурные "первичному" и, возможно, "третичному" слоям стенки раковины. Это, несомненно, служит подтверждением существования локальных зон роста связанных с элементами внутреннего строения раковин, что открывает возможности для использования при их сопоставлении у разных групп брахиопод представлений о физиологической гомологии, успешно применяемых к моллюскам.
Перспективны исследования раковинного вещества брахиопод и на ультраструктурном уровне. Так, у современных [715] и ископаемых [1222] ринхонеллид показана неоднородность строения фибр, которые считались до этого "сплошными" монокристаллами. В последние годы получены данные об отсутствии органического матрикса внутри фибр, которые представляются в виде закономерного сдвойникованного вдоль оси каждой фибры монокристалла кальцита (рис. 63).

Рис. 63. Микроструктура слоев раковины брахиоподы Terebratulina retusa. L., Баренцево море, современные. Материал О. Н. Морозовой. а - внутренняя поверхность раковины, фиброзный слон. ув. 2100; б - фибра со следами травления по плоскостям двойникования, расположенным косо к ее продольной оси, ув. 5700; в - структура первичного слоя, ув. 1000
Интересны и важны исследования специфических групп брахиопод, которые обладают "нестандартным" режимом секреции. Так, Т. Н. Смирновой прослежены закономерности изменений структуры раковин в филогенетически связанных родах тецидеидей [968] редукция слоев и перестройка их микроструктуры, изменения происходят независимо в брюшной и спинной створках. По меньшей мере в двух отношениях необычна структура раковин давидсониаций и строфоменаций [36] - по наличию слоев специфического перекрещенно-листового типа (crossed bladed) и по присутствию "псевдопористости", которая имеет разное структурное выражение у разных групп этих организмов. Интересны также новые данные о структуре фосфатных раковин беззамковых брахиопод [1782].
Изучение структур раковин брахиопод, среди которых много вымерших крупных групп, более всего перспективно в историческом аспекте. Получаемые данные имеют значение для выяснения филогенетических отношений внутри типа, для решения актуального вопроса о его таксономической однородности или неоднородности на уровне высших таксонов, а также для разработки более общей проблемы - познания особенностей и причин разнообразия структурного выражения процесса биоминерализации.
Характеристики микроструктуры скелета беспозвоночных становятся обязательной составной частью описаний таксонов различного ранга и филогенетических построений. Степень и уровень их использования в исследованиях разных групп различны, но интенсивно возрастают.
Данные о микроструктуре раковин фораминифер успешно используют в систематике. Изучение процесса обызвествления раковины в онтогенезе некоторых форм показало, в частности, что существенные различия на начальных стадиях минералообразования (нуклеация, первичный рост) у различных таксонов могут приводить в конечном итоге к образованию идентичных структур.
Весьма важны для построения общей системы саркодовых данные о вероятной филогенетической смене при образовании их раковины типов минерализации от "индуцируемой" к "матрицируемой" [54, 55, 1749].
Вполне вероятно, что особенности эволюции биоминерализации и смены ее типов удастся установить при изучении структуры скелета низших беспозвоночных (губок, археоциат, кишечнополостных), как и древнейших моллюсков, брахиопод, членистоногих и иглокожих. Интеграция процесса биоминерализации со скелетообразованием происходила, по-видимому, во многих группах параллельно и начиналась на стадии "индуцируемой" минерализации. Подтверждения этому содержатся в исследованиях структуры скелета археоциат и губок [54, 2186].
В развитии структуры раковин моллюсков и брахиопод наблюдается переход от одноклеточного (внеклеточного и внутриклеточного, спикульного) к тканевому типу секреции. При этом у моллюсков эволюционно перспективным оказался второй тип. Секреция раковины брахиопод, как полагают, происходит по первому типу, хотя в последние годы появились данные о тканевой секреции фиброзного слоя у брахиопод (см. рис. 62).
Изучение микроструктуры скелета кораллов также показывает сочетание различных типов биоминерализации [1799, 2304, 2305]. Особенно интересны в этом отношении исследования палеозойских кораллов, так как предполагается, что у них происходила смена кальцитовой биоминерализации на арагонитовую, что не наблюдается среди других организмов и наводит на мысль о независимом приобретении скелета мезозойскими склерактиниями.
Много проблем возникает и при изучении микроструктуры скелета мшанок, обладающих большим разнообразием ископаемых представителей. Известные в этой области работы [2179 и др.] рисуют сложную картину сочетания примитивных и специализированных микроструктур.
Для членистоногих характерна весьма сложная внутренняя организация скелетных образований при относительно простой микроструктуре их собственно минеральной фазы [1993, 2098, 2394, 2507]. Есть основания предполагать, что специальные исследования помогут выяснить различия в микроструктуре интегумента у морских и пресноводных форм, что может оказаться важным при изучении палеоэкологии ряда древних групп водных членистоногих.
Биоминерализация скелета иглокожих весьма специфична и резко отличается от биоминерализации других беспозвоночных сочетанием эпителиальной и мезодермальной секреции, тканевого и одноклеточного (спикульного) типов минерализации, сложностью внутренней организации (стереом) скелетных элементов и простотой их кристаллографической структуры, где каждый элемент независимо от сложности морфологии представляет собой оптический монокристалл [2006, 2030, 2183, 2392]. Приведенные особенности, а также большое разнообразие крупных таксонов ископаемых иглокожих делают исследования структуры скелета этих организмов весьма перспективными и важными.
Структурные исследования в палеонтологии включают изучение не только первичноминерализованных остатков скелета, но и кутикулярных остатков, не имевших или имевших прижизненно очень незначительное количество минеральной фазы, а также изучение строения органической фазы после его деминерализации.
Очень интересные результаты получены А. Урбанеком [2409, 2411, 2412], изучающим структуры скелета граптолитов на ультраструктурном уровне. А. Урбанек установил не только исключительно хорошую сохранность ультраструктурных деталей строения оболочки ордовикских, силурийских и девонских граптолитов, но также обосновал их биологическую принадлежность и таксономический статус, разработал на совершенно новой основе систему и проследил важнейшие закономерности эволюции этих организмов.
Следует упомянуть еще исследования по ультраструктуре челюстей полихет - сколекодонтов [1622], а также обратить внимание на перспективность изучения деминерализованных скелетных остатков, сохраняющих органические матрицы, коллагеновые нативные фибриллы с их характерной поперечной исчерченностью, специфичной для таксонов различного ранга и обнаруживаемой в ископаемом состоянии как в скелетах позвоночных, так и беспозвоночных, в том числе низших [1602, 1622].
Использование методики ультратонких срезов и изучение декальцифицированного скелетного материала с помощью электронных микроскопов высокого разрешения открывают все новые возможности в детальнейшем изучении тончайшего его строения.
Структурные (палеогистологические) исследования находятся на магистральном пути развития современной палеонтологии. Они стали уже неотъемлемой частью палеонтологических работ, и задача состоит не только в дальнейшем накоплении фактических данных, но и в их обобщении в целях дальнейшей разработки таксономических, филогенетических, палеоэкологических и других проблем.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'