
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
2.4. Палеогистология скелета позвоночных животных (В. Н. Талимаа, Э. И. Воробьева, В. П. Пегета, К. Е. Михайлов)
В ископаемом состоянии обычно сохраняются минерализованные скелетные остатки позвоночных. В их число, помимо костей, могут входить рога, волосы, кератиновые зубы, плавниковые лучи, скорлупа яиц и др. Случаи же фоссилизации мягких тканей крайне редки и в основном отмечаются у плейстоценовых животных. Иногда сообщается о подобных находках среди палеозойских форм - например, о сохранении покровов у позднепермского дейноцефала Estemmenosuchus uralensis [1144], но эти данные представляются спорными.
Первоочередной задачей палеонтологических исследований обычно является выяснение природы самих находок и определение их таксономической принадлежности. Однако по мере накопления таких сведений естествен переход к их классификации, филогенетическому осмыслению и использованию в построения палеонтологических рядов форм, т. е. форм, связанных геохронологической и морфологической последовательностью.
В течение последних 30 лет палеогистологические исследования позвоночных все чаще связывают с общей проблемой эволюционных преобразований тканей в животном мире. Эта проблема прямо влияет на решение вопроса о специфике эволюции тканевого уровня организации по отношению к органному и возможности создания естественной системы тканей на филогенетической основе.
Дискуссии в палеонтологии затрагивают широкий круг вопросов происхождения крупных таксонов позвоночных положение круглоротых и челюстноротых в системе, их связи с древними рыбообразными, пути эволюции хрящевых рыб, хрящевых ганоидов, двоякодышащих, кистеперых, низших тетрапод и т. д. Уровневый и филогенетический аспекты взаимосвязаны, поскольку от правильности построения филогенетических рядов позвоночных зависит во многом объективность выводов о характере перестроек тканей в эволюции и соответственно о специфике их исторического развития. Однако наличие конвергенции и параллелизмов нередко затушевывает истинное родство организмов.
Работы У. К. Вильямсона [2494, 2495] - одно из наиболее ранних и тщательных исследований костной ткани у современных и ископаемых рыб, в результате которого выделены несколько типов микроскопического строения их чешуи. Костная ткань костистых рыб изучалась также А. Келликером [1840, 1841, 1842], впервые описавшим у них бесклеточную костную ткань и полагавшим, что наличие или отсутствие остеоцитов можно использовать как важный признак в их систематике.
Первые шаги в решении проблемы тканевой эволюции были предприняты Э. Геккелем в 70-80-х годах прошлого столетия, когда он попытался распространить исторический подход на гистологию. Однако усилия Э. Геккеля не увенчались успехом прежде всего из-за недостаточной изученности тканей в разных типах животного мира и отсутствия необходимых сведений об особенностях их гистогенезов. Гистология долго развивалась преимущественно в области медицины, внимание ее было сосредоточено на изучении тканей человека и рецентных групп позвоночных. Естественно, что и классификация тканей позвоночных была первоначально разработана на рецентном материале. После неудачной попытки Э. Геккеля подвести филогенетическую основу под гистологию предпринимались лишь отдельные исследования тканей позвоночных в эволюционном аспекте [1630, 1656, 1732, 1835, 2176, 2334, 2342].
Двадцатые - шестидесятые годы нашего столетия - новый этап в изучении тканей позвоночных и прежде всего скелетных тканей, представляющих наибольший интерес с позиций палеонтологии.
В это время появляются фундаментальные труды Ф. Вайденрайха [2459-2463 и др.] и В. Гросса [1676, 1677], в которых приведены классификации "твердых" тканей позвоночных, основанные на особенностях гистогенезов. Гистология ископаемых бесчелюстных, рыб и тетрапод в связи с решением вопросов их филогении все чаще обсуждается в отечественной литературе [125, 127, 761, 1393-1397 и др.].
Особо следует выделить работу А. В. Румянцева [894], который с позиций теории филэмбриогенеза А. Н. Северцова [926] попытался создать целостную концепцию эволюции хрящевой и костной ткани на основе анализа накопившихся сведений по гистогенезу эндо- и экзоскелета у рецентных и ископаемых позвоночных. Эта книга, опубликованная спустя 11 лет после смерти автора, была написана, когда палеонтология еще не располагала достаточным палеогистологическим материалом. Тем не менее рассмотренные в ней вопросы эволюционной гистологии и общая картина гистогенезов не утратили актуальности и в наше время.
В опубликованной в середине нашего столетия сводке Т. Эрвига [2033] приведена новая, существенно более детальная, чем прежние, классификация скелетных тканей ископаемых позвоночных и методика их исследований, предпринята попытка объяснения особенностей гистогенезов дентиновых и костных тканей у бесчелюстных и рыб, а также описаны эндоскелетные твердые ткани нетелеостомных рыб и некоторых ганоидов. Этот труд положил начало многолетним и весьма плодотворным исследованиям в области эндо- и экзоскелетных тканей низших позвоночных.
Общее число публикаций по скелетным тканям (в основном ископаемым) в течение 20-60-х годов XX в. насчитывает более 140 наименований [2035]. Наряду с накоплением фактического материала в этот период были уточнены классификация, природа и место образования, выяснены основные закономерности минерализации твердых тканей, усовершенствована методика лабораторных гистологических исследований. Результаты этих работ нашли отражение в трех томах справочного издания "Основы палеонтологии" (1962-1964 гг.), посвященных ископаемым позвоночным. Здесь прослежены эволюционные преобразования скелета у разных групп животных, введены гистологические характеристики в систематику ряда групп, в особенности рыбообразных и рыб. Рассматриваемая сводка послужила источником для дальнейшего развития эволюционного подхода к оценке гистологических преобразований в истории позвоночных [185] и его практического использования в стратиграфических целях.
Современные методики исследования субфоссильных и ископаемых тканей описаны в ряде работ [793, 1972, 2033, 2035, 2042 и др.]. Они предусматривают изготовление шлифов и (или) после декальцификации ткани - срезов на микротомах, изучаемых под биологическим и поляризационным микроскопами, применение электронной микроскопии, рентгеновского анализа и микрорадиографии. Применение этих методик позволяет изучать ткани на микро- и ультраструктурном уровнях [1957, 2038, 2039, 2041 - 2044, 2107, 2134, 2135 и др.], прослеживать происхождение однотипных тканей в разных группах и создавать сравнительные модели гистогенезов. В течение последних 10-15 лет появились новые классификации костных, дентиноподобных, гиперминерализированных эмалеобразных и эмалевых тканей [2038, 2039, 2041-2044, 2134, 2135, 2162-2167]. Список наиболее важных работ в данной области опубликовал Т. Эрвиг [2043].
Сохранность тканей в ископаемом состоянии позволяет не только проследить многие детали их первоначальной структуры, такие, как полости клеток, канальцы-отростки клеток, коллагеновые волокна, положение кровеносных сосудов, нервов и соединительных волокон, обычно заполненных вторичным веществом, но также использовать метод тройного параллелизма для решения многих спорных вопросов эволюции скелетных образований. Характеристики основных типов тканей позвоночных, встречающихся в ископаемом состоянии, представлены на табл. 2-5.

Таблица 2. Дентин и дентиновые твердые ткани
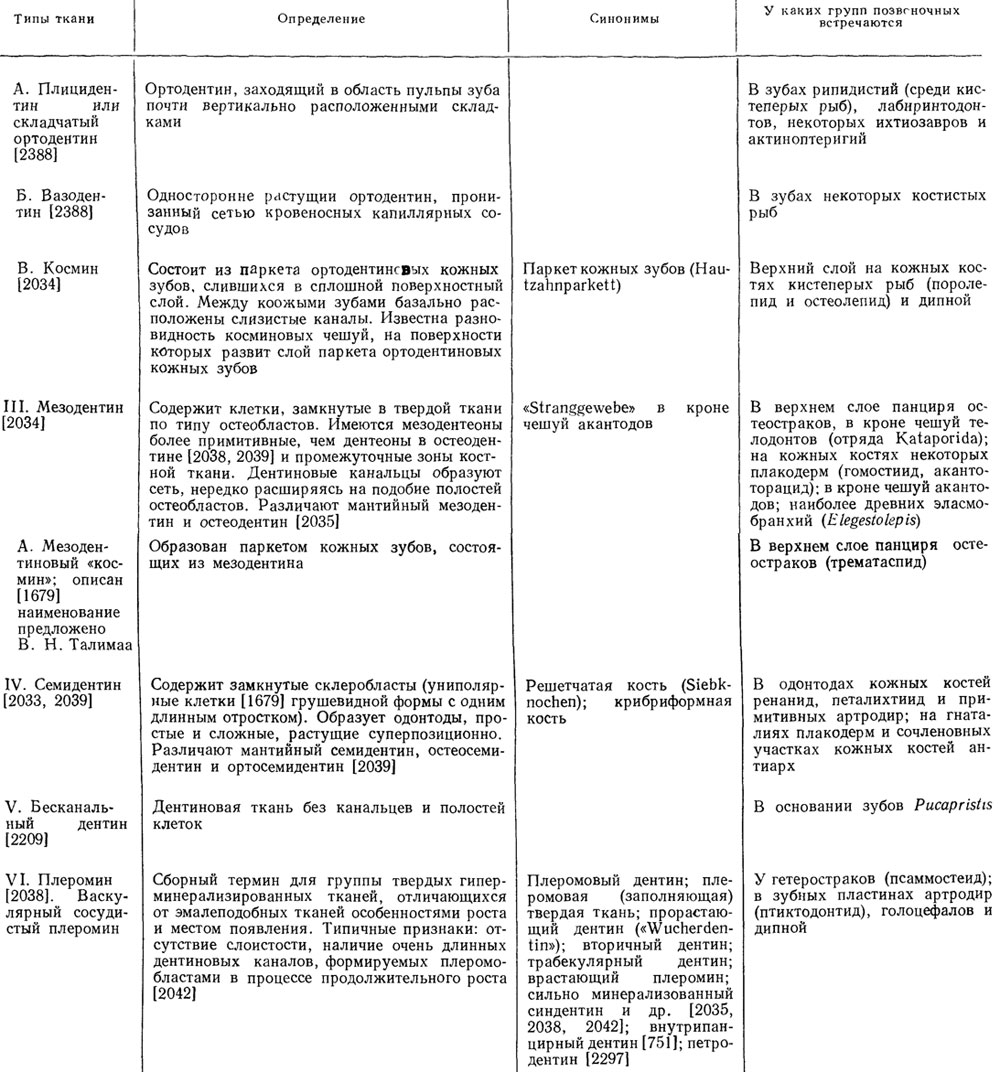
Продолжение табл. 2
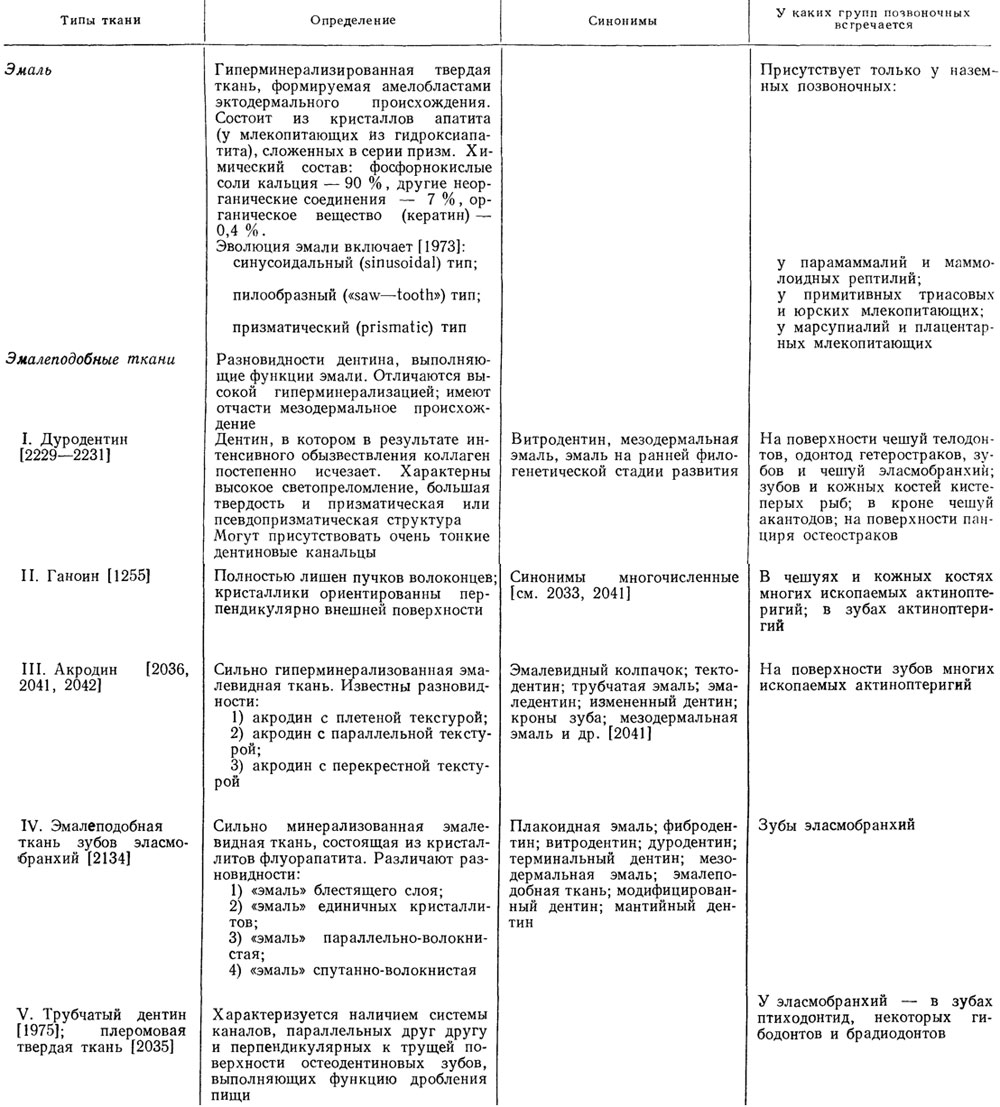
Таблица 3. Эмаль и эмалеобразные ткани
![Таблица 4. Классификации костной ткани. Классификация Ф. Вайденрайха [2459, 2460, 2462]](pic/000072.jpg)
Таблица 4. Классификации костной ткани. Классификация Ф. Вайденрайха [2459, 2460, 2462]
Классификация В. Гросса [1676]

Таблица 4. Классификации костной ткани
Примечание. Костная ткань представляет собой продукт дифференциации мезенхимы, характеризуется парой специфических костных элементов - остеобластами и остеокластами, а также основным обызвествленным волокнистым веществом, минеральная фаза которого состоит преимущественно из гидроксиапатита.
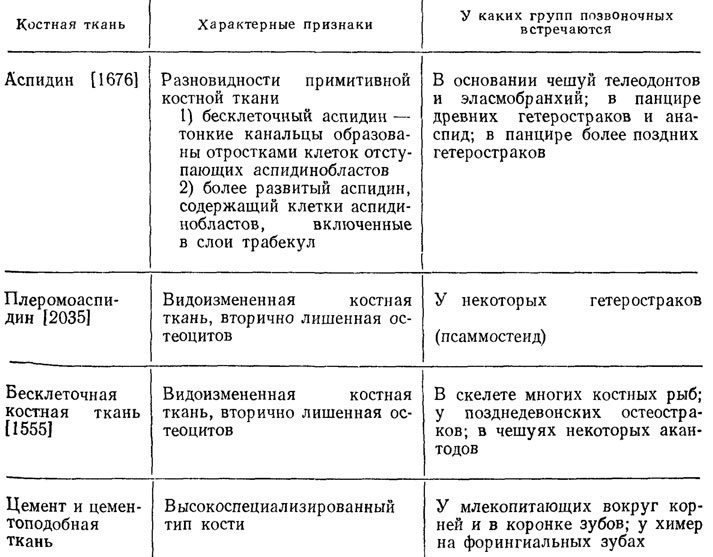
Таблица 5. Некоторые типы костных тканей, не вошедшие в классификацию А. Риклеса (см. табл. 4) и широко представленные у ископаемых и рецентных позвоночных
Согласно Ф. Ж. Менье [1948], фактический материал, накопленный неонтологами и палеонтологами, свидетельствует об отсутствии существенных различий между скелетными тканями у современных и ископаемых позвоночных: основные типы тканей остаются теми же, как и клетки, которые их продуцируют. Однако, очевидно, что у современных форм нет существовавших ранее, по данным Т. Эрвига [2033, 2035], нескольких разновидностей дентина и аспидина.
Теория биологической минерализации рассмотрена применительно к позвоночным животным во многих работах [1780, 1928, 1929, 1973, 2033, 2034 и др.]. Поэтому мы остановимся лишь на основных ее положениях, касающихся процессов обызвествления и окостенения.
Исходно такие твердые ткани, как костные, дентин и эмаль, образованы кристаллами фосфорнокислых солей - гидроксиапатита, который при фоссилизации переходит в более устойчивый фторапатит. Установлено, что обызвествление основной органической массы может протекать двумя различными путями: сферическим (шаровым) и инотропным (однородным). Но в настоящее время нельзя с полной уверенностью сказать, какой из этих путей был первичным. Т. Эрвиг [2033, 2035] полагал, что исходным следует признать сферическое обызвествление. М. А. Мосс [1972] же считал более примитивной инотропную кристаллизацию.
Согласно Т. Эрвигу [2033, 2035], механизм формирования твердых тканей у разных хордовых протекал примерно так же, как у предковой группы беспозвоночных организмов кристаллизация развивалась в мезодерме сферическим путем. На смену этому пути пришло инотропное образование твердых тканей, типичное для большинства позвоночных, где необходимым элементом гистогенеза являются активные склеробластовые клетки. Последние выделяют особые коллагеновые нити, взаимодействующие с ионами кальция и фосфора и дающие начало ориентированным по ним ядрам кристаллов апатита.
Обызвествление чешуи у современных костных рыб происходит путем инотропии [1948]. Сферическая кальцификация встречается у них крайне редко и обнаружена пока лишь в периферийной части чешуи Salmo и некоторых других рыб.
Развитие и структура костной ткани. Гистогенез костной и дентиновой тканей, а также замещение хрящевой ткани костной в эндоскелете изучены достаточно детально [185, 190, 894, 1129, 2462, 2463, 2073].
Формирование грубоволокнистой костной ткани (см. табл. 6) начинается обособлением остеогенных участков в мезенхиме, увеличением количества и толщины коллагеновых волокон. Далее следует разрастание последних и превращение мезенхимного вида клеток в клетки с маленькими отростками - остеобласты. Возникновение коллагеновых волокон между остеобластами приводит к их замуровыванию, однако полностью связь между соседними клетками не прерывается, так как они соединяются друг с другом своими отростками. Вместе с набуханием и гомогенизацией волокон в основном веществе развивающейся ткани появляются соли кальция и начинается процесс обызвествления, что приводит к обособлению периоста.
Дифференцировка пластинчатой костной ткани в онтогенезе (см. табл. 6) происходит позднее, чем грубоволокнистой, причем ее формирование обусловлено наличием определенным образом ориентированных кровеносных сосудов, вокруг которых параллельно располагаются коллагеновые волокна, образующие концентрические пластины. В пределах этих пластин (между волокнами) накопляется цементирующее их основное вещество, а между рядами пластин замуровываются обособившиеся остеобласты.
Пластинчатая костная ткань вокруг кровеносного сосуда обычно представлена совокупностью остеонов или гаверсовыми системами (рис. 64, а). В последующей перестройке костной ткани происходит разрушение ранее образовавшихся остеонов и развитие новых (см. рис. 64, б), причем в этом процессе принимают участие остеокласты.
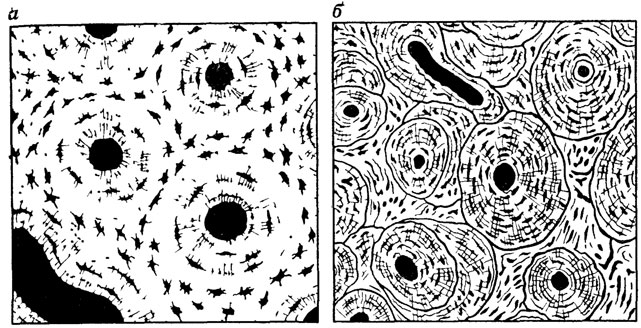
Рис. 64. Костная ткань. По Т. Эрвигу. а - с первичными остеонами; б - со вторичными остеонами
В тканях эндоскелета необходимо различать костную ткань перихондрального, периостального, мезостального, энхондрального и эндостального происхождения.
Изучить детально гистогенез костной ткани и усложнения микроструктуры костей только на ископаемом материале весьма трудно. Сравнительное изучение костной ткани и микроструктуры отдельных костей современных и ископаемых животных [1948, 2162-2167] показало, что типы гистоструктуры костных тканей у современных и ископаемых животных одни и те же. В частности, описанные Ф. Вайденрайхом [2459, 2460, 2462] у современных форм грубоволокнистая, скорлуповая (пластинчатая) и смешанный тип костных тканей (см. табл. 6) впоследствии были обнаружены В. Гроссом [1676] у ископаемых животных. На основании изучения микроструктуры костной ткани у стегоцефалов и рептилий В. Гросс и предложил свою классификацию (см. табл. 6).
Еще более детальную классификацию типов костного вещества и костных тканей разработал А. Риклес [2163], который показал соответствие гистроструктуры ископаемых и современных типов костных тканей (см. табл. 6). Доказательством этого является возможность сведения к синонимам типов гистроструктуры костных тканей ископаемых к типам гистроструктуры современных позвоночных.
Согласно Ф. Ж. Менье [1948], у костных рыб скелетные ткани (в частности, костные ткани и их производные) образуют широкий континиум вариаций, которые могут проявляться в одной и той же анатомической структуре, одном и том же организме, а также костях различных видов современных, субфоссильных [793] и ископаемых тетрапод [2162-2167]. На основании пространственной организации коллагеновых фибрилл Ф. Ж. Менье выделил у костных рыб следующие типы костной ткани 1) волокнистая с беспорядочно ориентированными волокнами; 2) с параллельными волокнами (псевдоламеллярная), 3) ламеллярная [1948].
И. А. Риклес и Ф. Ж. Менье не считают возможным рассматривать эволюцию костных тканей с позиций биогенетического закона. По их мнению, особенности гистроструктуры этих тканей можно объяснять лишь функционально. Поэтому А. Риклес [2167] предложил вместо понятия "эволюция костных тканей" другое понятие "история костных тканей", различие между которыми очевидно. Значение функционального момента в формировании гистоструктуры было также подчеркнуто И. Шаффером [2216] и К. Л. Паавером [793].
Поскольку костную ткань невозможно изучать вне соответствующих анатомических структур (отдельных костей), в последнее время в учебниках гистологии [1129] введен специальный раздел "Костная ткань и кости". Такое различие имеет определенный смысл, поскольку иначе зачастую затушевываются особенности костных тканей различного происхождения и появляются неверные термины - например, периостальная кость, перихондральная кость, энхондральная кость, остеонная кость и др. На самом деле речь идет в данном случае о костной ткани перихондрального, периостального и энхондрального происхождения и о ее остеонной организации. Различия между костью-органом и ее тканевыми компонентами различного происхождения можно показать на схеме продольного разреза длинной кости у позвоночных (рис.65). Другим примером может служить мембранозная кость-орган, которая в эндоскелете первичноводных анамний и некоторых амфибий состоит из костной ткани периостального происхождения; у остальных тетрапод в лакунах мембранозных костей может присутствовать кроветворный костный мозг.
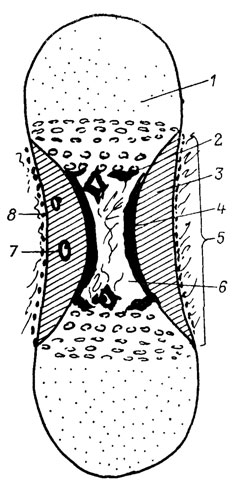
Рис. 65. Обобщенная схема микроструктуры длинной кости позвоночных на продольном срезе. По В. П. Пегете. 1 - хрящевой эпифиз; костная ткань: 2 - энхондрального происхождения, 3 - периостального происхождения, 4 - эндостального происхождения; 5 - костный диафиз; 6 - медуллярная полость; 7 - остеон (костная ткань мезостального происхождения; 8 - волокнистый периост
Развитие и структура дентиновой ткани. Дентин (в широком смысле) представляет собой модификацию костной ткани [762]. Продуцирующие его одонтобласты отличаются от остеобластов тем, что не включаются в твердую ткань, а размещаются на стенках внутренней поверхности дентеона. От каждого одонтобласта к наружной поверхности дентеона отходит длинный протоплазматический отросток, расположенный в дентиновом канальце (см. табл. 4).
Начальные этапы гистогенезов дентина и различных дентиноподобных тканей имеют в общем те же стадии развития, которые проходит и костная ткань (рис. 66, 67).
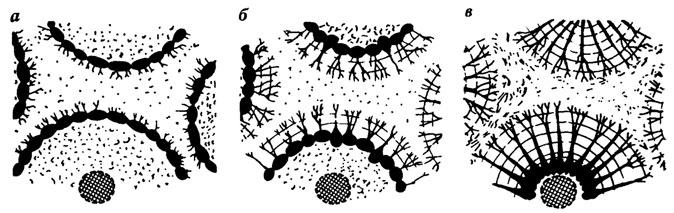
Рис. 66. Три стадии (а-в) развития остеодентина. По Т. Эрвигу
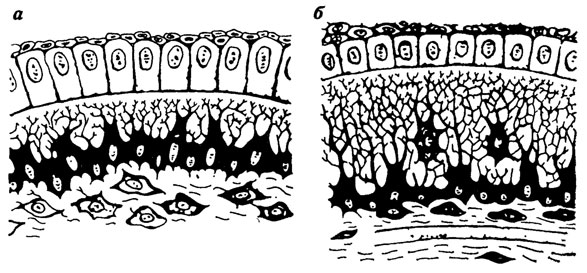
Рис. 67. Две стадии (а-б) формирования паллиального мезодентина. По Т. Эрвигу
Формирование остеодентина происходит вокруг сети кровеносных сосудов, а ортодентина - в одной из полостей пульпы (чешуя телодонтид, зуб эласмобранхий, фтентеон на поверхности кожных костей). Однако как в первом, так и во втором случае дентиновая ткань образована пластинками, состоящими из коллагеновых волокон, между которыми в радиальном направлении протягиваются дентиновые канальцы с расположенными в них протоплазматическими отростками одонтобластов (отростками Томеса).
Для гистогенеза мезодентина (см. табл. 4) характерно то, что не все клетки одонтобластов "отступают" в глубь пульпы одновременно. Некоторые из них замуровываются подобно тому, как это происходит при образовании костной ткани (см. рис. 67).
Типичным признаком дентина и почти всех дентиноподобных тканей является слоистость, образование которой обусловлено последовательной минерализацией, начинающейся с поверхностного слоя (отложение солей кальция происходит ритмично расположенными слоями). Исключение представляют плеромин, характеризующийся отсутствием слоистости (рис. 68), что свидетельствует о непрерывности его роста [2043], и петродентин [2297], образующий зубные пластинки дипной и характеризующийся двухфазовым ростом. Петродентин является уникальным для дипной по способу образования. Он продуцируется исключительно в мезодермальных клетках - петробластах (без участия эпителия), хотя по микроструктуре сходен с энамелоидом в зубах эласмобранхий и лучеперых. Это отличие объясняется изменениями в клеточной дифференциации.
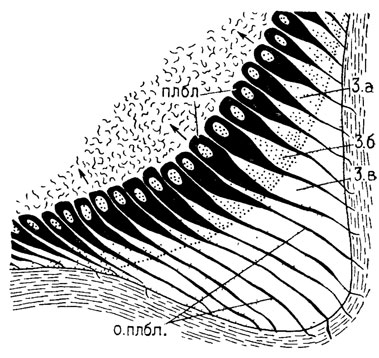
Рис. 68. Развитие плеромина в зубной пластинке Ctenurella gladbachensis Orvig. По Т. Эрвигу. плбл - плеромобласты, о. плбл - отростки плеромобластов, 3.а, 3.б, 3.в - зоны роста
Среди дентиновых образований особый интерес представляют лабиринтодонтные зубы у кистеперых рыб, амфибий и рептилий. Различают три основных типа складчатости этих зубов - полиплакодонтный, эустенодонтный и дендродонтный (рис. 69), характеризующиеся неодинаковой складчатостью и строением пульпарной полости, отсутствием (или наличием) в ней балок остеодентина [184, 186, 1393, 2246]. Эти типы обычно используются для филогенетических характеристик, в частности для различения крупных таксонов. Так, остеолепиды и ризодонтиды характеризуются полиплакоидным и эустеноидным типами микроструктуры зубов, а поролепиды - дендроидным типом (рис. 70). Этот признак важен и для обоснования происхождения тетрапод от единого - остеолепидного - ствола [2247]. Правда, дальнейшие исследования [2432] показали, что все рассматриваемые типы микроструктуры зубов связаны переходами так, что четкая грань между ними (особенно между полиплакодонтным и эустеноидным) стирается. При этом весьма сходная микроструктура может прослеживаться не только в таких далеких филумах, как рыбы, амфибии и рептилии, но также в пределах разных отрядов и семейств кистеперых рыб. Так, микростроение зубов триасового ихтиозавра и каменноугольной кистеперой рыбы Picnoctenium сходно. Подобное палеогистологическое сходство с далекими филумами может, очевидно, рассматриваться как результат параллелизмов [189].
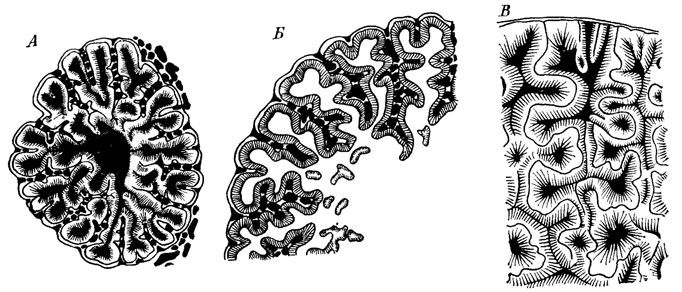
Рис. 69. Типы строения зубов у кистеперых рыб. По Г. П. Шультце. А - полиплакодонтный тип у рода Panderichthys, Б - эустенодонтный тип у рода Platycephalichthys (по Э. И. Воробьевой), В - дендродонтный тип у рода Porolepis

Рис. 70. Распространение типов складчатых зубов во времени и направления их филогенеза. По Г. П. Шультце
Природа эмалеобразных тканей (см. табл. 5) пока не имеет единого объяснения. Вероятно, они представляют собой гетерогенную группу. Дуродентин, например, является поверхностным слоем дентиновой ткани и развивается под базальной мембраной. Эмалеобразные же ткани типа ганоина (?) и акродина налегают на дентин и, возможно, формируются амелобластами [2248]. Рост этих тканей происходит подобно эмали - центробежно над базальной пластиной.
Концепции эволюционной гистологии. Среди эволюционных направлений гистологии, созданных в 20-40-е годы отечественными учеными [697], особенно выделяются альтернативные позиции школ А. А. Заварзина [379-381] и М. Г. Хлопина [1124].
Концепция А. А. Заварзина исходит из принципа параллельного развития гистоструктур в разных типах животного мира и несоответствия высоты организации тканей положению организмов в системе. Согласно этой концепции, ткани должны развиваться прежде всего по специфическим законам, определяющим их принадлежность к той или иной тканевой системе.
Концепция М. Г. Хлопина, напротив, исходит из представления о дивергентной эволюции тканей, где их специализация носит в основном адаптивный характер и ставится в онто- и филогенезе в прямую зависимость от источника происхождения связанных с этими тканями органов.
Особый подход к изучению эволюции тканей предложен А. В. Румянцевым [894], который руководствовался теорией филэмбриогенеза А. Н. Северцова и теорией целостности развития организма И. И. Шмальгаузена. Рассматривая теорию филэмбриогенеза как метод изучения тканевой эволюции, А. В. Румянцев пришел к выводу, что этот метод применим в рамках одного типа. В тех же случаях, когда эволюция тканей исследуется в пределах разных типов многоклеточных, плодотворен сравнительно-гистологический метод А. А. Заварзина, основанный на принципе параллелизма гистоструктур. Однако тканевые параллелизмы широко прослеживаются в скелете разных групп позвоночных [185, 189, 753, 814 и др.].
Согласно А. В. Румянцеву, на тканевом уровне не существует полного параллелизма между онтогенезом и филогенезом "Повторение стадии, - писал он, - совсем не означает, что трубчатая кость млекопитающих проходит последовательно стадии онтогенеза рыб и рептилий" [894, с. 307]. Основным модусом преобразований костной и хрящевой тканей А. В. Румянцев, считал анаболию. Детальное изучение развития и дефинитивной микроструктуры преформированных хрящем костей у представителей разных классов позвоночных [2073] показало, что эволюция костей может происходить не только путем анаболии, но и путем девиации за счет коррелятивных связей между хрящевой, костной и миелоидной тканями. А. Н. Северцов признавал за гистогенезами и определенную специфику в их постоянстве и устойчивости, обусловленную определенной автономностью тканевых систем.
А В. Румянцев относил к автономным системам хрящевую ткань, которую рассматривал как систему низшего порядка. Автономность и свойство самодифференцировки тканей трактуется им в качестве древнего приобретения. При этом А. В. Румянцев обращал внимание на автономность клеток внутри тканей и автономность тканей внутри органов, что расценивалось как необходимые предпосылки эволюционного усложнения на тканевом уровне. Хрящ и кость, согласно этому исследователю, в пределах подтипа позвоночных подчиняются исключительно общим филогенетическим и морфологическим закономерностям, тогда как на уровне беспозвоночных в развитии этих тканей обнаруживается параллелизм. Хрящ и кость, развиваясь у позвоночных путем эволюционных дифференцировок из скелетной мезенхимы, преобразуются в тесном взаимодействии по пути постепенной замены хряща костной тканью в эндоскелете. Такая замена происходит в результате специфических морфогенетических влияний, исходящих от самого хряща.
А. В. Румянцев, как и многие другие отечественные ученые, не разделял бытовавшие долгое время в сравнительной анатомий и палеонтологии представления о хряще как о деградации костной ткани.
Разная степень автономности тканей отмечалась и И. И. Шмальгаузеном [1187]. В частности, им признавалась автономность покровных тканей, которая обеспечивалась их перестройками преимущественно по пути архаллаксисов, т. е. качественных изменений зачатков в онтогенезе при относительной редкости рекапитуляции. Отсюда следует относительная лабильность автономных тканевых систем.
Новые фактические материалы по эволюции скелетных тканей в палеонтологических рядах позвоночных позволяют внести некоторые дополнения и уточнения в существующие концепции тканевого уровня.
Соотношение экзо- и эндоскелета. Центральное место в палеогистологических исследованиях позвоночных занимает вопрос о соотношениях между экзо- и эндоскелетом, а в конечном счете - к проблеме соотношений между хрящевой и костной тканями в онто- и филогенезе. До недавнего времени под экзоскелетом (дермоскелетом) подразумевались все наружные скелетные образования, а под эндоскелетом - все внутренние скелетные части в интерпретации К. М. Бэра [1290], предложившего термины "наружный" и "внутренний" скелет. Мнение об отсутствии хряща в экзоскелете было общепринятым.
А. Н. Северцов [925, 931], как и К. Гегенбаур [1630], признавал, что хрящ в филогенезе появился раньше кости.
Э. А. Стенше [2328, 2329] и его последователи считали, что у предков современных эласмобранхий в эндоскелете имелась кость и что костный эндо- и экзоскелеты возникли одновременно.
В конце концов сложилась компромиссная точка зрения [185, 381, 894, 1496, 1973]: хрящ и кость в филогенезе появились независимо друг от друга (хрящ - в эндоскелете, кость - в экзоскелете); при этом кость в эндоскелете появилась позже.
В тканевом составе экзоскелета позвоночных в процессе их длительной эволюции происходили изменения; количественные соотношения между хрящевой и костной тканями в эндоскелете также не были постоянными (рис. 71).

Рис. 71. Схема эволюции скелетных тканей в эндо- и экзоскелете низших позвоночных по новейшим палеонтологическим и неонтологическим данным. По Э. И. Воробьевой (с уточнением В. Н. Каратаюте-Талимаа и дополнениями В. П. Пегеты). 1 - экзоскелетные (сверху) и эндоскелетные (снизу) ткани, 2 - дентин и его разновидности, 3 - аспидин, 4 - костная ткань (экзо), 5 - костная ткань (эндо), 6 - хрящевая ткань; 7 - резорбция хрящевой ткани, 8 - костная ткань энхондрального происхождения
Определенный диссонанс в проблему соотношений между хрящевой и костной тканями внес К. Паттерсон [2070]. Его критические замечания по поводу применения специалистами различного профиля одной и той же терминологии к костным образованиям кажутся вполне справедливы, хотя трудно согласиться сего формулировкой, что "хрящ имеется в дермоскелете". К. Паттерсон назвал три источника существующих недоразумений: 1) термины употребляются в работах, касающихся разных уровней организации тканевого - в гистологической литературе, органного (мембранные и хрящевые кости) и систем органов (дермальный скелет) - в анатомической литературе; 2) под дермальными и мембранными костями подразумевается одно и то же; 3) термины "хрящевые" и "энхондральные кости" используются как синонимы.
Для обозначения костей-органов К. Паттерсон предложил использовать следующие понятия [2070].
1. Хрящевые кости - кости, преформированные в хряще и оссифицирующиеся в онтогенезе путем развития перихондрального кольца или диска на поверхности хрящевой модели. Позже они могут оссифицироваться периостально или энхондрально и могут развивать во время роста мембранную кость.
2. Дермальные кости - кости, не преформированные в хряще, лежащие на базальной мембране вблизи эктодермы и несущие на своей поверхности дентин и (или) эмалеподобные образования, а также "гомологичные" [1791] им кости (тека и эпитека черепах, броня броненосцев и т. д.). У млекопитающих и птиц дермальные кости могут переходить в адвентиционный хрящ.
3. Мембранные кости - кости, которые развиваются в мезодерме без онто- и филогенетической связей с эктодермой. Мембранные кости могут быть пластинчатыми, губчатыми или их комбинациями.
По мнению К. Паттерсона, следует различать два вида костей: гомологичные хрящевым костям примитивных рыб; новообразования в малых таксономических группах и патологические образования костей.
Экзоскелет и его происхождение. Все известные твердые ткани и тканеподобные образования экзоскелета позвоночных являются производными эпидермиса (волосы, рога, кератиновые зубы, эмаль), эпидермиса и эктомезенхимы (зубы, чешуи, дентиновые слои дермального кожного скелета, плавниковые лучи) и мезодермы (дермальные кости, рога оленей, мезодентин зубных пластинок дипной). Их формирование обычно начинается на границе эктодермального или энтодермального эпителия с мезодермой (эктомезенхимой). Именно здесь возникает базальная мембрана, от которой происходит дальнейшее центробежное развитие ткани определенного типа, причем особенно важную роль играют базальные клетки эпидермиса [1973].
Дальнейший ход процесса толкуется неоднозначно. Согласно принципу "деляминации" (расслоения), выдвинутому Н. Хольмгреном [1754] и детально разработанному затем Э. Ярвиком [1794], каждая новая генерация ткани, возникшая у основания базальной пластины, постепенно опускается в более глубокие слои кожи, уступая место последующим генерациям. При этом типы тканей в последовательно появляющихся генерациях могут быть неидентичными, как, например, в сложных тканях кожного скелета с циклическим строением (дентиновая ткань-кость). Предполагается в то же время, что между разными типами экзо- и эндоскелетных образований не могло существовать фундаментальных различий [1794].
Г.-П. Шультце [2248] считал, что возникновение различных разновидностей твердых тканей зависит от глубины их формирования в кориуме (коже). Так, костная ткань в чешуе лучеперых и, возможно, кистеперых рыб образуется в нижнем и среднем кориуме, а в верхнем кориуме, под эпидермисом, развиваются униполярные клетки - одонтобласты, продуцирующие дентиновые ткани центростремительно от эпидермиса вплоть до срастания с костной тканью. Данный путь формирования кожного скелета предусматривался, по-видимому, и в отношении гетеростраков и артродир [1497, 1498, 2484], у которых прослеживается независимое развитие поверхностного и более глубоких слоев панциря.
Уже более 100 лет привлекает внимание исследование закономерностей формирования и эволюции кожного окостенения - экзоскелета. В отличие от монофилетической теории происхождения чешуи [1732] Е. К. Гудрич [1656], учитывая данные У. К. Вильямсона [2494] о наличии в чешуях разных рыб различных компонентов, развил теорию о полифилетическом происхождении чешуи, получившую впоследствии наименование теории Вильямсона - Гудрича. Вместе с тем долгое время преобладала точка зрения Э. А. Стенше [2329], полагавшего, что чешуйный покров в линии телодонты - гетеростраки является результатом распада (редукции) сплошного кожного окостенения. Позже Д. В. Обручев [761] обосновал концепцию, согласно которой большие элементы панциря представляют собой продукт слияния плакоидных чешуи и тессер.
Новым импульсом к возобновлению дискуссии по этому вопросу явилась лепидомориевая теория, краеугольные положения которой были высказаны Б. Ф. Матвеевым [630, 1930], в завершенном же виде она была сформулирована Н. Хольмгреном [1754]. Согласно этой теории, образование костного скелета позвоночных связано с первичными элементами экзоскелета - лепидомориями, формирующимися в кориуме на начальных стадиях гистогенеза вокруг петли кровеносного сосуда и состоящими из дентиновой кроны, базальной пластинки и пульпарной полости (рис. 72). Возникновение разнообразных типов чешуи и тессер объясняется сращением разного числа лепидомориев, которое может протекать двумя путями: 1) цикломориальным - лепидомории срастаются в основании последовательно, их пульпарные полости остаются самостоятельными, а новые первичные элементы экзоскелета в виде ареальных зон роста концентрируются вокруг первичного лепидомория (примордиума); 2) синхрономориальным - одновременное срастание лепидоморий до стадии обызвествления и образование твердой дентиновой ткани над всей группой крон. Пульпарная полость такой структуры разделяется на участки с разным количеством слившихся лепидомориев. Кроме этих основных типов роста экзоскелета возможны их различные комбинации.
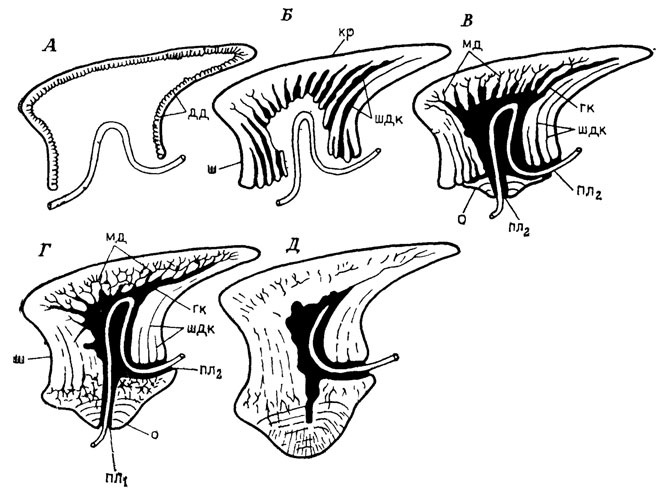
Рис. 72. Онтогенез чешуи Elegestolepis grossi Kar. - Tal. По Б. Н. Каратаюте-Талимаа. А - папиллярная стадия; Б - ювенильная стадия без основания; В - ювенильная чешуя с основанием; Г - взрослая чешуя; Д - старая чешуя с заросшим пульпарным отверстием в основании; дд - дуродентин; мд - мезодентин; пл1 - пульпарное отверстие в основании; пл2 - пульпарное отверстие шейки; гк - главный канал; шдк - широкие дентиновые каналы, кр - крона; о - основание; ш - шейка
На основе изучения гистологии скелета эласмобранхий Э. А. Стенше [2332, 2333] утверждал, что девонские и каменноугольные их представители обладали цикломориальными чешуями, позднепермские - цикломориальными и синхрономориальными, а все более молодые - исключительно синхрономориальными. Он считал цикломориальный тип роста чешуи примитивным, а синхрономориальный - специализированным. Аналогичный подход к изучению скелета был использован Л. Б. Тарло [2355-2357], который разработал схему эволюции гетеростраков, не получившую, однако, признания.
Лепидомориевая теория вызвала большие дискуссии, в ходе которых был выявлен ряд ее недостатков [1497, 1506, 2136-2138, 2484 и др.]. В частности, исследования чешуи эласмобранхий, тессер и панцирей гетеростраков, артродир, актиноптеригий и других групп показали, что в процессе эволюции кожных окостенений у этих первичноводных животных происходила как ассимиляции, так и распад элементов панциря. В этой связи цикломориальный тип роста не следует считать более примитивным, чем синхрономориальный. Находки в верхнем силуре Тувы [1814], а позже и в нижнем силуре (лландовери). Иркутского амфитеатра* чешуи эласмобранхий, представляющих простые лепидомории, синхрономориальных чешуи гетеростраков в нижнем ордовике Шпицбергена [1351] и Австралии [2170] и аналогичных чешуи эласмобранхий в нижнем девоне Подолии и Шпицбергена [471, 1660], обнаружение большого разнообразия типов роста чешуи постпермских селяхий [2136, 2137] и палеозойских кроссоптеригий [186] позволили опровергнуть приведенные выше представления Э. А. Стенше и Л. Б. Тарло о единых линиях со сменой типов роста.
* (Данные Л. И. Новицкой и В. Н. Талимаа.)
Дальнейшая разработка лепидомориевой теории связана с разграничением стадий развития экзоскелета и выделением ассимилятивной и регрессивной фаз в его эволюции [2035, 2040 и др.]. Согласно этим представлениям, степень ассимиляции кожного скелета, наблюдаемая у различных групп низших позвоночных, достигнута независимо и вне соответствия уровню их организации, причем фаза регрессии скелета сопровождалась обычно увеличением подвижности животных.
Для обозначения составных структур кожного скелета, так называемых "кожных зубов" (дотетраподного уровня) введен термин "одонтод" [2035, 2040], широко используемый гистологами. Следует подчеркнуть, что одонтод не принадлежит к озублению и не выполняет функции зубов. Одонтод образован дентином или дентиновыми тканями и часто покрыт поверхностным эмалеподобным слоем. Формирование его начинается с одной папиллы мезенхимальных мягких тканей, окруженных снаружи эпителиальным слоем эпидермиса.
Т. Эрвиг предложил схему роста одонтода, согласно которой развитие одонтода может осуществляться ареально и суперпозиционно. Чешую, состоящую из одного или нескольких одонтодов и костного основания, называют "одонтодиумом"; соответственно различают монодонтодии и полиодонтодии, а у периодически растущих чешуи - одонтокомплексы.
Наиболее простым путем происходило, по-видимому, формирование плакоидных чешуи (монодонтодия) у телодонтов [472]. Началом этого процесса могла служить закладка поверхностного слоя дуродентина под базальным слоем эпидермиса в пределах папиллы и центростремительное нарастание слоев ортодентина у телодонтиформных или мезодентиноподобной ткани у катопориформных телодонтов (рис. 73). Далее происходило врастание чешуи в слой мезодермы и образование аспидинового основания, в котором располагались шарпеевские волокна, соединявшие чешую с кожей. Для такого наиболее примитивного типа чешуи характерны постоянные размеры на протяжении онтогенетического развития (нерастущая чешуя). После отмирания (выпадения) чешуи на ее месте происходила закладка более крупной чешуи следующей генерации.
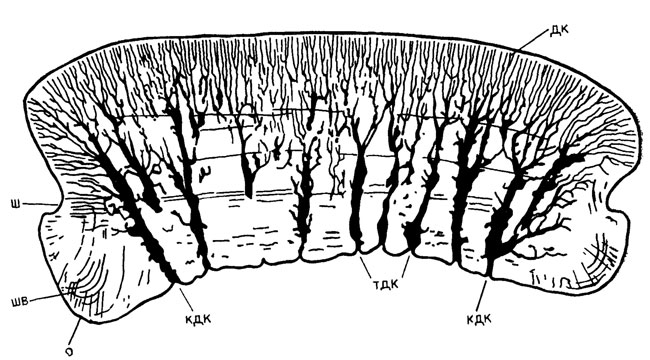
Рис. 73. Строение мезодентиноподобной ткани в чешуе катапоридного телодонта Katoporus timanicus Kar. - Tal. Вертикальный продольный срез чешуи. По В. Н. Каратаюте-Талимаа. ш - шейка; о - основание; дк - дентиновые канальцы; тдк - толстый дентиновый канал; кдк - краевой дентиновый канал; шв - шарпеевские волокна
Другим примером монодонтодия с костным основанием может служить чешуя эласмобранхий типа Elegestolepis (см. рис. 72), рост которой происходил как центростремительно, так и центробежно [1814]. Это типичные нерастущие чешуи, возникшие над сосудистой петлей, состоящие из кроны, шейки и основания, обладающие пульпарной полостью и двумя пульпарными отверстиями - базальным и шейным. После закладки дуродентинового поверхностного слоя формирование ортодентиновой и мезодентиновой (?) тканей кроны шло центростремительно. Позже, на определенном этапе развития кроны, закладывалось небольшое костное основание и дальнейший его рост происходил уже центробежно в глубь мезодермы. После зарастания базального пульпарного отверстия чешуя отмирала и выпадала так же, как у телодонтов.
Концепция одонтода успешно применена и при исследовании чешуи акул, в которой различают нерастущие (простые лепидомории и синхрономориальные) и растущие (цикломориальные) чешуи [2133, 2134, 2136, 2137].
Чешуи акантодов можно рассматривать как своеобразный одонтокомплекс суперпозиционного и отчасти ареального типов роста. Закладка эмбриональной чешуи происходила на границе эпидермиса и мезодермы. Крона образована слоями мезодентиновой ткани и дуродентина, а основание - клеточной или бесклеточной костной тканью, иногда пронизанной восходящими сосудистыми каналами [137, 1679]. У некоторых родов акантодов в кроне развита сложная система поровых и слизистых каналов. Рост чешуи происходил центробежно путем образования новых концентрических слоев поверх существующих.
Определенный вклад в разработку лепидомориевой теории внесли сравнительные исследования разных групп гетеростраков [752], показавших, что формирование цикломориальных тессер подвержено индивидуальной изменчивости, определяется свойствами дермального покрова и что бугорки на гребешках скульптуры некоторых гетеростраков (у птераспид), объясняемые как первичные лепидомории, являются на самом деле вторичными образованиями.
Эволюция экзоскелета. О филогенетической последовательности экзоскелетных твердых тканей не существует единого мнения. Так, Т. Эрвиг [2035] полагает, что первична костная ткань, а дентин и аспидин (бесклеточная кость) - ее производные. Ряд других исследователей [894, 1496, 1973, 2358] считают дентин и дентиноподобные ткани исходными в эволюции. Установлено, в частности, что панцирь ранне- и среднеордовикских гетеростраков состоит из ортодентина и аспидина. Недавние находки В. Н. Каратаюте-Талимаа в основании силура Сибирской платформы плакоидных чешуи трех типов (логаниидного, характерного для наиболее древних телодонтов, а также акантодного и эласмобранхиевого, в частности чешуи Elegestolepis) послужили основанием для пересмотра увеличения возраста мезодентиноподобной ткани и аспидина. Кроме того, находки чешуи данного типа указывают на возможность появления всех крупных групп бесчелюстных и рыб, по-видимому, уже в кембрии и даже, возможно, в докембрии.
Высказано предположение, что древние экзоскелетные ткани, обнаруживая разнообразие состава, находились на стадии незаконченной дифференциации, которая происходила независимо в разных группах [185].
Эволюцию экзоскелетных тканей изучали многие специалисты (А. П. Быстров, Д. В. Обручев, Т. Эрвиг, В. Гросс и др.), большинство из которых исходной группой в развитии стволов бесчелюстных считают телодонтов в широком смысле - позвоночных, покрытых дискретными элементами кожного скелета. Иначе говоря, подразумевается, что "...древнейшие позвоночные не имели окостенений во внутреннем скелете, а наружный состоял из дискретных чешуек, пластинок и отдельных кожных зубов" [764, с. 343]. По А. В. Румянцеву [894], основной филогенетический ствол по характеру тканевых перестроек скелета включал в восходящем порядке следующие группы: остракодермы - акантоды - антиархи - эласмобранхии - кроссоптеригии - дипнои - амфибии (стегоцефалы) - рептилии - маммалии. Соответственно он различал и стадии гистогенеза скелета, причем в качестве боковой ветви рассматриваемого ствола выделял актиноптеригии - хондростеи - голостеи - телеостеи.
В свете новейших представлений соотношение гистогенезов экзоскелета бесчелюстных имеет вид пучка ветвей, сходящихся на нециклостомных позвоночных типа телодонтов [752], которые должны рассматриваться, таким образом, в качестве сборной группы.
Работа А. В. Румянцева [894] подводила итоги эволюционных исследований гистоструктуры экзоскелета разных групп позвоночных как ископаемых, так и современных. Автор пришел к выводу, что развитие дентина у всех позвоночных происходит одинаково. Дентиноподобные ткани у рыб развиваются самостоятельно, в то время как у высших позвоночных - только во взаимодействии с покровным эпителием.
Плакоидные чешуи телодонтов и панцирь гетеростраков, по классификации А. В. Румянцева [894], следует причислить к дентиновой стадии гистогенеза.
В чешуйчатом покрове телодонтов присутствуют дентиновые ткани: ортодентин - у телодонтиформных; мезодентиноподобная ткань - у катопориформных; дуродентин, тонким слоем покрывающий снаружи крону чешуи. Основание же чешуи образовано своеобразной бесклеточной костной тканью - аспидином. Катопоридный (логаниидный) тип чешуи считается наиболее древним. Телодонтиды, появившиеся в середине силура, продолжали существовать до позднего девона.
Кожный скелет анаспид образован только аспидином. Панцирь гетеростраков с поверхности орнамента покрыт дуродентином и ортодентином, а в желобках между элементами орнамента, а также в среднем и базальном слоях состоит из аспидина. В случае редукции панциря ортодентиновый слой утрачивается. Микростроение панциря (число и форма панцирных каналов, наличие ретикулярного слоя) у гетеростраков изменчиво на всех таксономических уровнях. В ходе эволюции наблюдается вытеснение призматических камер в канцеллярном слое губчатой тканью и гистоструктуры приобретают подчас "псаммостеидный" облик [753]. У псаммостеид, как известно, установлено присутствие плеромина.
В экзоскелете остеостраков установлены мезодентин и костная ткань. В хрящевой мозговой коробке каналы и полости выстланы костной тканью. Редукция экзоскелета происходила путем утраты наружного мезодентинового слоя и отчасти костного среднего слоя. Это соответствует следующей стадии гистогенеза: включению остеоцитов в твердую ткань и образованию грубоволокнистой костной ткани. К этой стадии можно относить и кожный скелет акантодов, в котором присутствует грубоволокнистая костная ткань.
Среди плакодерм выделяются две группы; 1) ренаниды, петалихтииды и примитивные артродиры, которые характеризуются развитием в верхнем скульптурном слое семидентина [2043], а в среднем (губчатом) и базальном (изопединовом) слоях - грубоволокнистой костной ткани; 2) антиархи и те из артродир, у которых все три слоя панциря состоят из грубоволокнистой костной ткани.
На следующей стадии гистогенеза, характерной для геологически поздних артродир, происходила редукция семидентина и включение кровеносных сосудов в грубоволокнистую костную ткань губчатого слоя с образованием первичных остеонов. Кроме того, у некоторых петалихтиид и ренанид, например Plourdosteus canadensis (Woodw.), наблюдается перихондральное окостенение по всему эндоскелету и частичное обызвествление (сферическое) хрящевой мозговой коробки [1499, 2033].
В кожном скелете дипной и кроссоптеригий (саркоптеригий), хорошо различаются три слоя: верхний - ортодентиновый (иногда косминовый), средний и базальный - костный [194].
В ходе эволюции дипной и кистеперых происходила последовательная редукция дентиновых образований поверхностного слоя, вплоть до полного их исчезновения. Механизм этой редукции, также как и механизм формирования и смены поверхностного слоя в онтогенезе, остаются до конца неясными и служат поводом для дискуссий.
По всей видимости, эти механизмы, сохраняя определенную общность, претерпевали вместе с тем эволюционные преобразования, которые происходили параллельно и неравномерно в разных филумах [191]. При этом могло сохраняться примитивное состояние (например, у поролепидных кистеперых) либо развитие могло идти как у остеолепидных кистеперых по пути усложнения и обеспечения многообразия этих механизмов.
Предполагается [1678], что паркет у поролепид появлялся к моменту завершения роста рыб. У остеолепид в отличие от поролепид косминовый паркет образовывался, по-видимому, неоднократно, подвергаясь в течение жизни полной или частичной редукции - "линьке" [1793]. В качестве закладок нового космина рассматривают своеобразные вздутия - "блистеры", среди которых различают погруженные и поверхностные. Высказываются, однако, сомнения в значимости блистеров для замещения паркета. Ряд исследователей [1394, 1679] связывают блистеры с регенерацией (залечиванием повреждений) либо считают их лишь имитацией формирования косминового паркета. Оспаривается также возможность слияния блистеров из-за присутствия дуродентина [1679].
На основании наблюдений над остеолепидами Э. И. Воробьева [186] полагает, что формирование нового косминового паркета протекает одновременно с резорбцией старого, а также путем "шелушения" - отслаивания более или менее крупных кусков паркета, на месте которых одновременно мог формироваться новый паркет.
Можно допустить [191], что смена покровных костей и чешуи в онтогенезе рипидистных кистеперых рыб (поролепид и остеолепид) обеспечивалась взаимодействием разных механизмов морфогенеза: формированием последовательных генераций костных зубов с последующей их резорбцией, формированием костного паркета по цикломориевому и синхромориевому типам; локальной резорбцией (рис. 74, а) паркета и его замещением с помощью "блистеров" (см. рис. 74,6); периодическим сбрасыванием паркета путем полной или частичной "линьки" с одновременным либо последовательным развитием нового паркета на поверхности костей (см. рис. 74, в, г), локальным отслаиванием - "шелушением" косминового и костного слоев (рис. 75, а); развитием дополнительных окостенений над покровными костями по принципу деляминации (см. рис. 75, б).
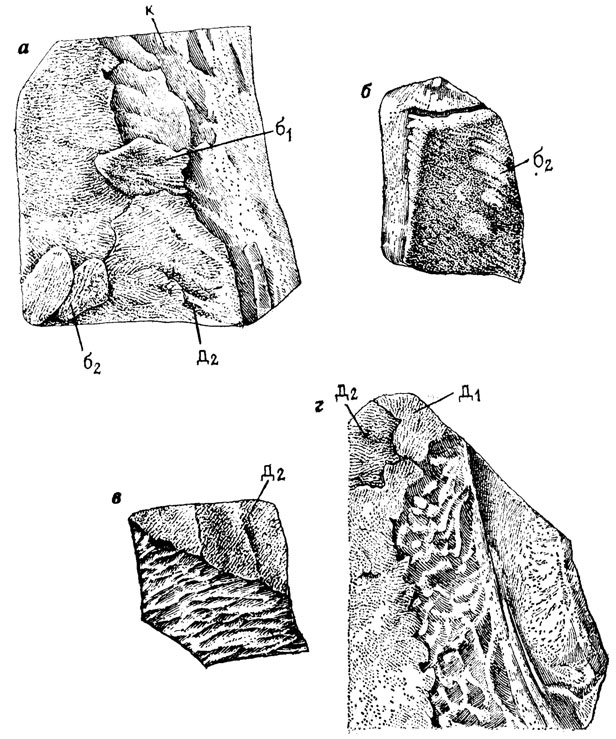
Рис. 74. Развитие и смена паркета дентина. По Э. И. Воробьевой. а - на фрагменте кости Thysanolepis: б1, б2 - блистеры, д1, д2 - старый и более новый паркет, к - кость; б - чешуя остеолепид с 'блистерами' - б2; в - залегание молодого косминового паркета (д2) на поверхностном костном слое у остеолепид; г - захождение молодого косминового паркета под старый у остеолепид
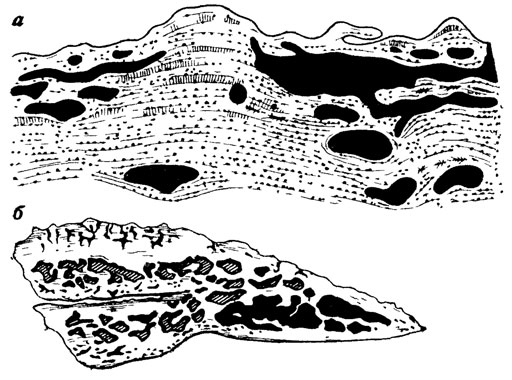
Рис. 75. Разные типы смены трабекулярного костного слоя на челюсти кистеперой рыбы Jarvikina. По Э. И. Воробьевой. а - 'шелушение'; б - расслаивание кости
Здесь мы встречаемся, очевидно, с параллелизмами в развитии покровных скелетных образований у разных групп позвоночных и со сходными явлениями "переноса" приложения морфогенетических процессов с космина на кость и далее вглубь с последовательным увеличением масштаба самого процесса; распространением с поверхностного костного слоя на глубинные мезенхимные и эктомезенхимные по происхождению слои в результате индуктивных взаимодействий между их клетками.
У амфибий (стегоцефалов) поверхностный слой кожных костей состоит из грубоволокнистой костной ткани зонарной структуры, а в губчатом и базальном слоях появляются вторичные остеоны и типичная пластинчатая кость. А. В. Румянцев [894] рассматривает строение экзоскелета амфибий и рептилий как особую стадию гистогенеза.
В основании боковой ветви рассматриваемого эволюционного ствола находятся актиноптеригии, которые выводятся обычно от общих предков с кистеперыми [894]. Исходной для древних костных рыб признана ромбическая чешуя с дентиновыми бугорками на поверхности (см. рис. 74, б). Чешуя такого типа дивергировала в космоидную, характерную для остеолепидных и поролепидных кистеперых и дипной, и ганоидную, характерную для лучеперых рыб, в которой постелено появлялись все более мощные слои ганоина (рис. 76).
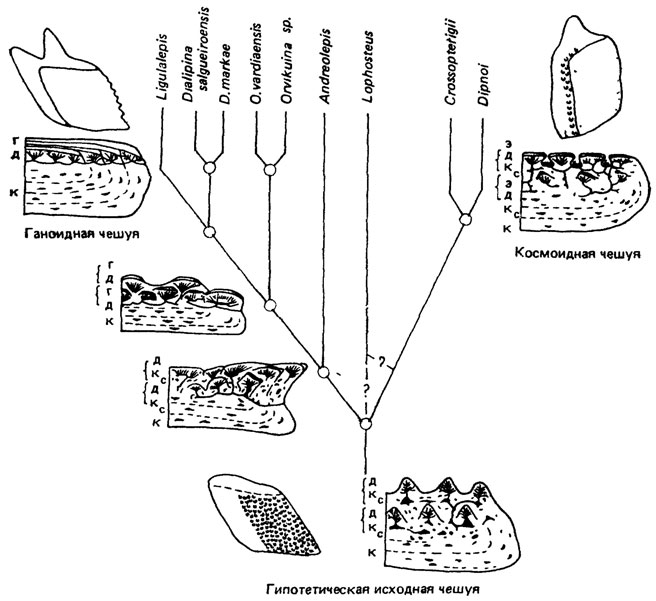
Рис. 76. Схема образования космоидной и ганоидной чешуи. По Г.-П. Шультце. к - костная ткань: кс - космин; д - дентин; г - ганоин; э - эмалеподобная ткань
В дальнейшем развитии ганоидных чешуи редуцируется дентин, а ганоин располагается непосредственно на кости. Следующая стадия отличается постепенной утратой ганоина, связанной с сильной редукцией костного слоя. Это приводит к образованию нового типа округлых чешуи, значительно отличающихся от ромбических ганоидных [2245] и состоящих исключительно из грубо-волокнистой костной ткани (например, циклоидные и ктеноидные чешуи телеостей).
Костные чешуи и дермальные кости у современных рыб в основном, состоят из костной ткани - продукта деятельности остеобластов, гистогенез которой идет по типу интрамембранной оссификации.
Среди рецентных образований особого внимания заслуживает микроструктура кожного скелета многоперов, используемая в качестве модели гистогенетических механизмов, проявляющихся в дивергентной эволюции дермоскелета всех костных рыб [1947]. Согласно Ф. Ж. Менье [1947], чешуя многоперых сходна с чешуей древних лучеперых (палеонисков) и, как и у дермальных костей, состоит из трех слоев. Дермальные кости костных рыб образованы костными тканями и их производными (различными вариациями изопедина и перманентной предкостной тканью).
Эндоскелет и его происхождение. Под эндоскелетом понимаются все внутренние скелетные образования - хорда, хрящи, мембранозные и преформированные хрящем кости. К мембранозным костям относят все кости эндоскелета, в том числе и гомологичные дермальным.
Согласно К. Гегенбауру [1630], хрящ первоначально появляется в эндоскелете, а кость - несколько позже в экзоскелете. В ходе эволюции кости экзоскелета погружаются в эндоскелет, приходят в соприкосновение с хрящем и начинают его вытеснять, что в конечном итоге приводит к образованию замещающих костей.
Неонтологи [1512] и палеонтологи [1784, 2502], работавшие в те же годы, что и К. Гегенбаур, выдвинули предположение об упрощении костного эндоскелета путем его редукции и замещения хрящевым. Это представление легло в основу работ Э. А. Стенше [2328, 2329], полагавшего, что кость и хрящ в эндоскелете являются одинаково древними и что в эволюции имело место вытеснение в эндоскелете кости хрящем. А. С. Ромер [2174], напротив, считал хрящ эмбриональным приспособлением, а сохранение его у взрослых - примером неотении.
Современники Э. А. Стенше [925, 926, 931, 1318] были сторонниками взглядов К. Гегенбаура. Правильность предположения последнего о возможности погружения дермальных костей в филогенезе в эндоскелете подтверждалась палеонтологическими находками. Как это ни парадоксально, но получены они были самим Э. А. Стенше [2328]г изучившим редукцию дермального скелета у немакропеталихтиидных артродир. Позже он гомологизировал дермальные кости плакодерм с эндоскелетными костями костных рыб [2330].
Теория К. Гегенбаура о происхождении костного эндоскелета была возрождена и дополнена А. В. Румянцевым [894], который, не отрицая возможности вхождения отдельных покровных костей в эндоскелет, полагал, что преформированные хрящем кости возникли в результате активного перемещения поверхностной скелетогенной мезенхимы внутрь с дальнейшим распространением ее в хрящевой ткани.
Сравнительный анализ ископаемых материалов по первичноводным позвоночным дает возможность прежде всего выявить основные соотношения хрящевой и костной тканей в экзо- и эндоскелете низших позвоночных [185]. Есть основания утверждать, что развитие тканей в эндоскелете, несмотря на его тесную связь с экзоскелетом, протекало в известной мере самостоятельно. Источник такой самостоятельности отмечался еще А. В. Румянцевым: "Можно допустить, что из скелеточной мезенхимы выделялись две нерезко отграниченные камбиальности: хрящевая и костная", т. е. эндо- и экзоскелетные системы" [894, с. 231].
Эволюция эндоскелета у ископаемых позвоночных. Эволюция скелетных тканей в экзо- и эндоскелетах происходила асинхронно и мозаично в разных группах. Однако у низших позвоночных прослеживаются и некоторые общие тенденции. К числу их относится преобладание хрящевой ткани в эндоскелете древних форм и постепенное замещение ее костью у геологически более поздних представителей тех же групп, что свидетельствует в пользу первичности хряща в эндоскелете позвоночных. Так, у большой группы бесчелюстных, гетеростраков, предполагаемых предков челюстноротых [752], мозговая коробка вместе с нерасчлененным висцеральным скелетом и посткраниальный скелет были образованы исключительно хрящом (см. рис. 71). Полностью хрящевым был эндоскелет у большинства панцирных рыб. Некоторые исследователи полагают даже, что хрящ эндоскелета имел более длительную историю, чем кость экзоскелета, т. е. древние позвоночные не имели дермальных окостенений. Была выдвинута гипотеза [1619], согласно которой скелетные ткани первоначально несли сенсорную функцию, формируясь в голове провертебрат вокруг органов чувств, и лишь вторично приобрели опорную функцию.
Древнейшая (из ордовика) находка хряща описана Т. Эрвигом [2033] как принадлежащая гетеростракам. Л. Б. Халстид (Тарло) в 1968 г. обратил внимание на сходство этой ткани с глобулярным обызвествленным хрящем у современных акул [2359]. Он классифицировал ее как хрящ со сферической кальцификацией полагая, что это древнейшая скелетная ткань позвоночных. Преформированная хрящом кость в эндоскелете древних бесчелюстных (остеострак) и артродир появляется первоначально в виде тонкой перихондральной выстилки поверхностей мозговой коробки и некоторых элементов позвонков, а также по ходу каналов, проникающих в хрящ черепа и посткраниального скелета рыб. Эндохондральная оссификация эволюционно рассматривается как более позднее образование [1973]. Так, у большинства древних силурийских акантодов мозговая коробка сплошь покрыта перихондральной выстилкой. У геологически более поздних каменноугольных форм череп состоит из серии окостенений, разделенных хрящом (например, у рода Acanthades). Наиболее рано окостеневает у них висцеральный скелет и эндоскелет плечевого пояса. Акантодов иногда приводят как пример группы, где редукция экзоскелета у поздних представителей компенсируется усилением окостенений эндоскелета.
В ряду костных рыб (двоякодышащих и древних лучеперых - палеониксов и кистеперых) к середине палеозоя (в позднем девоне) череп состоит из монолитных двух-трех окостенений. Хряща в черепе очень мало, он представлен лишь в области контакта этих окостенений (например, в ушной и затылочной фонтанели у палеонисков) и локально - в носовой капсуле у кистеперых рыб [186]. Висцеральный скелет также, как правило, окостеневает. Процесс окостенения в посткраниальном скелете у позвоночных обычно отстает от окостенения черепа. Например, у поздних акантодов в осевом скелете преобладает хрящ (слабое окостенение невральных дуг и позвонков). На более поздних этапах эволюции у костных рыб, за исключением рецентных хондростей, отмечается усиление окостенения посткраниального скелета, протекающее независимо в разных линиях.
В более поздней истории первичноводных и низших наземных позвоночных наряду с прогрессирующей оссификацией имело место также вторичное увеличение хряща в скелете. Количественные соотношения кости и хряща могли быть различными, и в целом увеличение хряща в эндоскелете носило мозаичный характер. Например, главные отличия древнейших тетрапод от рипидистных кистеперых состоят в слиянии черепных блоков и наличии охрящеваний в этмоиде. В конце палеозоя у амфибий начинает преобладать хрящ в черепе, что, однако, сочетается у них с обычным присутствием костных позвонков. У целакантов в карбоне - перми в отличие от девонских их представителей с двублочным костным эндокранием последний в значительной степени становится хрящевым с отдельными окостенениями; позвоночник остается хрящевым (костные позвонки найдены лишь в хвостовой области у древних форм). Начиная с перми, у этих рыб помимо окостеневших невральных и гемальных дуг встречаются также костные ребра. Однако у целакантов-лаугиид, напротив в раннем триасе отмечаются окостенения мозговой коробки в области межглазничной перегородки и отико-окципитального отдела.
Сходство с целакантами в общих тенденциях к раннему появлению кости, а затем к повторному увеличению количества хряща на поздних этапах филогенеза обнаруживают также дипнои. Сравнительно полное окостенение черепа в девоне сменяется у них постепенно на уровне современных видов столь же полным охрящеванием: в черепе сохраняются только две преформированные хрящом кости в ушной области; посткраниальный скелет уже начиная с триаса преимущественно хрящевой, тогда как девонские дипнои имели массивные костные кольцевидные позвонки.
У многих лучеперых рыб, за исключением рецентных хондростей, наблюдается обратный процесс; начиная с мезозоя, прослеживается увеличение костной ткани в осевом скелете и хряща - в эндокрании. Хрящ преобладает в эндоскелете у глубоководных кистеперых рыб. По сравнению с палеонисками хрящ преобладает у них в этмоидной области и исчезает между окостенениями ушного и затылочного сегментов, вступающих в шовные соединения. На некоторых участках черепа на месте хряща развиваются соединительно-тканные мембраны (межглазничная перегородка) и эндоскелет вытесняется дермальными костями (ушная область и крышка черепа).
Многие морфологи [1319, 1797, 1958, 2174, 2208] склоняются к восприятию хряща как эмбриональной адаптации, обеспечивающей в силу пластичности наиболее благоприятные условия роста. Сохранение хряща дефинитивно, а также вторичное увеличение количества хряща в эволюции разных групп расценивается с этих позиций как отрицательная анаболия тканей [185, 813J в эндоскелете, что связывается с водным образом жизни анамний. Однако попытка отыскать адаптивный смысл этого явления - связь с механическими нагрузками, равновесием, плавучестью [2208] - мало что дает для выявления самого механизма и его причин. Связь с водным образом жизни может быть здесь чисто морфогенетической.
Процесс гистологической "эмбрионализации" путем отрицательной анаболии мог прогрессировать в ходе эволюции (с охватом ранних стадий онтогенеза), что имеет место, по-видимому, у дипной, целакантов, древних лучеперых и амфибий. Однако это отнюдь не сопровождалось превращением черепа анамний в эмбриональный. Процессы кажущейся "хондрификации" у дипной и хрящевых ганоидов при отсутствии у них настоящей энхондральной оссификации, с одной стороны, и у целакантов и амфибий при наличии у них типичной энхондральной оссификации [2073], с другой, - явления различной природы.
Таким образом, можно заключить, что хрящ и кость в эндоскелете позвоночных с самых первых этапов их эволюции образуют целостную и динамичную тканевую систему, коррелятивно связанную с экзоскелетом и обладающую определенной автономностью и способностью к самоорганизации. Развитие этой системы в процессе эволюции первичноводных позвоночных идет дивергентной мозаично по пути последовательного замещения хрящевой ткани костной, обеспечивающей общее упрочнение эндоскелета, что нередко сопровождается редукцией экзоскелета.
Критический анализ сведений по микроструктуре хряща и преформированных хрящом костей у ископаемых позвоночных [185, 1690, 2033, 2328] позволяет отметить не только тенденции в усложнении структуры этих образований, но также и некоторые параллелизмы в рядах позвоночных.
| Формы | Состав эндоскелета |
|---|---|
| Antiarchi | Хрящ [185] |
| Первые Acanthodii, некоторые Arthrodira | Обызвествленный хрящ [185] |
| Поздние Acanthodii, Arthrodira | Перихондральная костная манжетка [2328, 185] |
| Chondrichthyes | Обызвествленный хрящ [2033] |
| Amphibia | Костная ткань энхондрального происхождения [42, 1691] |
| Reptilia | Перихондральная костная манжетка и костная ткань энхондрального происхождения [2162, 2163] |
Оссификация эндоскелета, как и формирование сходных типов микростроения зубов, происходит параллельно в разных филумах, демонстрируя, таким образом, принцип параллелизма дивергентных изменений гистологических структур В. П. Михайлова и А. А. Брауна [116]. Увеличение хряща в эндоскелете, прослеживающееся на более поздних этапах эволюции ряда групп позвоночных, ведет к кажущейся обратимости тканевых преобразований, отражая вместе с тем специфичность тканевого уровня. Осуществляясь по принципу отрицательной анаболии, эта "хондрификация" может рассматриваться как своего рода деспециализация, которая открывает новые пути и возможности для дальнейших преобразований отдельных структур в эволюции [185]. Асинхронность и мозаичность переходов в близких филумах (от преобладания хрящевой ткани в скелете к преобладанию костной ткани) и в то же время параллелизмы в этих процессах между разными группами, а также независимое достижение сходных структурных состояний в экзо- и эндоскелете как в близких, так и в филетически удаленных линиях - все вместе взятое заставляет с большой осторожностью относиться к попыткам создания естественной системы хрящевой и костной тканей в рядах позвоночных. Более того, как показывают наблюдения над эволюцией экзо- и эндоскелетных тканей среди первичноводных позвоночных [186, 189], использование особенностей микростроения скелетных тканей при реконструкции филогенезов и диагностике возможно лишь в сочетании с другими морфологическими признаками, доказывающими таксономическую принадлежность находок.
Можно выделить несколько путей формирования элементов костного эндоскелета [815].
1. Погружение дермальных костей внутрь тела [1630, 2328, 2330].
2. Образование костных тел позвонков вокруг хорды:
а) без участия хрящевой ткани - у костных рыб и некоторых амфибий [114] (Считалось [1948], что в данном случае имеет место выпадение хрящевой стадии. По мнению В. Г. Борхвардта [114], образование перихондральных костных манжеток без участия хряща у костных рыб и некоторых амфибий первично);
б) с участием хрящевой ткани - у всех тетрапод, за исключением некоторых амфибий.
3. Образование перихондральных костных манжеток вокруг хрящевых моделей, в результате чего возникли преформированные хрящом кости:
а) без настоящей энхондральной оссификации у хрящевых ганоидов и двоякодышащих (Р. У. Хайнс [1691] считает этот способ вторичным. По В. П. Пегете [814], это явление первично);
б) с последующим замещением хрящевой ткани костной, типичной энхондральной оссификацией, но без костномозгового кроветворения - у костных ганоидов, костистых рыб, многоперых, кистеперых и некоторых амфибий;
в) с энхондральной оссификацией и костномозговым кроветворением - у некоторых амфибий и всех амниот.
Не преформированные хрящом кости эндоскелета всех позвоночных вне зависимости от их положения в системе развиваются очень сходно, на что в свое время обратил внимание еще А. Келликер [1840]. Способ развития мембранных костей получил название интрамембранной оссификации [1129].
В местах, где в будущем будет развиваться кость, обособляются остеогенные участки мезенхимы, что выражается в увеличении числа и толщины коллагеновых волокон. Мезенхимные клетки оттесняются волокнами к периферии. Эти клетки в эта время имеют звездчатую или веретеновидную форму. Затем они округляются, а их отростки утолщаются и соединяются с подобными отростками соседних клеток. Таким путем остеогенные клетки превращаются в остеобласты, которые продуцируют основное вещество костной ткани. В клеточной костной ткани всех позвоночных остеобласты окружаются органическим матриксом костной ткани и превращаются в остеоциты. В очень молодой бесклеточной костной ткани некоторых костистых рыб остеобласты могут также замуровываться, приобретая сходство с остеоцитами клеточной кости [1972], В дальнейшем развитие клеточной и бесклеточной костей идет за счет надкостницы (периоста). В результате периостального остеогенеза образуется грубо-волокнистая костная ткань [2462; см. табл. 6]. В клеточных костях остеобласты замуровываются органическим матриксом костной ткани, а в бесклеточной кости в этот момент остеобласты исчезают. Заметить разрушение остеобластов очень трудно и поэтому создается впечатление, что остеобласты периоста "отступают" от бесклеточного костного матрикса [1972]. Возможно, что таким путем формировался аспидин у агнат.
Образовавшаяся грубоволокнистая костная ткань у позвоночных может разрушаться, на ее месте формируется скорлуповая (пластинчатая) костная ткань [2462; см. табл. 6].
Развитие хряща и преформированных хрящом костей в онтогенезе у современных позвоночных. Попытки проследить усложнение микроструктуры преформированных хрящом костей предпринимались неоднократно [894, 1311, 1327, 1539, 1880].
Сравнительное исследование [2073] этих костей в ряду позвоночных (круглоротых, хрящевых рыб, хрящевых ганоидов, двоякодышащих, костистых рыб, амфибий, рептилий, птиц, метатерий и эутерий) позволило выявить усложнения в их микроструктуре, которые происходят путем анаболии и девиации (рис. 77).
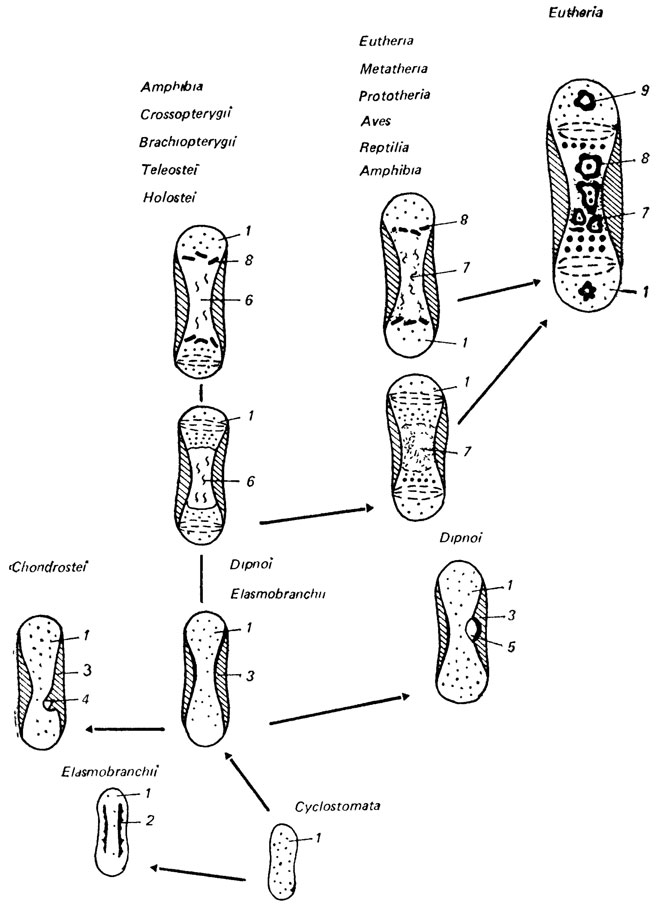
Рис. 77. Усложнение микроструктуры преформированных хрящом костей в онтогенезе современных позвоночных. По В. П. Пегете. 1 - хрящевая ткань; 2 - локальная кальцификация хряща; 3 - перихондральная костная манжетка; 4 - примитивная энхондральная оссификация; 5 - локальная резорбция хряща; 6 - медуллярная полость; 7 - миелоидная ткань; 8 - трабекулы энхондральной костной ткани; 9 - вторичный центр оссификации
Эндоскелет современных круглоротых в течение всей жизни остается хрящевым. Хрящ круглоротых богат хондроцитами и беден основным веществом. В онтогенезе хрящевых рыб элементы эндоскелета проходят две стадии: хрящевой закладки и локальной кальцификации хряща. Хрящ хрящевых рыб богат основным веществом. Перихондральные костные манжетки описаны в нейральных дугах собачьей акулы [2074].
У двоякодышащих (лепидосирен и неоцератод) преформированные хрящом кости проходят также две стадии - хрящевой закладки и перихондральной костной манжетки, а у протоптеруса еще и третью стадию - локальной резорбции хряща.
У хрящевых ганоидов различают три стадии: хрящевой закладки, перихондральной костной манжетки и примитивной энхондральной оссификации (разновидность периостальной). При примитивной энхондральной оссификации костная ткань образуется вокруг каналов, заходящих из перихондральной костной манжетки в хрящ, преформировавший кость [812]. Хрящ при этом не теряет своей непрерывности по всей длине кости. Образование костной ткани по ходу каналов, проникающих в хрящ, было отмечено ранее у ископаемых агнат и гнатостомат [2328, 2329].
Преформированные хрящом кости костистых рыб в онтогенезе проходят четыре стадии: 1) хрящевой закладки; 2) перихондральной костной манжетки; 3) образования медуллярных полостей вследствие резорбции хряща; 4) энхондральной оссификации. У мелких представителей костистых рыб описаны только первые две стадии [1691].
Первая и вторая стадии сходны у костных ганоидов и многоперых, а также у двоякодышащих, хрящевых ганоидов и костистых рыб, а дефинитивная микроструктура этих костей у костных ганоидов, многоперых и кистеперых (латимерия) соответствует морфологическим характеристикам четвертой стадии костистых рыб. По-видимому, преформированные хрящом кости упомянутых рыб в онтогенезе также проходят четыре стадии.
У амфибий известны два способа развития преформированных хрящом костей. Древний, такой же как и у костистых рыб (по способу анаболии), и более прогрессивный. При втором способе в образующейся медуллярной полости на третьей стадии (девиации) появляется кроветворный костный мозг, который сохраняется и на четвертой стадии энхондральной оссификации.
Амниоты унаследовали последний способ развития костей от амфибиеобразных предков. Их кости в онтогенезе проходят стадии хрящевой закладки, перихондральной костной манжетки, образования медуллярных полостей вследствие резорбции хряща и начала миелогенеза, энхондральной оссификации*. Третья и четвертая стадия у большинства Eutheria начинаются почти одновременно.
* (В. П. Пегета и М. В. Бевольская показали, что в онтогенезе длинных костей нанду энхондральная оссификация предшествует миелогенезу.)
При сравнении конечных стадий развития преформированных хрящом костей в ряду современных позвоночных обнаруживаются такие же закономерности в усложнении их дефинитивной микроструктуры, как и в их онтогенезе [2073]:

Перихондральная (соответственно периостальная) часть преформированных хрящом костей у всех позвоночных независимо от их положения в системе развивается удивительно сходно (по типу интрамембранной оссификации), на что обращали внимание уже давно [1840]. Сравнение стадий онтогенеза преформированных хрящом костей с дефинитивной микроструктурой хряща у современных и ископаемых позвоночных позволяет утверждать, что закономерности усложнения их микроструктуры в онто- и филогенезе идентичны [2073].
Эндохондральная оссификация проявляется в образовании трабекул костной ткани на остатках резорбированного хряща, преформировавшего кость, причем у низших и у высших позвоночных костная ткань эндохондрального происхождения всегда тонковолокнистая.
Усложнение микроструктуры посткраниального осевого скелета. В осевом скелете позвоночных представлены элементы различного происхождения. Преформированные хрящом кости черепа, нейральные и гемальные дуги позвоночника формируются таким же путем, как и другие скелетные элементы, развивающиеся на хрящевой основе.
В развитии же тел позвонков имеются особенности, связанные с наличием хорды. Но хорда, как и необызвествленный хрящ, в ископаемом состоянии не сохраняется. Поэтому отношения между хордой и хрящевыми либо костными элементами осевого скелета реально прослеживаются только на неонтологическом материале [113, 114].
У современных бесчерепных и круглоротых хорда сохраняется в течение всей жизни и несет хрящевые нейральные дуги. Среди современных эласмобранхий известны формы, у которых нет тел позвонков (наиболее примитивные), и формы с хрящевыми телами позвонков [114]. У современных эласмобранхий (кладодонтиды и хибодонтиды) осевой скелет представлен также исключительно хордой [2210].
У хрящевых ганоидов и современных двоякодышащих тела позвонков отсутствуют. В то же время у ископаемых двоякодышащих имелись хорошо развитые тела позвонков в виде костных амфицельных дисков с маленькими отверстиями для хорды или без них [194]. Г. П. Шультце в 1971 г. пытался найти хрящевую ткань в телах позвонков ископаемых двоякодышащих. Т. С. Вестолл и С. М. Эндрюс [1276] полагают, что хрящевые элементы могли присутствовать в позвонках многих ископаемых кистеперых рыб. В то же время у современного представителя кистеперых (латимерии) тел позвонков нет.
По представлениям В. Г. Борхвардта [113], тела позвонков в эволюции разных групп костных рыб (за исключением панцирной щуки) возникали сразу костными, причем хрящевая стадия полностью отсутствовала на всех стадиях онтогенеза. По его мнению, хрящевая стадия появляется только у тетрапод.
Проведенная ревизия структуры позвонков у ископаемых рипидистных кистеперых [195] дает возможность предполагать, что морфогенез позвонков у древних рыб отличается большой пластичностью и их формирование происходило, согласно гипотезе Ж. Лаерма [1861], под значительным влиянием механических нагрузок, связанных с локомоцией в водной среде. С переходом к тетраподам происходила принципиальная перестройка позвоночника, которая могла идти по пути значительных изменений в онтогенезе. Вместе с тем механизм взаимоотношений между хрящевой, костной и миелоидной тканями в развитии элементов позвонков на уровне амниот принципиально, по-видимому, не отличался. Поэтому усложнение микроструктуры в телах позвонков тетрапод в онто- и филогенезе могло происходить путем анаболии и девиаций, по А. Н. Северцову.
Гистоструктура и развитие скорлупы яиц. Твердая скорлупа яиц зауропсид слагается карбонатом кальция: у птиц, динозавров, крокодилов, чешуйчатых и ринхоцефалов - в форме кальцита; у черепах - в форме арагонита; в малых количествах присутствует и карбонат магния [1344, 2400, 2062]. Минеральная составляющая скорлупы пронизана сетью органического матрикса, имеющего мукополисахаридно-протеиновый состав [1561, 1960, 2400].
В строении скорлупы яиц на радиальном (поперечном) сечении в зависимости от методов изучения традиционно подчеркивается либо ее горизонтальная (параллельно линиям прироста), либо вертикальная (вдоль направления прироста) структурированность (рис. 78). При исследовании в световом неполяризационном и сканирующем электронном микроскопах скорлупа яиц птиц (модельный объект) отчетливо делится на горизонтальные слои (см. рис. 78, а, в) - сосочковый (нижний) и губчатый (верхний). Губчатый слой выявляется в той или иной степени как морфологически "целое" образование; сосочковый слой состоит из отчетливых отдельностей (сосочков, или конусов), слагаемых радиирующими субъединицами - "клиньями" (см. рис. 78, г). При исследовании шлифов в поляризованном свете скорлупа яиц подразделяется на вертикальные единицы - экзосфериты, внутри которых выявляются субъединицы, одновременно затухающие при вращении столика микроскопа в скрещенных николях; горизонтальная стратификация скорлупы, напротив, совершенно не прослеживается (см. рис. 78, б).
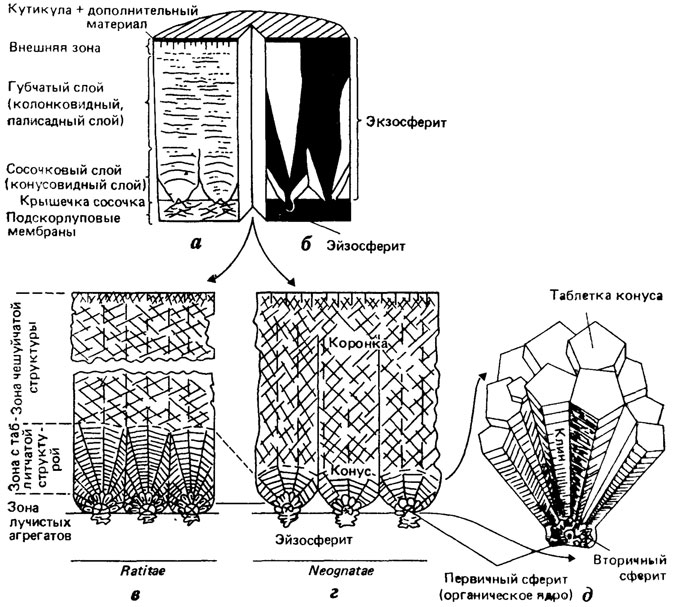
Рис. 78. Микроструктура скорлупы яиц. По X. К. Эрбену с изменениями. Строение скорлупы яиц птиц в радиальном сечении под оптическим микроскопом: а - в неполяризованном свете; б - в поляризованном свете; в, г - схема микроструктуры скорлупы яиц бескилевых и килевых птиц по данным СЭМ; д - трехмерное изображение структуры сосочка (крышечки сосочка и конуса); внутри крышечки сосочка - первичный сферит, окруженный лучистыми кристаллитами, горизонтальная исчерченность клиньев означает слои прироста; в пределах клина субмикроскопические элементы достраиваются до многоугольных таблеток
Эти два аспекта структурированности скорлупы, строго говоря, отражают разные физические свойства объекта и говорят о двойственности его природы. Выявляемая поляризационными и дифракционными методами кристаллография материала скорлупы яиц не связана с морфологическими особенностями биогенного минералообразования и потому малоинформативна для изучения скорлупы в таксономическом и филогенетическом аспектах. Напротив, горизонтальная стратификация скорлупы на зоны (слои) с разными типами структуры, обнаруживаемая в СЭМ, отражает биологически детерминируемые особенности минерализации. Границы между структурными зонами свидетельствуют о перерывах в минерализации; отсутствие преемственности в микроструктуре и макроструктуре слагающего скорлупу материала (по обе стороны от границы) - об изменении характера минерализации. Эти изменения связаны с функциональной дифференциацией секретирующего эпителия яйцевода: формирование новой структурной зоны обусловлено либо перемещением яйца в другой отдел яйцевода, либо цитобиохимическими изменениями в функционировании секретирующего эпителия одного и того же отдела. Наиболее сложно строение скорлупы яиц птиц. По мере роста скорлупы образуются три структурные зоны: с лучисто-радиальной структурой (в основании сосочка), с таблитчатой структурой (собственно сосочковый слой) и с чешуйчатой структурой (губчатый слой [1561, 1960]). Иногда выше губчатого слоя может формироваться зона с вертикально-пластинчатой структурой (внешняя зона или внешний кристаллический слой). Выше внешней зоны могут откладываться слои органического или смешанного состава [1344], однако они утрачиваются в процессе фоссилизации и не могут быть обнаружены на ископаемом материале. Лучисто-радиальная структура слагается игольчато-призматическими кристаллами кальцита, радиирующими от поверхности органических ядер - крупных сферических тел, во множестве откладываемых на поверхности подскорлуповой оболочки яйца в терминальной части "перешейка" яйцевода. Ниже органических ядер появляются эйзосфериты - рыхлые агрегаты кристаллических пластин, формирующихся между волокон внешней подскорлуповой оболочки яйца. Дискретная минерализация верхних слоев подскорлуповой оболочки, возникавшая, вероятно, независимо в разных группах зауропсид, может рассматриваться как начальная стадия в эволюции твердой скорлупы яиц у архозавров, чешуйчатых и ринхоцефалов (рис. 79).
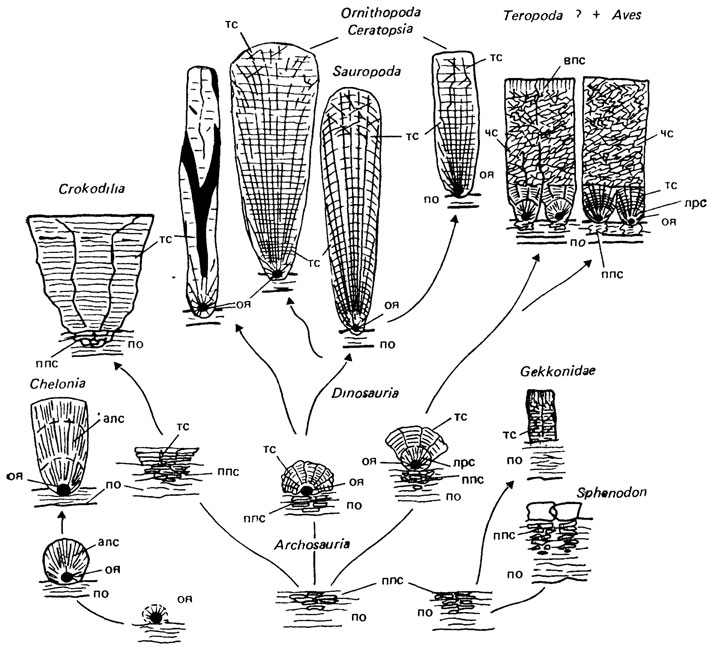
Рис. 79. Эволюционные преобразования в структуре скорлуповой оболочки яйца у зауропсид (тонким контуром показаны гипотетические стадии). К. Е. Михайлов, оригинал. Структуры: алс - арагонитовая лучистая; впс - вертикально-пластинчатая; лрс - лучисто-радиальная (кальцитовая); ппс - первичная пластинчатая; тс - таблитчатая; чс - чешуйчатая (слагает губчатый слой); оя - органическое ядро; по - подскорлуповая оболочка (кожистая оболочка у лепидозавров)
Собственно сосочковый и губчатый слои (большая часть скорлупы) формируются у птиц уже в следующем за "перешейком" отделе яйцевода - "скорлуповой железе". Таблитчатая структура, слагающая "клинья" сосочка, представлена плотно спрессованными стопками пластинчатых агрегатов кальцита (см. рис. 78, г). Губчатый слой, сложенный чешуйчатой структурой, представляет собой совершенно новое образование в эволюции скорлуповой оболочки яйца. Кроме скорлупы яиц птиц он обнаружен в скорлупе яиц некоторых меловых динозавров (вероятно, теропод). "Орнитоидное" строение скорлупы яиц, свойственное птицам и тероподам, характеризует наиболее прогрессивную из трех линий филогенеза скорлуповой оболочки яйца у архозавров. Совершенно особый тип строения скорлупы яиц характерен для зауропод, орнитопод и цератопсов. У этих динозавров вся скорлупа представлена одной структурной зоной (с таблитчатой структурой), формирующейся непосредственно выше органического ядра. Вертикальная отдельность скорлупы морфологически представляет собой здесь веер тонких радиирующих "клиньев" (эта зона целиком аналогична зоне "клиньев" в скорлупе яиц птиц). В пределах этой линии, филогенеза скорлуповой оболочки яйца разным группам динозавров свойственны разные варианты строения скорлупы, отличающиеся морфологическими особенностями экзосферитов и строением поровых каналов [1003, 1004].
Совершенно особое строение скорлупы яиц (третья линия филогенеза скорлуповой оболочки у архозавров) характерно для крокодилов. Крупные массивные "клинья" также сложены здесь исключительно таблитчатой структурой; отсутствует не только зона с лучисто-радиальной структурой, но и органическое ядро [1561]. Независимо от архозавровой линии зауропсид появилась скорлупа яиц у черепах: она сложена арагонитом и не имеет гомологичных архозавровым типам строения гомологичных элементов кристаллической структуры. Особые типы строения скорлупы яиц обнаружены у ринхоцефалов (гаттерия) и гекконов [2062].
Значительный интерес представляет многослойная скорлупа яиц динозавров из верхнемеловых отложений в Южной Франции. Строение этой скорлупы рассматривалось как патология, повлиявшая на исчезновение обитавших в меловое время на территории Южной Франции динозавров [1559, 1561]. В последнее время многослойную скорлупу яиц не связывают с проблемой вымирания. Подобные патологии известны у черепах и вызываются задержкой кладки яиц в яйцеводе до следующего цикла овуляции.
Не все рептилии откладывают яйца с твердой скорлупой. Яйца ящериц (исключая гекконов, гаттерию), змей и, вероятно, ряда ископаемых групп имеют (имели) лишь кожистую оболочку с отдельными группами кристаллов кальцита (или без них) на поверхности. Наиболее древние из ископаемых яиц рептилий (вероятно, текодонтов), найденные в верхнем триасе Африки, также, видимо, имели кожистую, эластичную оболочку.
Исследование гистогенезов твердых тканей экзоскелета у представителей разных групп позвоночных имеет большое значение для их классификации, систематики и использования в биостратиграфии. Во избежание ошибок важно не только установить степень родства между группами, но и проследить направленность и закономерности развития гистологических особенностей эволюции. Результаты изучения экзоскелета позвоночных все более широко применяют в геологической практике. Использование мелкофрагментного материала для корреляции отложений (в особенности континентальных) с каждым годом все более расширяется. Особенно успешно в стратиграфии используют элементы экзоскелета (чешуя, зубы, дискретные части панциря, фрагменты костей таких групп, как телодонты, гетеростраки, акантоды, кистеперые, стегоцералы, ископаемые рептилии).
Исключительно важную роль играют микроостатки экзоскелета позвоночных при расчленении и корреляции разнофациальных раннепалеозойских отложений (в первую очередь, прибрежно-морских, дельтовых и лагунных). При помощи химической препаровки из этих отложений удается получить массовый и разнообразный материал фосфатного состава. В процессе его обработки, как правило, ведутся микроструктурные исследования и на их основе определяется таксономическая принадлежность остатков. Большое значение при этом имеет выяснение принципов роста кожных окостенений у разных групп низших позвоночных. Если остатки экзоскелета (например, чешуи бесчелюстных и рыб) представлены разными возрастными стадиями, то удается проследить их онтогенетическое развитие. В частности, удается восстановить те изменения, которые происходили с момента закладки чешуи в кориуме до ее дефинитивного состояния. Это дает возможность установить тип роста чешуи, связав его с систематическим положением. Исследования такого направления в сочетании с морфологическими и учетом топографической изменчивости чешуйного покрова позволяют приблизить искусственную классификацию, разработанную на основе дискретных частей экзоскелета, к естественной.
Правильное определение родовой и видовой принадлежности микроостатков дает возможность успешно применять их при выделении дробных стратиграфических подразделений и разработке зональной стратиграфии.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'