
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
3.2. Основные концепции и проблемы палеоихнологии (М. А. Федонкин)
Палеоихнология (от греческ. ichnos - след) - часть палеонтологии, изучающая ископаемые следы жизнедеятельности организмов. От других областей палеонтологии она отличается характером объектов исследований. Особенность последних состоит в том, что они являются результатом непосредственного взаимодействия организмов и внешней среды. Пожалуй, ни одна группа окаменелостей не дает столь ярких примеров теснейших и многообразных связей организмов и среды их обитания, какими богата палеоихнология.
Первые описания ископаемых следов жизнедеятельности организмов появились в начале прошлого века, однако статус самостоятельной дисциплины палеоихнология получила лишь 25-30 лет назад. Долгое время многие ископаемые следы организмов описывались как остатки морских водорослей. Об этом свидетельствуют такие наименования следов, как Algacites, Fucoides, Chondrites и многочисленные их названия, оканчивающиеся на - phycus. Лишь неоихнологические и актуопалеонтологические наблюдения, которые начали проводиться в конце прошлого века, развеяли это заблуждение и открыли истинную природу многих подобных окаменелостей.
В современную эпоху палеоихнология концентрирует внимание на исследованиях следов жизнедеятельности беспозвоночных животных, что нашло отражение в публикации специального тома в рамках американской сводки "Treatise on invertebrate paleontology". Следы же позвоночных в данном случае выделяются в особую область, которой занимаются преимущественно не палеоихнологи, а специалисты по морфологии и систематике этих животных. Работы, специально посвященные следам жизнедеятельности позвоночных животных, относительно немногочисленны [201, 208, 210, 937, 1876 и др.].
Следы позвоночных довольно часто привязывают к определенным видам животных и не дают им особых наименований. В изучении же следов беспозвоночных организмов ситуация совершенно иная, рассмотрению которой главным образом и посвящена настоящая глава.
Традиционно палеоихнология рассматривает разнообразные ископаемые следы: покоя, передвижения по поверхности и в толще осадка; следы, связанные с питанием, образованием норок-жилищ беспозвоночных организмов и т. д. Кроме того, изучаются копролиты, которые часто прямо связаны со следами передвижения животных, а также следы литофагов и сверлильщиков в твердом субстрате, например, в каменном дне, гальках, раковинах и других скелетных образованиях. Своеобразие рассматриваемых окаменелостей заключается в следующем.
1. В подавляющем большинстве случаев следы жизнедеятельности находятся in situ. Редчайшие случаи перемещения следов организмов обычно легко диагностируются. Это качество весьма важно в палеоэкологических и стратиграфических исследованиях, так как прочие остатки организмов после гибели часто переносятся на значительное расстояние.
2. Следы часто находят в толщах, не содержащих никаких других органических остатков, что объясняется более высокой их способностью к сохранению (особенно тех, которые оставлены ниже поверхности осадка, например норки илоедов) по сравнению с бесскелетными организмами, составляющими важнейшую часть бентических сообществ.
3. Ископаемые следы крайне редко могут быть таксономически соотнесены с какими-либо видами сопутствующих окаменелостей (за исключением позвоночных и некоторых членистоногих). Таксономическая привязка следов часто невозможна даже с точностью до типа. Это объясняется тем, что различные группы животных нередко оставляют похожие следы, имея одинаковую, например червеобразную форму тела, и сходным образом реагируя на одинаковые воздействия среды или ведя одинаковый образ жизни. С другой стороны, одно и то же животное может оставлять существенно различающиеся следы на протяжении своего существования, например, из-за возрастных и (или) сезонных различий в образе жизни, в разных условиях среды и в различные фазы поведения (покоя, передвижения, питания и т. д.).
4. Форма следа в большинстве случаев отражает не столько морфологию животного, сколько функционирование его органов, его деятельность и поведение. Эта особенность следов является наиболее важным отличием их от других окаменелостей.
С большим интересом относятся к следам жизнедеятельности организмов седиментологи. Бентос играет заметную роль в разрушении и формировании структур и текстур, в изменении осадков вообще. Можно привести множество примеров, когда осадок полностью перерабатывается илоедами, а в абиссальных зонах океана, где скорость накопления осадков чрезвычайно мала, животные успевают несколько раз переработать осадок даже при относительно низкой плотности популяций. Седиментологи используют следы для определения относительных, а иногда и абсолютных скоростей накопления или эрозии осадка, направления и скорости течений, состояния грунта в момент образования следов [1646, 2139, 2261 и др.].
Возможности палеоихнологии в реконструировании условий среды особенно широки, поскольку следы отражают поведенческую реакцию животного на подвижность воды вблизи поверхности осадка, тип субстрата, наличие пищевых ресурсов и т. п. В общем случае с увеличением глубины уменьшается подвижность воды, осадок делается более тонким, причем доля органического детрита в его составе увеличивается. Благодаря чувствительности животных к условиям среды ископаемые следы могут быть хорошими экологическими индикаторами.
Устойчивые и общие для большинства бентосных животных реакции на среду позволили А. Зейлахеру [2266, 2268] создать модель батиметрической зональности следов. Эта модель (иногда ее называют ихнофациальной), основанная на изучении ископаемого материала, позже была подтверждена данными об экологии и поведении современной фауны и подводным фотографированием [1566, 1753]. А Зейлахер показал, что существует несколько типов ассоциаций ископаемых следов (ихнофаций), контролируемых глубиной бассейна и проходящих практически через весь фанерозой. Используя понятие ихноспектра, которое позволяет проводить качественное и количественное сравнение ихноценозов в соответствии с этологической классификацией следов [2261], можно видеть, как направленно меняется облик ихнофаций с ростом глубины. В мелководной зоне доминируют вертикальные трубки, разнообразные, часто глубокие норки-жилища и следы покоя. В глубоководной зоне обычны субгоризонтальные поверхностные следы проедания и пастьбы, имеющие сложную конфигурацию.
Однако, как свидетельствует опыт, модель батиметрической зональности следов нельзя механически применять для установления глубины древнего бассейна в данной точке. Это связано с тем, что батиметрические ассоциации следов или ихнофаций, предложенные А. Зейлахером, в значительной степени определяются факторами их сохранности. С удалением от берега и возрастанием глубины существенно увеличивается возможность сохраниться в ископаемом состоянии именно у приповерхностных следов, тогда как на мелководных участках дна эти следы чаще размываются. Кроме того, известны случаи совместного нахождения форм, характерных для разных ихнофаций. Иногда следы, типичные для более глубоководных зон бассейна, находят в мелководных фациях [1695]. Это происходит обычно в тех случаях, когда тонкий осадок, не возмущаясь волнами и течениями, накапливается вблизи береговой линии. Иллюстрацией сказанного могут служить находки Zoophycus в мелководных фациях [2056, 2057]. Ясно, что в большинстве случаев требуется комплексный подход к изучению среды древних бассейнов вне зависимости от того, решается ли частный вопрос или проводятся широкие палеоэкологические исследования. Блестящим примером такого подхода служат работы Р. Ф. Геккера и его соавторов [226, 232, 235 и др.].
Весьма велико и палеонтологическое значение ископаемых следов. Палеонтологи начинают понимать, что их традиционные объекты изучения - остатки организмов - не единственные свидетельства жизни (а часто - не столько жизни, сколько смерти). Подлинными свидетельствами жизни, и прежде всего жизнедеятельности во всем ее многообразии, являются следы, оставленные существовавшими когда-то организмами. По ним можно судить о количестве и разнообразии животных в пределах биотопа, о плотности и динамике популяций, о трофических явлениях и связях. По следам можем расшифровать способы передвижения животных, особенности их поведения в процессе питания и реакции на некоторые экологические факторы. Нередко имеется возможность получить также информацию о морфологии животных, уровне их физической, физиологической и даже психологической организации, а в ряде случаев - об их таксономической принадлежности.
Что касается докембрия, то вполне вероятно, что именно ископаемые следы позволят определить время появления первых бентосных Metazoa в геологической летописи и существенно дополнить историю освоения животным миром морского (а, возможно, и не только морского) дна как местообитания и экологической ниши.
Основное значение для верной интерпретации ископаемых следов имеют данные неоихнологии и актуопалеонтологии. Тем не менее палеоихнологи продолжают испытывать острую нехватку данных о следах современных животных, их образе жизни и поведении. Это объясняется не только малым количеством актуопалеонтологических наблюдений, но и ограниченным числом доступных биотопов: большинство исследований касается самой мелководной прибрежной части моря - литорали. При этом нередко изучаются лишь поверхностные следы, а большая часть структур, образованных ниже поверхности осадка (эндогенных), не учитывается, так как приемы их исследований весьма трудоемки и требуют специального оборудования [1574, 1756].
Большинство ископаемых следов относятся именно к группе эндогенных, так как следы на поверхности осадка обычно расплывались или смывались течением. Интересно, что многие типы эндогенных следов были открыты в ископаемом состоянии задолго до того, как обнаружили их современные аналоги. Возможности детально изучить ископаемые следы в зафиксированном состоянии, их этологическая и экологическая интерпретация позволили экстраполировать в современность некоторые особенности образа жизни и поведения определенных групп животных. Особенно важно это оказалось для биологов, изучающих ныне живущих морских беспозвоночных, населяющих толщу осадка, наблюдать которых в естественных условиях чрезвычайно сложно. Это еще один пример "обратной" связи науки о прошлом с наукой о настоящем.
Для обозначения ископаемых следов при их описании используют бинарную латинскую номенклатуру. Употребляя термины genus и species, исследователи никак не коррелируют их с определенными родами и видами животных. Больше того, одно и то же животное может оставить столь разнообразные следы, что их будут определять как различные виды, а чаще роды. Например, следы покоя трилобитов относят к роду Rusophycus Hall, следы ползания в толще осадка - к роду Crusiana d'Orb., следы бегания по поверхности - к Dlplichites Daw и т. д. Вместе с тем совершенно различные животные могут образовывать идентичные следы, которые должны относиться к одному ихнороду. Это положение в номенклатуре ископаемых следов представляется наиболее удачным, поскольку в большинстве случаев продьюсер (организм, оставивший след) - неизвестен. Случаи совместного нахождения ископаемых следов и их продьюсеров очень редки и относятся в основном к скелетной фауне [1250, 2056].
Номенклатурные сложности палеоихнологии тесно связаны с проблемой классификации ископаемых следов. Разнообразие следов, которое может оставить отдельное животное, а также зависимость формы следа от случайных свойств осадка исключают возможность решения классификационных и номенклатурных вопросов так, как это делается с нео- и палеозоологическими объектами.
Действительно, за всю историю палеоихнологических исследований предпринято несколько попыток создания различных классификаций ископаемых следов [201, 231, 1608, 1703]. В их основу были положены: 1) форма ископаемых следов (морфологический подход); 2) характер сохранности и положение следов в осадке (топономический, или стратиномический подход); 3) поведенческая интерпретация ископаемых следов (этологический подход); 4) таксономическая интерпретация следов (таксономический подход); 5) комбинация таксономического, морфологического и топономического подходов.
Морфологический подход к классифицированию ископаемых следов имеет глубокие исторические корни. На ранних этапах развития палеонтологии, когда многие следы, особенно U-образные и цилиндрические ветвящиеся норки, описывались как остатки морских водорослей, чисто морфологический подход к изучению был вполне логичным. И хотя в конце прошлого столетия была выяснена и доказана истинная природа этих окаменелостей [1788, 2000, 2001], позже не раз предпринимались попытки объединить ископаемые следы по общности морфологических особенностей в группы, подобные семействам [145, 1610, 1848, 1876, 2156, 2159], хотя последовательно морфологический подход отличает только классификацию Н. Б. Вассоевича. Одной из последних в ряду морфологических является классификация А. Хоровитца [1607], имеющая некоторое сходство с упомянутой ранее классификацией Ж. Лессертиссера.
Как в одной, так и в другой следы подразделяются на две главные группы: 1) интерстратальные структуры и 2) биотурбации, приуроченные к поверхности напластования. Эти группы в свою очередь подразделяются по форме, структуре, характеру наполнения, размерам и ориентировке. Чисто морфологический подход в изучении ископаемых следов, по-видимому, может быть полезным в случаях, когда неизвестна или малоизвестна природа их признаков или когда существует необходимость проследить изменчивость формы следов, в том числе пространственную и временную. Возможно, именно подробные морфологические классификации, созданные для некоторых групп ископаемых следов, окажутся полезными при решении некоторых вопросов стратиграфии, палеоэкологии и палеонтологии. Тот факт, что большинства из уже имеющихся морфологических классификаций не используется палеоихнологами, вероятно, подтверждает целесообразность разработки не всеобъемлющих морфологических классификаций, а детальных классификаций отдельных групп следов.
Большинство ископаемых следов сохраняется на поверхности раздела литологически разных пород, например между глинами и более грубозернистыми осадками. Последние обычно играют роль скульптурного материала, заполняющего следы, запечатленные в более пластичной глинистой матрице. Это хорошо известно по ископаемым следам, которые встречаются во флишевых толщах на подошвах более грубозернистых кластических пород [1850, 1851]. Однако нередки случаи нахождения следов и на верхней поверхности напластования - верхние биоглифы [206].
Характер сохранности и положение следов по отношению к поверхности раздела литологически разных пород послужили основой для создания топономических (стратиномических) классификаций ископаемых следов. Одна из таких классификаций была разработана А. Зейлахером [2261, 2262, 2267]. Способы образования изображенных форм, их изменения и фоссилизация, отражение в специальной (геологической) терминологии, были продемонстрированы рядом исследователей [1420, 2056, 2267, 2456 и др.]. Делались попытки классифицировать следы на основе их хронологических взаимоотношений с вмещающей породой, в частности на предепозиционные и постдепозиционные [1850], хотя некоторые формы невозможно отнести ни к одной из этих групп [1703]. Топономическая классификация А. Мартинсона [1922] основана на взаимоотношении скульптурного материала (обычно более грубозернистых пород) и матрицы (обычно глин). Биотурбации на нижней поверхности слоя скульптурного материала относятся к группе Hypichnia, на верхней поверхности - Epichnia, внутри слоя - Endichnia, вне его - Exichnia. От названий этих четырех групп образуются прилагательные, которые используются с обычными описательными морфологическими и экологическими терминами. Классификация А. Мартинсона ценна тем, что при своей лаконичности отражает: 1) положение следов по отношению к скульптурному материалу, 2) механику образования следов, ликвидируя необходимость в параллельном наборе генетических терминов, 3) характер рельефа. Топономические классификации, несомненно, полезны при изучении следов с морфологической и топографической точек зрения, а также для определения характеристик осадконакопления и фоссилизации следов.
Даже невыразительные в таксономическом отношении ископаемые следы позволяют получить большое количество информации о поведении животных, о некоторых наиболее общих их функциях, таких, как, например, передвижение или питание. Поведенческая интерпретация следов, трактующая биологические функции, открыла еще одну возможность их исследований и классификации.
Известная этологическая классификация А. Зейлахера [2261] основана на том, что различные группы животных с одинаковым образом жизни и близкими особенностями поведения оставляют похожие следы, хотя сами животные имеют совершенно различную форму тела. А. Зейлахер различает пять этологических групп, с характерными морфологическими особенностями.
1. Domichnia (жилища) - простые, разветвленные или U-образные норки, перпендикулярные или наклонные к поверхности напластования, а также системы нор с горизонтальными и вертикальными компонентами. Более или менее постоянные жилища для большинства фильтратов, ведущих полусидячий образ жизни.
2. Fodinichnia (следы питания) - норки разнообразной формы со шпреитами и без них, располагающиеся под различными углами к поверхности напластования. Более или менее временные норки, созданные грунтоедами, которые использовались ими как жилища, ходы для перемещения и т. п.
3. Pascichnia (следы пастьбы) - сильно извивающиеся полосы и борозды, не пересекающие друг друга, отличающиеся интенсивным использованием поверхности осадка, годного для пастьбы и поедания. Обычно эти следы выглядят как орнаментация поверхности (меандры, "елочки", лабиринты и т. п.).
4. Cubichnia (следы покоя) - изолированные, главным образом мелкие депрессии, очертания которых грубо соответствуют форме продьюсеров. Обычно располагаются параллельно друг другу в результате ориентировки животных навстречу течению (реотаксис). Возможна вертикальная и горизонтальная повторяемость.
5. Repichnia (следы ползания) - борозды, трековые дорожки, тоннели различных направлений, искривленные и прямолинейные, иногда ветвящиеся, гладкие и со скульптурой.
Эта классификация была намеренно ограничена небольшим количеством категорий, наиболее полезных в функциональном исследовании биотурбаций и палеоэкологических реконструкций. Включение дополнительных категорий в экологическую классификацию следов [1981] не получило признания, и в настоящее время палеоихнологи используют в основном оригинальный вариант классификации А. Зейлахера (рис. 102). Однако нередко нет возможности отнести тот или иной след к определенной группе, поскольку он отражает сразу две функции. Например, на поверхности осадка, окружающего норку-жилище некоторых полихет, обычно видны радиально-лучистые следы проедания ила, таким образом, след в целом должен описываться, как комбинация Domichnia и Pascichnia [1072]. Известны следы, которые интерпретируются как камеры для выведения потомства каллианассид [1473]. Эти следы не находят места в указанной классификации. Кроме того, сложные следы, которые обычно относят к группе Pascichnia (следы пастьбы), отражают подчас самые разные действия. В одном случае - это поедание ила, в другом - создание ловушек для микроорганизмов, мигрирующих в осадке (некоторые мелкие аннелиды роют в песке на литорали плоскоспиральные норки, которые затем повторно используют, поедая диатомовые водоросли и другие микроорганизмы, попавшие в норку с током воды из осадка - своеобразный способ обогащения субстрата необходимой пищей). В третьем случае, как, например, с известной формой Paleodictyon сложная деятельность животных, ведущая к постройке хорошо аэрируемой системы тоннелей в виде сети с шестигранной ячеей, интерпретируется как создание "фермы" для выращивания грибов и других микроорганизмов "к столу" хозяина норки [2272].
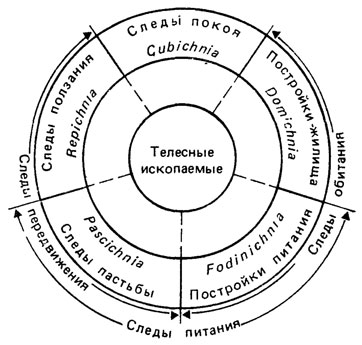
Рис. 102. Экологическая классификация следов жизнедеятельности. По А. Зейлахеру
Несмотря на эти сложности, этологическая классификация позволила сравнивать различные ихноценозы на основе ихнологических спектров, а также учитывать, какими этологическими группами представлен тот или иной ихноценоз и какой процент общего количества его следов относится к каждой из имеющихся этологических групп. Использование методики ихноспектров привело А. Зейлахера к созданию концепции батиметрической зональности ископаемых следов [2272], которая оказалась чрезвычайно плодотворной в палеоэкологии и палеогеографии.
Этологический подход к классификации следов был развит в ряде работ Ф. Фюрзиха [1612, 1613], в которых на примере родов Diplocraterion и Rhizocorallium предложена концепция "существенных" и "акцессорных" морфологических признаков, имеющих различное значение для таксономии ископаемых следов. Существенные признаку являются результатом поведения высокого уровня значимости и поэтому являются родовыми (например, наличие U-образной трубки и шпреитов у Rhizocorallium). Акцессорные признаки возникают в результате поведения низкого уровня значимости (это в основном признаки, отражающие разнообразные реакции организмов на физические и химические явления в окружающей среде) и должны использоваться для классификации следов на видовом уровне. Недостаточно разработана в этой концепции проблема выбора легко различимых и устойчивых акцессорных признаков, которые не должны быть результатом седиментационных и постгенетических явлений. Эта процедура в значительной мере субъективна.
Создание иерархической классификации следов, по-видимому, принципиально невозможно. Необходимо искать другие пути. Один из них - изучение поведенческих программ и их эволюции. Это интересное и перспективное направление современной палеоихнологии касается прежде всего морфологически сложных следов группы Pascichnia [1091, 1470, 2271, 2272]. Как правило, траектория движения животных изучается независимо от их таксономической принадлежности. Функции и побудительные стимулы, вызвавшие ту или иную реакцию поведения, нередко игнорируются (часто они выявляются в результате исследования).
Сложность траекторий ископаемых следов, особенно в фациях, отвечающих биотопам со стабильными условиями среды и ограниченными пищевыми ресурсами, давно привлекает внимание палеоихнологов. В качестве основных стимулов, заставлявших грунтоедов использовать сложные поведенческие программы при поедании ила, выдвигалось стремление наиболее полно и оптимально охватывать осадок и не пересекать свой собственный след. Формализация указанных стимулов для компьютера [2127] позволила получить с его помощью многие из траекторий, которые реально наблюдаются при изучении следов с повторяющимися элементами симметрии (меандры, спирали и пр.). Введение в компьютер иных ограничивающих стимулов привело к созданию целого мира "фантастических следов червей" [1621], которые хотя и далеки от реальных следов, но намечают методически интересный путь развития эволюционной этологической палеоихнологии, едва ли не самой увлекательной области изучения ископаемых следов.
Эволюция поведения морского подвижного бентоса, в частности грунтоедов и детритофагов, вероятно, шла двумя параллельными путями. Первый - возникновение новых простых способов охвата организмами осадка, например, меандрирование, движение по спирали, движение из одной точки в разных радиально расходящихся направлениях и т. п. Эти элементарные пространственные реализации (а мы говорим сейчас только о перемещении организмов в пространстве безотносительно к стимулам и пр.) характеризуются повторением движений одного типа. Второй путь - создание разнообразных комбинаций из двух, очень редко трех простых поведенческих программ, о которых уже сказано (рис. 103). Однако этим не исчерпывается содержание эволюции поведения: большинство простых и сложных поведенческих программ усложняются во времени, они как бы оптимизируются. Интересно сопоставить эволюцию поведения грунтоедов с развитием способов отработки месторождений полезных ископаемых. Оказывается, что и беспозвоночные (даже те, которые жили 600 млн. лет назад!) и человек во многих случаях использовали принципиально сходные приемы охвата пространства.
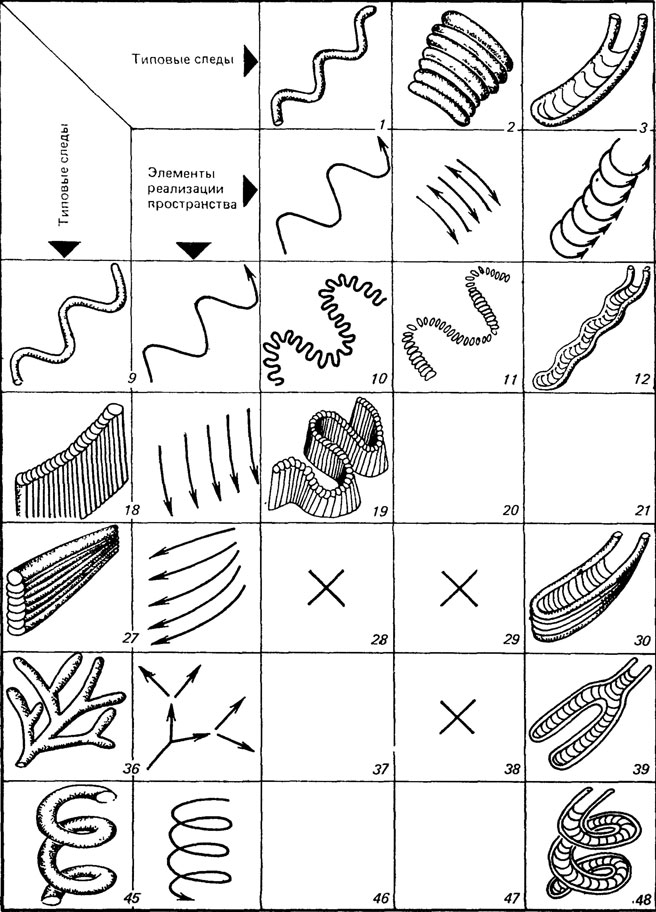
Рис. 103. Возможная классификация ископаемых следов с повторяющимися элементами симметрии. По М. А. Федонкину. Типовые следы и движения, в результате которых эти следы были образованы (ЭПР), обозначены одним номером (верхний ряд - 1-8, левый ряд - 9, 18, 27, 36, 45). На пересечении координат указанных ЭПР изображены следы, образовавшиеся путем комбинации двух ЭПР. Перечеркнутые клетки указывают на малую вероятность или запрещенность комбинации соответствующих ЭПР из верхнего и левого рядов, ничем не заполненные - теоретически возможные комбинации. 1, 9 - Cochlichnus; 2 - Palaeopascichnus delicatus; 3 - Rhizocorallium; 4 - Spirodesmos archimedeus; 5 - Oldhamia antiqua; 6 - Asterichnus tawrencensis; 7 - Helicodromites mobills; 8 - Phycodes pedum; 10 - Cosmorhaphe sinuosa; 11 - Palaeopascichnus sinuosus; 12 - Rhizocorallium irregulare; 13 - Spirophycus bicornis; 16 - Helicolithus sampelayoi; 18 - Syringomorpha nilssoni; 19, 22 - Dictyodora simpler, 24 - Hellochone hunsrueckiana, 27 - Feichichnus rectus; 30 - Vertically retrusivi Rhizocorallium; 31 - Asterosoma (Foom Helicoid Funnel); 32 - Oldhamia antiqua; 33 - Oldhamia radtata; 35 - 'Small' teichichnia; 36 - Chondrites fircatus von Sternberg; 39 - Rhizocorallium irregulare; 41 - Chondrites, 42 - Taenidium; 43 - Helicolilhus, 45 - Gyrolithet Häntzschel; 48 - Rhizocorallium uliarense; 51 - Zoophycos

Рис. 103. Продолжение. Возможная классификация ископаемых следов с повторяющимися элементами симметрии. По М. А. Федонкину. Типовые следы и движения, в результате которых эти следы были образованы (ЭПР), обозначены одним номером (верхний ряд - 1-8, левый ряд - 9, 18, 27, 36, 45). На пересечении координат указанных ЭПР изображены следы, образовавшиеся путем комбинации двух ЭПР. Перечеркнутые клетки указывают на малую вероятность или запрещенность комбинации соответствующих ЭПР из верхнего и левого рядов, ничем не заполненные - теоретически возможные комбинации. 1, 9 - Cochlichnus; 2 - Palaeopascichnus delicatus; 3 - Rhizocorallium; 4 - Spirodesmos archimedeus; 5 - Oldhamia antiqua; 6 - Asterichnus tawrencensis; 7 - Hdicodromites mobills; 8 - Phycodes pedum; 10 - Cosmorhaphe sinuosa; 11 - Palaeopascichnus sinuosus; 12 - Rhizocorallium irregulare; 13 - Spirophycus bicornis; 16 - Helicolithus sampelayoi; 18 - Syringomorpha nilssoni; 19, 22 - Dictyodora simpler, 24 - Hellochone hunsrueckiana, 27 - Feichichnus rectus; 30 - Vertically retrusivi Rhizocorallium; 31 - Asterosoma (Foom Helicoid Funnel); 32 - Oldhamia antiqua; 33 - Oldhamia radtata; 35 - 'Small' teichichnia; 36 - Chondrites fircatus von Sternberg; 39 - Rhizocorallium irregulare; 41 - Chondrites, 42 - Taenidium; 43 - Helicolilhus, 45 - Gyrolithet Hantzschel; 48 - Rhizocorallium uliarense; 51 - Zoophycos
Следы, отражающие определенные способы охвата пространства, в массовом количестве появились в венде (около 700 млн. лет назад), но следы многоклеточных животных известны и в более древних отложениях. Изучение ископаемых следов, особенно интенсивное за последние 15-20 лет, раскрыло не только их удивительное разнообразие, но и вполне определенную связь их морфологии, которая обусловлена, по-видимому, относительно небольшим набором функций и стимулов, общих для большинства бентосных животных, а также схожими реакциями на воздействие одинаковых экологических факторов.
Это явление, а также существование на протяжении десятков, а иногда и сотен миллионов лет определенных форм следов (меандрических, спиральных, винтообразных, радиально-лучистых и др.), оставленных заведомо различными группами животных, может свидетельствовать об унаследовании поведенческих программ. Наиболее ярко это видно на примере следов пастьбы грунтоедов. Уже в позднем докембрии появляются определенные системы поведения при поедании ила, с помощью которых достигается максимальный и оптимальный охват осадка, насыщенного органическим детритом (рис. 104). На протяжении фанерозоя непрерывно усложняется и оптимизируется поведение грунтоедов в процессе питания путем образования новых систем и в результате сочетания различных поведенческих программ [1091].

Рис. 104. Древнейшие следы жизнедеятельности морских бентосных животных и их стратиграфическое распространение. По М. А. Федонкину. 1 - Planolites Nicholson, 2 - Gordia Emmons 3 - Bilinichnus Fedonkin et Palij, 4 - Berga ueria Prantl, 5 - Bergaueria major Palij, 6 - Palaeopasci chnus sinuosus Fedonkin, 7 - P sinuosus Fedonkin, 8 - Coch lichnus Hitchock, 9 - Neonereites uniserialis Seilachev, 10 - N biserialis Seilachev, 11 - N renarius Fedonkin, 12 - Neon oxites curvus Fedonkin, 13 - Vendichnus vendicus Fedonkin, 14 - Suzmites volutatus Fedonkin, 15 - Vimenites bacillaris Fedonkin, 16 - Intrites puncta tus Fedonkin, 17 - Aulichnites Fenton et Fenton, 18 - Skolithos declinatus Fedonkin, 19 - Skolitos linearis Haldemann, 20 - Helminthoida Schafhautl, 21 - Medvezhichnus pudicum Fedonkin, 22 - Yelovichnus gracilis Fedonkin, 23 - Harlaniella podolica Sokolov, 24 - Didyma ulichnus tirasensis Fedonkin, 25 - Didymaulichnus meanderiformis Fedonkin, 26 - Gyrolithes polonicus Fedonkin, 27 - Phycodes pedum Seilacher, 28 - Treptichnus bifurcus Miller, 29 - Treptichnus triplex Palij, 30 - Teichichnus rectus Seilacher, 31 - Olenichnus irregularis Fedonkin, 32 - Planispihchnus grandis Fedonkin, 33 - Protospiralichnus circularis Fedonkin, 34 - Rusophycus Hall, 35 - Rlizocorallium Xenker, 36 - Diplocraterion Torell, 37 - Chondrites von Sternberg
Одна из важных задач палеоихнологии - установление наиболее древних ископаемых следов. Здесь исследователи сталкиваются со многими трудностями и прежде всего с проблемой распознавания биогенных структур. Критерии различия органогенных и неорганогенных окаменелостей разработаны весьма слабо [1647]; в данном случае большую пользу принесло бы коллекционирование заведомо абиогенных, но сходных с биогенными образований. Анализ литературы по древнейшим ископаемым следам [144, 1090] показал, что несомненные следы животных, хотя и немногочисленные, установлены уже в среднерифейских отложениях, возраст которых более 1 млрд. лет [801, 1391, 1431, 1636, 2196, 2262], хотя не исключено наличие и более древних следов [1819]. Типология их разнообразна: чаще всего это слепки удлиненных гладких, реже - поперечно-морщинистых норок и желобков, сходных со следами ползания современных аннелид и двустворок, небольшие плоскоспиральные образования Helminthoidichnites, которые в последнее время интерпретируются как остатки водорослей [249]. Реже встречаются билатеральные следы и округлые радиально-лучистые образования, органическое происхождение которых иногда оспаривается [1436].
Высказываются мнения, что переход Metazoa к ползанию вызвал появление у животных билатеральной симметрии [60, 327 и др.]. Некоторые исследователи связывают норообразование с возникновением целома, выполнявшего прежде всего опорную функцию (гидростатический скелет), сегментации аннелид и тканей на коллагеновой основе [1427, 1637]. Если допустить, что эти высказывания справедливы, то, судя по данным о древнейших следах, можно предполагать существование уже в среднем рифее билатерально-симметричных многоклеточных целоматной степени организации. В этом случае возникновение Metazoa и их эволюция в доцеломатной стадии могли происходить по крайней мере в раннем рифее, т. е. около 1,5 млрд. лет назад. В то же время мы не в праве исключать возможности существования в глубоком докембрии организмов неизвестной нам, но принципиально отличной от Metazoa организации, которые тоже могли оставлять следы в осадке.
Кроме того, следует помнить, что довольно глубокие крупные норы оставляют в мягком осадке не только целомические животные, но и такие примитивные организмы, как кишечнополостные, например бесскелетные кораллы. Количество и разнообразие следов постепенно возрастает на протяжении позднего докембрия, однако они все еще редки [2186, 2290, 2313 и др.]. В верхнерифейских толщах кроме следов получают широкое распространение комкообразные или палочковидные проблематики группы Vermiculites Reitl, которые, возможно, являются копролитами илоядных животных. Их количество резко возрастает к концу докембрия [370].
Венд, по-видимому, был особым этапом в длительном процессе освоения животными дна докембрийского океана. Даже внося поправку на относительно меньшую изученность довендских отложений, можно утверждать, что вендские следы существенно отличаются от следов из более древних толщ. От более древних следов их отличает морфологическая выразительность, сложность строения и разнообразие. Кроме того, в венде значительно возрастает количество следов. Достаточно сказать, что большинство следов, известных в докембрии, относится к венду и его аналогам.
Ископаемые следы, известные из вендских отложений, свидетельствуют о высокой организации вагильного бентоса этой эпохи [803, 1089, 1090, 1576, 1633, 1636]. В толще осадка эти животные передвигались с помощью перистальтических изменений формы всего тела, как современные немертины, аннелиды, сипункулиды и другие, пропуская осадок через пищеварительный тракт. Об этом свидетельствуют следы Planolites и Neonereites (см. рис. 104) В приповерхностном слое и на поверхности осадка вендские Меtazoa передвигались при помощи волн перистальтики, проходящих по центральной части тела, как у современных планарий, хитонов и гастропод (см. рис. 104), реже - при помощи волнообразных изгибаний тела (Cochliichnus). Чрезвычайно редки в венде следы передвижения с участием параподий.
В качестве особой черты вендских ихноценозов следует назвать следы с признаками симметрии. Эти следы были образованы в результате повторения однотипных движений илоядными животными для оптимального охвата осадка при передвижении. Животные, продвигаясь вперед, выедали широкую полосу богатого детритом осадка, чередуя крутые правые и левые повороты (Nenoxites, Palaeopascichnus и т. п.). Подобные следы широко распространены в вендских отложениях различных регионов и свидетельствуют о явном усложнении поведения грунтоедов в процессе питания в вендскую эпоху.
Наибольший интерес представляют те отложения венда, в которых наряду со следами встречаются отпечатки бесскелетной фауны [483, 756, 808, 983, 1092, 1639]. Как правило, эти животные таксономически никак не соотносятся со следами, так как представлены в основном нектонными формами - различными медузами, примитивными червями, редкими аннелидами и членистоногими и многочисленными седентарными (колониальными и одиночными полипами). Вагильный бентос, представленный, вероятно, аннелидами и моллюсками, судя по следам, в общем отличает более высокий уровень организации при значительно меньшем морфологическом разнообразии. Причины этих различий, если верна интерпретация следов, возможно, состоят в специфически направленной и относительно ускоренной эволюции вагильного бентоса по сравнению с пелагическими формами, хотя не исключено, что бентос имеет значительно более долгую историю развития, чем пелагические Metazoa. Этому не противоречат и находки следов в довендских отложениях, в которых пока не обнаружено достоверных отпечатков животных. Вполне вероятно, что мы не умеем диагностировать их или, как предполагает Б. С. Соколов [985, 986], биохимические функции выделения защитных склеропротеинов у древнейших многоклеточных были недостаточно развиты, поэтому из-за слабой уплотненности покровных образований продьюсеры древнейших ископаемых следов не могли сохраниться в виде отпечатков.
Поразительная смена характера ихноценозов наблюдается на границе венда и нижнего кембрия. В общем она характеризуется более высокой степенью биологической переработки осадка, значительным разнообразием следов и широким развитием сложных следов питания, среди которых наряду с субгоризонтальными и неглубокими (как в докембрии) появляются вертикально ориентированные "двухмерные" и "трехмерные" системы, не свойственные докембрийским ихноценозам (см. рис. 104).
Одна из характерных черт большинства форм ископаемых следов - широкий диапазон их стратиграфического распространения [1876, 2267, 2268]. Однако существуют и такие ихнороды, которые являются или могут служить руководящими формами при определении возраста отложений, например следы ползания трилобитов (Crusiana). В толщах неопределенного возраста, где отсутствуют обычные (телесные) окаменелости, нередко встречаются многочисленные и разнообразные следы. Они успешно используются как для определения возраста, так и для корреляции разрезов [1465-1467, 1469, 2264, 2270].
В палеозойских отложениях ископаемые следы наиболее многочисленны в мелководных песчано-глинистых фациях. В мезозойских и кайнозойских толщах наибольшее разнообразие, наоборот, характерно для глубоководных дистальных турбидитов, обычно бедных телесными окаменелостями [1469]. Особенно богаты палеоихнологическими объектами флишевые отложения [201, 202, 1850, 1851 и др.]. Они изобилуют разнообразными, морфологически сложными следами, дающими важный материал для размышлений о колонизации относительно глубоководных зон моря, подверженных периодическим воздействиям турбидитных потоков, о выработке у беспозвоночных организмов стереотипов поведения, устойчивых на протяжении очень длительного геологического времени, тогда как морская и наземная биоты претерпели колоссальные и неоднократные изменения.
Смещение количества и разнообразия следов в сторону более глубоководных фаций, которое наблюдается на протяжении фанерозоя, безусловно, отражает ход колонизации дна океанов бентосными животными [1468]. Этот процесс шел непрерывно, начиная с позднего докембрия, и сопровождался скачками на некоторых его этапах, в частности, на границе докембрия и кембрия, в ордовике и в позднем мезозое. Восстановление истории освоения организмами дна Мирового океана по палеоихнологическим данным является одной из актуальных, но пока мало разработанных палеобиологических проблем.
Нередко слои с определенными следами, например, с длинными вертикальными норками (Skolithos) или U-образными норками (Diplocraterion) служат великолепными маркерами и прослеживаются на десятки километров при небольшой мощности слоя. Это явление связано с тем, что многие следы имеют узкую фациальную приуроченность.
В тектонически осложненных районах ископаемые следы часто служат указателями подошвы и кровли пласта, степени сжатия пород и характера их деформации.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'