
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Основные направления исследований
Палеоэкологические исследования издавна ведутся по двум тесно связанным между собой направлениям - аутоэкологическому (экология таксонов) и синэкологическому (экология сообществ). Их можно рассматривать также как последовательные этапы работы палеоэколога: аутоэкологический анализ способствует выявлению жизненных форм, сведения о которых служат основой для реставрации палеосообществ и их характеристики. Однако, по нашему мнению, целесообразно выделить основные направления палеоэкологических исследований по иному принципу.
Исследование биологических систем. Для удобства изложения остановимся сначала на оценке значимости отдельных особей, затем перейдем к популяциям, биоценозам и, наконец, к биотам.
Особь. Все палеонтологические, в том числе и палеоэкологические, исследования начинаются с изучения отдельного экземпляра (особи), даже если в ориктоценозе (или образце) количество их исчисляется сотнями. Это связано с высокой информативностью каждой особи как представителя конкретного таксона - фенотипическом воплощении генотипа. В результате функционального анализа морфологии экземпляра (особи) у палеоэколога складывается представление об образе жизни конкретного организма. Данные функциональной морфологии до сих пор считаются наиболее надежными в палеоэкологии. Иногда они подкрепляются тафономическими наблюдениями над находками экземпляров в прижизненном положении. Нередко же тафономические свидетельства выступают на первый план: частые находки строго ориентированных в слое остатков какого-либо таксона позволяют судить об этоологии организма. Такие сведения особенно ценны для полностью вымерших и плохо сохраняющихся в ископаемом состоянии групп.
Ярким примером этого служат описанные Р. Ф. Геккером [223, 226, 229] остатки организмов на абрадированном скальном дне и галечниках девонского моря Восточно-Европейской платформы - приросшие раковины брахиопод и двустворок, скелеты кораллов (табулят, ругоз), морских лилий и текоидей, трубки морских червей. По находкам отдельных экземпляров какого-либо таксона в разных фациях устанавливают пределы фактора (или факторов) среды его обитания. Изучение периодичности роста скелетов морских беспозвоночных, позволяет предполагать сезонность изменений климата в далеком геологическом прошлом и даже судить о продолжительности года и суток в палеозое и мезозое. Раковины - главные объекты палеобиогеохимических и палеобиохимических исследований. Таким образом, информативная ценность отдельной особи в палеоэкологии весьма высока. Тем не менее на уровне особи не могут быть выработаны представления об оптимуме условий для процветания вида, взаимоотношениях видов, сравнительной значимости их в палеосообществах.
Все эти палеоэкологические характеристики можно получить только при изучении ископаемых популяций.
Популяция. Ископаемые популяции - это сохранившиеся в слоях части популяций некогда существовавших видов. Принадлежность экземпляров к единой популяции устанавливается с помощью комплекса полевых и лабораторных исследований. Локальная популяция в ископаемом состоянии представлена совокупностью экземпляров последовательных поколений вида в едином автохтонном танатоценозе, сформированном при непрерывном накоплении осадков [394, 395]. Это основано на признании таких особенностей популяции, как территориальные ограничения и относительно стабильные условия обитания в течение времени ее существования, общность исторической судьбы всех входящих в ее состав особей, определенная возрастная структура, морфологическая (следовательно, и генетическая) целостность. Следует подчеркнуть, что нормальное распределение признаков важное, но не решающее условие принадлежности к единой популяции. Понятие ископаемой популяции в принципе более емко, чем современной популяции. В этом нет ничего удивительного. В слое нередко запечатлены все этапы исторического развития популяции - становление, расцвет и угасание (рис. 94). Непосредственное наблюдение этапов развития современных природных популяций, учитывая значительную продолжительность их существования, нереально.
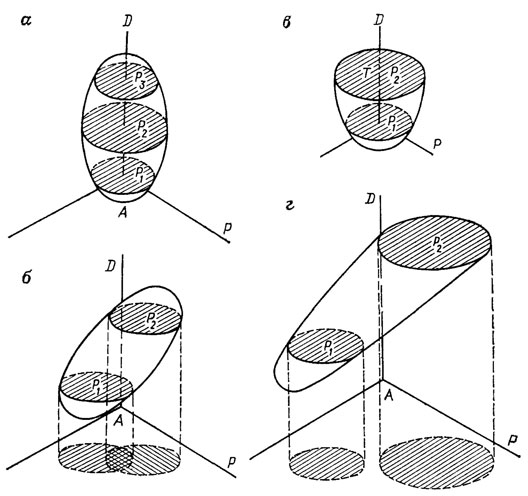
Рис. 94. Пространственно-временные модели ископаемых популяций, показывающие возможные варианты их судьбы. По В. А. Захарову. Зарождение и вымирание без смены (а) и со сменой местообитания (б); катастрофическую гибель (время Т) в пределах исходного местообитания (в) и со смещением ареала (г); ось А-Д - время; Р1, Р2, Р3 - площади расселения популяции в разные моменты времени
Чтобы эффективно оперировать популяциями в палеоэкологии, следует выявить их характеристики, конечно, лишь те из них, которые устанавливаются на ископаемом материале. С помощью методов, рассмотренных раньше, можно оценить популяционную плотность или показатели относительной численности, например частоту встречаемости в баллах. Количественные характеристики позволяют судить об экологическом состоянии вида, о роли его в разных сообществах, о взаимоотношении с другими экологически сродственными видами и прочем. Из динамических характеристик возможно определение динамики смертности: размерно-частотные кривые в какой-то мере будут отражать возрастную структуру популяций, позволят судить об условиях обитания на разных этапах ее существования. В ископаемых популяциях фиксируются также периодические колебания численности - циклические осцилляции [767]. Тафономические наблюдения над ископаемыми популяциями помогают понять структуру исходных популяций. Например, установление особенностей размещения экземпляров в пределах слоя, заключающего исследуемую ископаемую популяцию, способствует выяснению закономерностей распределения особей или их групп в исходной популяции.
Популяция должна быть основной (элементарной) операционной единицей при любых палеобиологических исследованиях: таксономических, эволюционных и экологических. Именно такую роль играют популяции при анализе палеобиоценозов.
Палеобиоценоз. Палеобиоценозом было предложено называть сохранившуюся в ископаемом состоянии часть биоценоза [226]. В современной литературе чаще используется термин "сообщество". Однако этот термин довольно часто употребляется для обозначения любых группировок таксонов, встреченных в захоронении как совместно, так и в пределах определенного биостратиграфического интервала. Нередко одни и те же группировки видов при стратиграфическом анализе называют комплексами, а при палеоэкологическом - сообществами. Такая аналогия не всегда правомерна. Палеоэколог только тогда получает право оперировать группировкой встреченных в едином ориктоценозе видов как с частью биотического сообщества, когда докажет, что в исходном сообществе эти виды сосуществовали. Условия совместного существования видов с определенными экологическими особенностями входит во все определения биотического сообщества, независимо от разных взглядов на природу сообществ.
В морской гидробиологии сообщества интерпретируются в свете концепции биоценоза, по К. А. Мёбиусу, как группировки популяций сосуществующих и функционально связанных между собой видов. В палеоэкологии донных беспозвоночных сообщества обычно рассматриваются как группировки популяций независимых видов, связанных лишь сходными реакциями на абиотические факторы среды [1308, 1802]. Если учесть, что многие явления природы не перестают существовать оттого, что до конца непоняты и не становятся более понятными от замены термина, то следовало бы вернуться к изначальному термину биоценоз*.
* (В настоящей работе термины палеобиоценоз, ископаемое сообщество и палеосообщество употребляются как синонимы.)
Следует еще раз напомнить, что в современных морских мелководных сообществах лишь около 30% видов имеют твердый скелет и способны сохраняться в ископаемом состоянии [1802]. Следовательно, палеосообщество даже по основным характеристикам иногда будет отличаться от исходного, за счет которого оно образовалось. Однако у палеоэколога есть одно преимущество перед экологом: он может наблюдать геологическую историю палеосообществ. Ископаемое сообщество обычно ограничено слоем (или частью слоя), заключающим совокупность остатков организмов (ассоциацию палеопопуляций разных таксонов), захороненных на месте жизни. Прижизненные группировки могут быть также выявлены в результате установленной наблюдениями на разрезе повторной встречаемости ассоциаций (реккурентности видов).
Сохранившиеся в слоях остатки биоценозов должны отражать результаты прижизненных взаимоотношений между вымершими организмами. Надо полагать, что эти отношения в прошлом контролировались теми же законами, что и сейчас. Следовательно, при описании палеобиоценоза правомочна ассимиляция концепции современного биотического сообщества и аппарата понятий и терминов, с помощью которых оно описывается [767]. Несмотря на утрату многих характеристик, палеобиоценоз сохраняет часть сведений об исходном биотическом сообществе - его таксономической структуре (доминировании, видовом разнообразии, выровненности), трофической структуре (способе и уровнях питания) и функциональных особенностях (свидетельствах конкуренции, комменсализма, паразитизма, хищничества, мутуализма), результатах r-отбора и K-отбора, специализации по нишам, жизненных циклах, т. е. устойчивости во времени. Палеобиоценозы именуются, так же как и современные донные морские биоценозы, по одному или нескольким доминирующим видам или же по типу биотопа. Иногда употребляются обе характеристики [678]. Палеобиоценозы можно также подразделять на ассоциации видов - группы таксонов, входящих наряду с другими в состав сообщества. Так, например, выделяется ассоциация организмов - обрастателей на крупных раковинах эпифаунного вида.
Элементарной функциональной единицей сообщества является жизненная форма. Она объединяет группы видов и родов со сходным образом жизни - способом питания, этологией, отношением к абиотическим факторам среды. Жизненная форма служит единицей палеоэкологических классификаций [233, 679]. В палеоэкологии морских беспозвоночных наиболее распространены группировки по трофическому признаку (рис. 95): способу питания и уровням питания [402, 905, 2448]. Выделяются также экологические кислородные, гидродинамические, батиметрические, термальные, солевые, эдафические и морфологические группировки. В сочетании с количественными показателями указанные классификации используются для характеристики палеосообществ, анализа взаимоотношений образующих их популяций, динамики во времени и пространстве. Палеоэкологические группировки служат основным источником информации при биофациальном анализе.
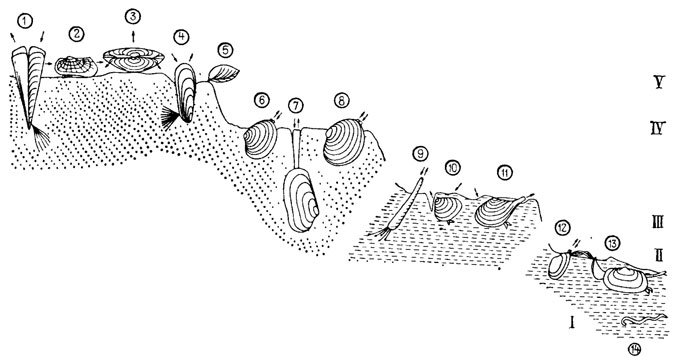
Рис. 95. Принципиальная схема размещения бентоса в юрском бореальном море по трофическим группам. По А. В. Захарову, Б. Н. Шурыгину. Цифры в кружках: 1-8 - фильтраторы; 9-14 - детритофаги по уровням питания (I-V). 1 - Pinna; 2 - Grammatodon; 3 - Boreionectes; 4 - 'Musculus'; 5 - Terebratulidae; 6 - Astarte; 7 - Pleuromya; 8 - Arctica; 9 - Scaphopoda; 10 - Nuculoma; 11 - Dacryomya; 12 - Nuculana; 13 - Matletia; 14 - Vermes
Изучение взаимоотношений организмов в палеобиоценозах. На палеонтологическом материале документально устанавливаются следующие типы взаимоотношений между организмами: конкуренция, симбиоз (комменсализм и мутуализм), паразитизм и хищничество. Некоторые необычные тафономические ситуации интерпретируются с позиций нейтрализма (терпимости) и антибиоза (аменсализма), однако приводятся лишь косвенные (актуалистические) соображения, прямых доказательств нет [1259].
Конкуренция. Кажется несколько неожиданным тот факт, что исследованию конкуренции - одному из фундаментальных явлений в органическом мире, в значительной мере обеспечивающему эволюционный процесс, уделяется очень мало внимания в палеоэкологии. Тезис о "конкурентной борьбе" еще со времени создания теории естественного отбора обычно лишь постулируется в палеонтологической и палеоэкологической литературе. Возможно, что это связано с бытующим и в настоящее время в палеоэкологии организмоцентрическим подходом.
В природе конкуренция имеет место внутри популяций и между популяциями. Экспериментально она может быть доказана только на популяционном уровне [617]. Таким образом, свидетельства конкуренции в органическом мире геологического прошлого следует искать, изучая не отдельные экземпляры, а ископаемые популяции видов.
Известно, что наиболее жесткая конкуренция существует между особями внутри популяций, а также между популяциями видов со сходными экологическими требованиями. Результаты конкуренции, закрепленной естественным отбором, можно изучать на единой ископаемой популяции путем сравнения последовательных кривых распределения мерических признаков, построенных для разных временных уровней. Серия таких кривых хорошо фиксирует типы внутрипопуляционного отбора - стабилизирующего, направленного или дизруптивного [685, рис. 27].
Под конкурирующими всегда понимают виды, одновременно нуждающиеся в одних и тех же жизненно важных ресурсах [594]. Острота межвидовой конкуренции возрастает по мере увеличения популяционной плотности сосуществующих видов. Вследствие этого два вида с одинаковыми экологическими требованиями не могут продолжительно сосуществовать (принципы конкурентного исключения, или правило Гаузе). Анализ 29 мелководных донных биоценозов Японского моря, изученных гидробиологами с помощью легководолазной техники [1002], показал что, двустворчатые моллюски и гастроподы в своем распределении по биотопам подчиняются указанному правилу. Среди 23 биоценозов, в которых присутствуют двустворки, нет ни одного случая совместного поселения хотя бы двух близкородственных видов (100% исключения!). Из 28 биоценозов, заселенных гастроподами, в 18 случаях (64,3%) каждый род представлен только одним видом, в четырех биоценозах (14,3%) один род - тремя видами, только в двух - четырьмя видами [394, рис. 2].
Палеонтологический материал также дает многочисленные свидетельства подобных отношений экологически родственных видов. Правда, палеонтологу обычно нелегко судить о том, что лежало в основе межвидовой конкуренции, поскольку в чистом виде вычленить регулирующий фактор сложно. Приведем примеры, касающиеся мезокайнозойских моллюсков. Так, Т. Зоргенфрай показал, что четыре вида Nassa более или менее взаимно исключались в среднем миоцене на юге Ютландии (рис. 96). Виды из рода Buchia в позднеюрских и раннемеловых бассейнах на севере СССР сосуществовали на единых биотопах лишь при низкой популяционной плотности одного или чаще обоих видов. В моменты резкого доминирования одного из видов другие были подавлены или полностью вытеснялись с биотопов (рис. 97). Экологическая близость не всегда предполагает таксономическую близость. Поэтому правило экологического исключения касается не только близкородственных видов, но и других "экологически сродственных" таксонов. К таким группам в раннемеловом море на севере Сибири принадлежали бухии и иноцерамы. Эти данные моллюски относились к одному экологическому типу биссусных и принадлежали к единой трофической группировке - фильтраторам высокого уровня. Благоприятны для их процветания были илисто-глинистые грунты в условиях средней части сублиторали. В этих условиях бухии и иноцерамы конкурировали между собой: в поселениях с высокой популяционной плотностью одного рода другой был подавлен (рис. 98).
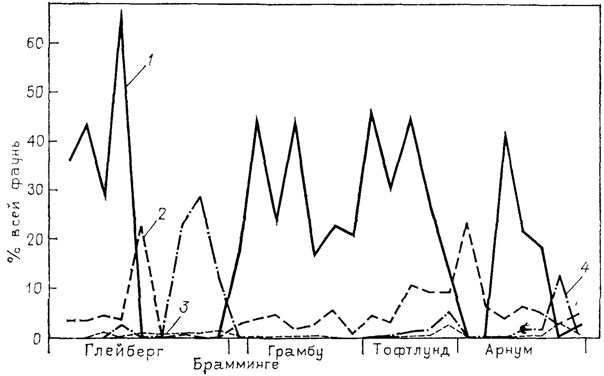
Рис. 96. Кривые, демонстрирующие взаимное исключение видов Nassa (гастроподы) в среднем миоцене на юге п-ова Ютландия. По Д. В. Эгеру. 1 - Nassa cimbrica; 2 - N. schlotheimi; 3 - N. facki; 4 - N. fuchsi
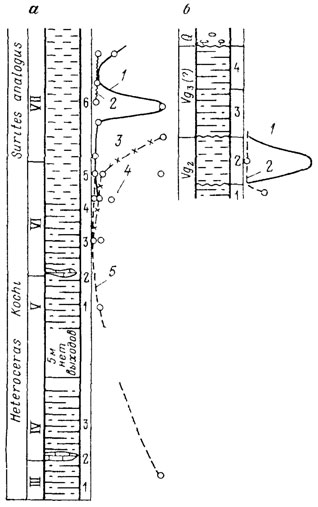
Рис. 97. Биотопическое взаимоисключение близкородственных видов Buchia демонстрируют кривые частоты встречаемости. По В. А. Захарову. а - Buchia unschensis (1), B. tolmatschowi (2), B. fasikivi (3), B. okensis (4), в берриасе, север Сибири, р. Боярка. б - B. russiensis (1), В. mosquensis (2) в пограничных слоях между средне- и верхневолжским подъярусами, Северное Притиманье, р. Волонга. Римские цифры - номера слоев в опорном разрезе неокома; арабские цифры - номера слоев в обнажениях. Масштаб 1:400
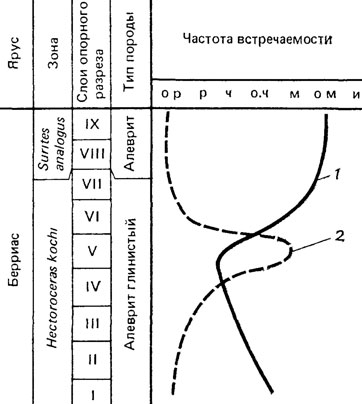
Рис. 98. Кривые, отражающие биотопическое исключение Buchia. По В. А. Захарову, А. С. Турбиной. 1 - бухии; 2 - иноцерамы; берриас на р. Боярке, Хатангская впадина севера Сибири (о. р - очень редко; р - редко; ч - часто; о. ч - очень часто; м - много; о. м - очень много; и - изобилие)
Симбиоз. Кооперация как один из путей адаптации в борьбе за существование была широко распространена в органическом мире геологического прошлого. Палеоэкологическая литература изобилует примерами симбиотических взаимоотношений между морскими беспозвоночными в течение фанерозоя. Этот тип биотических отношений документально подтверждается находками окаменелостей, запечатлевших совместное проживание организмов в течение всей жизни или на отдельных ее этапах.
В симбиоз вступали группы, далекие в филогенетическом и таксономическом отношениях. Даже в современном мире нелегко точно определить конкретный характер связи между организмами. Замечено, однако, что мутуалистические отношения обычно устанавливаются между автотрофами и гетеротрофами. Этот тип симбиоза обнаружен у мшанки Fenestella и бурой водоросли, а также у каменноугольной мшанки Thamniscus pinnatus и красной водоросли [1551]. Предполагается, что мельчайшие (обычно 2-15 мкм) шаровидные или овальные образования, обнаруженные у мезозойских и третичных жгутиковых, ордовикских и силурийских фораминифер, являются остатками симбионтов типа современных зооксантелл или зоохлорелл [1547].
Очень полно в палеоэкологической литературе освещен другой тип симбиоза - комменсализм. Заключение об этом типе связи основывается обычно на закономерном расположении скелета гостя - комменсала на раковине хозяина. Р. Ф. Геккер [232] выделил три ситуации, фиксирующие комменсальные биотические взаимоотношения.
1. Нарастание (цементация) одних форм на другие. Например, трубочки аулопор (кораллы, табуляты), серпул и спирорбиса покрывают раковины девонских брахиопод [223, 226, 232, 622, 1239, 1258 и др.]; комменсалами позднемеловых брахиопод были фораминиферы, черви-трубкожилы, одиночные склерактинии и альционарии, мшанки и двустворки [480]; колонии мшанки Berenicea инкрустировали раковины среднеюрских гастропод при их жизни, а затем вступали в симбиотические отношения с раками-отшельниками, селившимися в пустых раковинах [2065]; описан симбиоз морских уточек (усоногие раки) и аммонитов в раннемеловом море на Северном Кавказе [346], устриц и аммонитов [398, 1946] и другие [см. 1752].
2. Внедрение одних форм в твердые образования других. Среди инквилинистов-комменсалов отмечаются фораминиферы, например, в оксфордских кремниевых губках [1820], а также многощетинковые черви, например Palaeosabella в раковине позднедевонской брахиоподы Cyrtospirifer (232, рис. 6] и Polydora в раковинах палеогеновых устриц [235] и раннемеловых пектинид [393] и склерактинии [549].
3. Обрастание одних форм другими. Обрастание или иммурация гостя-комменсала при тесном совместном проживании с организмом-хозяином широко распространено у колониальных животных. Примером могут служить черви-комменсалы в колониях ордовикских мшанок [221, 232], силурийских табулят [980] и раннемеловых склерактинии [549]; В. Н. Дубатолов [351] обнаружил симбиоз позднедевонских строматопороидей и сирингопорид.
В случае менее прочных связей между симбионтами (прикрепление биссусом двустворок, ножкой брахиопод, ногой гастропод и др.) свидетельства взаимосвязей, как правило, утрачивались. Хотя несколько примеров симбионтов-гастропод описаны, например, поселение платицератид вблизи анального отверстия карбоновой морской лилии [1239] или же поселения калипторид и брахиопод на раковине крупных раннемеловых пектинид [392]. Таким образом, возможно выделение четвертой ситуации - разновидности эпибиоза: мягкого прикрепления гостя-комменсала на раковине хозяина.
Анализ прижизненных поселений организмов-комменсалов на раковине животного-хозяина нередко помогает в реконструкции образа жизни последнего. Так, например, особенности размещения домиков Spirorbis подтверждают мнение о том, что прижизненным для силурийских брахиопод Camarotoechia и Homoeospira было положение брюшной створкой вверх [1768]. По особенностям ориентировки и строения сверлений усоногих на рострах юрских и нижнемеловых белемнитов сделан вывод о передвижении этих головоногих моллюсков преимущественно головой вперед [2269].
Паразитизм. К документальным свидетельствам паразитизма среди организмов геологического прошлого обычно относятся такие случаи внедрения одних организмов в другие, которые сопровождаются аномальным развитием раковины хозяина.
Следствием паразитизма кольчатого червя из подкласса Myzostomidae в стебле мезозойских и палеозойских морских лилий явилось утолщение стебля, изменения формы и расположения его члеников [226, 232, 1239]. А. Н. Соловьев [1000] рассматривал рачка Canceripustula nocens, внедрявшегося в панцири юрского морского ежа Collyrites elliptica, как эндопаразита. Предполагается паразитизм червя Mycelites в раковинах юрских аммонитов из ФРГ и Центральной части Европейской территории СССР. Обнаруженные плотные скопления нитей на протравленных поверхностях перегородок и раковин были интерпретированы как гифы паразитических грибов [2225, 2485]. Однако не всегда возможно провести четкую грань между действием инквилиниста-комменсала и инквилиниста-паразита. Так, если моллюски-камнеточцы Lithophaga, сверлящие губки из рода Cliona и сверлящие многощетинковые черви Polydora, в массовом количестве селились на раковины палеогеновых устриц при их жизни, то это приводило к разрушению раковин и гибели моллюсков-хозяев [232].
Хищничество. Последствия взаимоотношений хищник - жертва фиксируются на палеонтологическом материале в виде залеченных травм на раковине и скелете, следов сверлений (химической или механической природы), содержания желудков хищников, фоссилизированных находок хищников на своих жертвах. В литературе описаны многочисленные случаи регенерации раковин морских беспозвоночных после укусов хищников: брахиопод [913], белемнитов [226, 904], аммонитов [226, 341, 467, 1295], двустворок [1817]. Обычными палеонтологическими находками считаются следы сверлений хищных брюхоногих (натицид) на раковинах двустворчатых и брюхоногих моллюсков мезозоя и кайнозоя; такие же следы были обнаружены на раковинах палеозойских брахиопод [232, 1237, 1239, 1404].
Содержание желудков юрских плезиозавров и ихтиозавров показывает, что эти крупные хищники питались белемнитами и рыбами, в желудочной их массе обнаружено много крючков Onychites и рыбья чешуя [1259, 2096]. Остатки девонских морских звезд обнаружены на раковинах двустворок; омар из золенгофенского верхнеюрского сланца встречен на фоссилизированной рыбе. Все эти случаи трактуются как гибель и захоронение хищников в момент захвата жертв [1259].
Исследование палеобиоценозов в пространстве и времени. При наличии детальной стратиграфической основы палеоэколог имеет возможность изучить палеобиоценозы как на площади палеобассейна (в пространстве), так и на разных этапах его развития (во времени).
Исследуя топологию палеобиоценозов в пределах палеобассейна определенного интервала времени, палеоэколог стремится вскрыть закономерности их размещения на площади. Обычно закономерная смена типов палеосообществ опосредована "эффектом берега" и соответствует нормальному спектру фаций. В морской палеоэкологии ряды палеобиоценозов, прослеживающиеся от палеоберега в сторону моря, были названы "гаммами" [226]. Закономерная смена ассоциаций наземных растений на площади получила название катена [520]. Нормальную последовательность палеобиоценозов в принципе можно восстановить не только в результате сравнительного изучения синхронных отложений по латерали, но и при изучении последовательности наслоения в едином разрезе, имея в виду, что контактирующие в нормальном залегании слои формировались в палеобассейне в пограничных фациальных обстановках (закон Головкинского - Вальтера). Однако последовательные ряды, полученные в результате распределения асинхронных палеобиоценозов, не будут адекватны исходным из-за эволюционных новаций в палеосообществах. Эволюционный эффект тесно связан с продолжительностью времени формирования толщи, оценить которое всегда нелегко. Таким образом, пространственная и временная последовательности сообществ отличаются качественно.
Исследованию латеральных рядов палеобиоценозов в бассейнах геологического прошлого уделялось исключительное внимание в работах Р. Ф. Геккера и его группы [224, 226, 235]. В последнее время, особенно в зарубежной литературе, снова возрос интерес к палеобиоценозам. Значительное внимание, как и раньше, уделяется экологии сообществ, но главным образом изучается структура палеобиоценозов [1271, 1372, 1614, 1877, 2515, 2516 и др.]. Основные положения, обсуждаемые в этих работах, сводятся к следующему. Распределение бентосных сообществ в прошлом, как и теперь, контролировалось факторами среды. Суммарный эффект факторов обусловливал субпараллельное размещение их палеоберегу. В том случае, если удаление от палеоберега сопровождалось постепенным углублением дна, границы сообществ были также субпараллельными и по мере углубления возрастало разнообразие состава сообществ при доминировании в каждом из них одной или немногих групп бентоса. Число сообществ, образовывавших нормальный ряд (гамму), лимитировалось тектонической обстановкой, типом береговой линии и уклоном дна. В областях со спокойным тектоническим режимом, устойчивой береговой линией и пологим дном гаммы сообществ были более многочисленными (рис. 99), чем на участках с изменчивой тектонической обстановкой, изрезанной береговой линией и крутыми уклонами дна (рис. 100). Большинство исследователей склоняется к выводу о прямом контроле сообществ гидродинамикой; как правило, тип сообщества был связан с определенным типом грунта, который в свою очередь опосредован гидродинамикой (в условиях нормально падающего профиля равновесия дна). Все случаи существенного расхождения с такой интерпретацией должны быть тщательно проверены [2515].
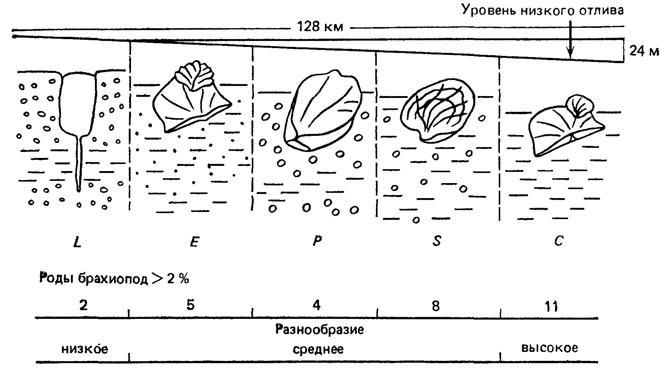
Рис. 99. Нормальный ряд (гамма) из пяти бентосных сообществ пологого дна раннесилурийского моря в области спокойного тектонического режима и устойчивой береговой линии. По Э. Ж. Андерсону. На абсолютной (верхняя линия) и относительной (нижняя линия) шкалах показано разнообразие наиболее распространенных родов брахиопод; изображены преобладающие роды: Lingula (L), Eocoella (E), Pentamerus (P), Stricklandia (S), Clorinda (С). Уклон дна показан приблизительно; для раннего силура он составляет 30-120 см на 1,6 км, для позднего ордовика - 120-360 см на 1,6 км (приведены только минимальные цифры)
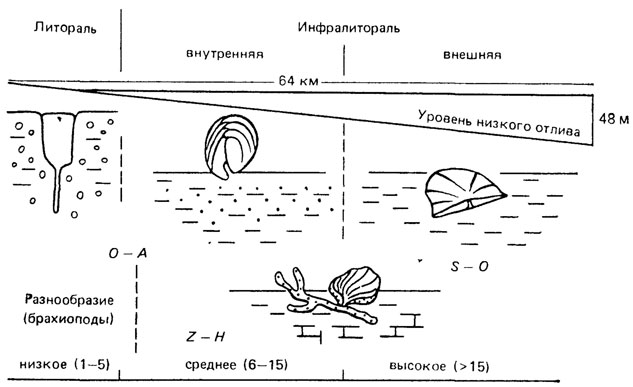
Рис. 100. Нормальный ряд (гамма) из трех или двух бентосных сообществ позднеордовикского моря с относительно крутым уклоном дна, изменчивой тектонической обстановкой и изрезанной береговой линией. По Э. Ж. Андерсону. Разнообразие наиболее распространенных родов брахиопод показано на относительной шкале. Изображены формы трех сообществ: Orthorhynchia - Amborhynchia (О-А), Sowerbyella - Ohniella (S-O) Zygospira - Hebertella (Z-H). Сообщество (О-А) разделяется на два, сообщество (Z-H) замещает два других в инфралиторали при слабом поступлении пластического материала
Один из выводов, полученных на основании анализа главным образом палеозойских донных сообществ, состоит в том, что таксономическое разнообразие растет с удалением от палеоберега и углублением субстрата. Этот вывод не вызывает сомнения для батиметрических моделей в условиях верхней сублиторали. Однако более широкое обобщение - разнообразие растет с глубиной, так как стабильность среды увеличивается [1518] - вряд ли справедливо. В современной гидробиологии преобладает мнение, что общее разнообразие в сообществах снижается как в полностью стабильной среде, так и часто изменяющейся. Максимально видовое разнообразие присуще средам с оптимальной частотой колебаний физических факторов. К такому заключению склоняют также данные экспериментальной экологии [1655, 2058]. Хотя использование более совершенных орудий лова существенно меняет бытовавшие представления о значительной бедности фауны, в частности мейофауны на больших глубинах в океане.
Последовательность автохтонных элементов ориктоценозов в разрезе позволяет судить о смене сообществ во времени. В палеоэкологии наземных растений такую последовательность иногда называют палеосукцессией [520]. В морской палеоэкологии различают два разных по природе типа последовательностей - один связан с реккуренцией сообществ, другой с собственно сукцессией. Реккурентное сообщество почти не меняется во времени ни по таксономическому составу, ни по структуре. Повторное появление остатков такого сообщества в разрезе обычно объясняется возвратом условий обитания, чрезвычайно сходных по параметрам с предшествующими. Примером может служить бентосное сообщество, повторявшееся 49 раз в свите Макохета (ордовик, шт. Минессота в США). Остатки сообщества, главным образом из брахиопод и двустворок, заключены в глинистые органогенные известняки и полностью отсутствуют в чередующихся с ними доломитах [1313]. Для контактирующих реккурентных сообществ характерны резкие границы и отсутствие переходной зоны хроноэкотона.
Сообщества, развивающиеся сукцессионно, как правило, проходят последовательные стадии от пионерных до климаксных. Обычно они образуют композиционно-структурный континуум во времени, поэтому границы между контактирующими сообществами нечеткие. Первые стадии сукцессии проходят под преимущественным контролем абиотических факторов, среди которых преобладает r-отбор, позднее состав и структура сообществ формируется при существенном влиянии биотических составляющих среды и возрастает роль K-отбора. Процесс сукцессии сообществ характеризуется следующими чертами, зафиксированными в серии соответствующих ориктоценозов: постепенным возрастанием видового разнообразия и дифференциацией по уровням питания; усилением доминирования на главных трофических уровнях немногих видов; уменьшением выровненности; появлением групп раковинных видов и усилением их роли; сокращением экологических ниш ряда видов; усилением роли комменсалов, хищников и другими признаками всеобщего гомеостаза. Предполагается, что экологические сукцессии должны смягчать стрессовые воздействия среды обитания и увеличивать в целом стабильность экосистем [767]. Сукцессии морских бентосных донных сообществ наиболее полно представлены в трансгрессирующих сериях (рис. 101). В литературе описаны экологические сукцессии морского бентоса, сопровождавшиеся постепенными преобразованиями биотопов: медленной литификацией морского дна [1648] или заменой мягкого илистого биотопа более жестким песчаным вследствие усиления донных течений [1373]. При сукцессии рифовых сообществ активную роль в преобразовании биотопов играют сами организмы.
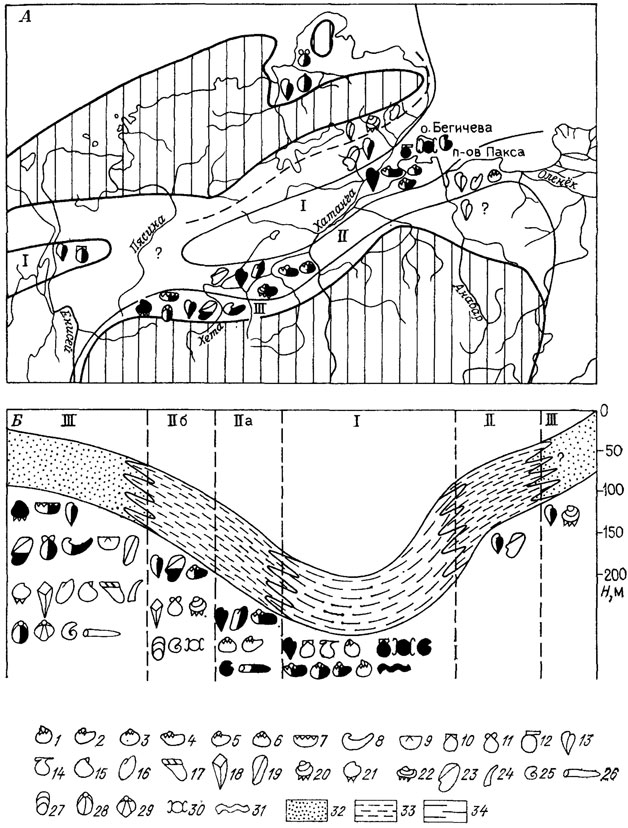
Рис. 101. Схема фаций, батиметрический профиль и сообщества беспозвоночных в раннеберриасском море на севере Средней Сибири. По В. А. Захарову. А - сообщества беспозвоночных, значительно дифференцированные по биономическим зонам трансгрессирующего моря. Б - схема фаций и батиметрический профиль раннеберриасского моря в направлении: р. лета - р. Боярка - п-ов Пакса - Восточный Таймыр Заштрихована суша; сплошные и пунктирные линии - границы фациальных и биономических зон: I - относительно глубоководная, II - умеренно глубоководная (а - относительно удаленная от беррга, б - приближенная к берегу), III - мелководная. 1 - Nuculoma-Palaeonucula; 2 - Dacryomya; 3 - Malletia; 4 - Taimyrodon; 5 - Jupiteria; 6 - Sarepta; 7 - Grammatodon; 8 - Praexogyra; 9 - Boreionectes; 10 - Capmtonectes; 11 - Entollium; 12 - Aequipecten; 13 - Buchia; 14 - Oxytoma; 15 - Plagiostoma; 16 - Inoceramus; 17 - Aguilerella; 18 - Pinna; 19 - Musculus; 20 - Astarte; 21 - Neocrassina; 22 - Prorokia; 23 - Homomya; 24 - скафоподы (денталиум); 25 - аммониты; 26 - белемниты; 27-29 - брахиоподы (27 - лингулиды, 28 - теребратулиды, 29 - ринхонеллиды); 30 - рыбы; 31 - следы илоедов; 32 - песок; 33 - ил; 34 - глина. Контурами показаны таксоны, встречающиеся сравнительно редко; наполовину зачерненные - встречающиеся часто; полностью зачерненные - встречающиеся в изобилии
Так, анализ экологической сукцессии в восьми древних рифах (из ордовика и силура США, из силура - девона Пакистана, из верхнего девона Бельгии и из верхнего мела Антильских островов) показал присутствие четырех вертикальных зон, соответствующих следующим стадиям формирования рифовых сообществ: 1) подготовки субстрата (скопление остатков иглокожих); 2) колонизации (поселение рифостроящих организмов); 3) диверсификации (рост разнообразия рифостроителей; возрастание сложности пищевых цепей и дифференциация по экологическим нишам); 4) доминирования, характеризующейся резким усилением роста немногих таксонов [2447].
Палеоэкология тесно связана со многими биологическими, геологическими и некоторыми другими естественными науками. Этот аспект палеоэкологии лишь в общих чертах затронут в данной главе (более подробно см. 8.1). Рассмотрен он также в недавно опубликованном учебно-методическом пособии [397].
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'