
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
3.4. Тафономия бесскелетных многоклеточных животных (М. А. Федонкин)
Ни в одной из многочисленных областей палеонтологических исследований вопросы тафономии не имеют столь большого значения, как при изучении фации, включающей организмы без минерального скелета. Здесь палеонтологи сталкиваются с тафономическими задачами на всех стадиях работы - от поисков отложений, благоприятных для сохранности остатков бесскелетных, до морфологической и экологической интерпретации окаменел остей. Вместе с тем условия захоронения бесскелетной фауны и особенно сложные и разнообразные процессы образования окаменелостей и местонахождений остатков бесскелетных беспозвоночных наименее изученная область тафономии. Это объясняется прежде всего дефицитом материала - местонахождения бесскелетных крайне редки на фоне изобилующих скелетными остатками, а иногда и нацело сложенных скелетным материалом пород фанерозоя.
Уникальным по распространению мягкотелых организмов можно считать вендский период. При значительном разнообразии животного мира (в венде сейчас известны все основные типы беспозвоночных) организмы этого периода были бесскелетными. В настоящее время известно несколько богатейших местонахождений бесскелетной фауны, подобных беломорскому и эдиакарскому [483, 756, 1092, 1094, 1639].
Остановимся подробнее на анализе наиболее важных факторов среды, которые способствовали массовому захоронению и фоссилизации бесскелетных метазоа. При этом попытаемся отметить и чисто вендские явления, не характерные для последующей геологической истории. Обобщение же данных по тафономии мягкотелых беспозвоночных фанерозоя имеет значение для выявления широкого спектра условий, в которых фоссилизировались остатки этих организмов, а это, в свою очередь, может послужить основой для направленного поиска новых местонахождений докембрийской фауны.
Анализ местонахождений бесскелетных многоклеточных свидетельствует, что в большинстве случаев мы имеем дело с танатоценозами. Однако то, что последние представлены остатками лишенных скелета беспозвоночных, имеет два важных следствия. Во-первых, местонахождения бесскелетных представляют собой ассоциации одновременно живших организмов, большая часть которых обитала на небольшом расстоянии от мест захоронения. Мягкие ткани легко подвергаются бактериальному разложению и обычно не отличаются устойчивостью к деструктивным механическим воздействиям. Это исключает возможность длительного переноса тел погибших организмов, а переотложение фоссилизированных остатков делает просто невероятным, поскольку окаменелости бесскелетных сложены ни чем иным, как веществом вмещающей породы и формировались in situ. Это важно не только для палеоэкологических реконструкций, но и для биостратиграфических построений.
Процессы захоронения остатков бесскелетных организмов, как и характер сохранности образовавшихся ихнофоссилий, контролировались разнообразными факторами. Различные сочетания этих факторов так или иначе нарушали представления о первичном (прижизненном) соотношении численности представителей разных видов и других таксонов и экологических групп.
Среди факторов, определяемых особенностями каждого организма в отдельности, можно назвать; 1) размеры тела, 2) плотность тканей тела, 3) устойчивость их к механическим, химическим и биохимическим агентам деструкции, 4) наличие или отсутствие химических или механических приспособлений, отпугивающих хищников и трупоедов, 5) место обитания, 6) положение в пищевой цепи. На эти факторы накладывалось действие других, внешних по отношению к каждому животному факторов: 1) характера движения воды, 2) свойства субстрата, на котором обитало животное, 3) свойства осадка, в котором оно было погребено, 4) скорости осадконакопления и 5) интенсивности биологической переработки осадка. Мы еще проиллюстрируем влияние каждого из перечисленных факторов в отдельности либо в совокупности с другими.
В беломорской биоте венда (впрочем, это характерно и для эдиакарской биоты) наблюдается интересная закономерность - по мере усложнения организации животных уменьшаются их численность, видовое разнообразие и размеры тела [1091, 1096]. При этом чем меньше размеры данного бесскелетного организма, тем соответственно меньше вероятность сохранения его остатков в ископаемом состоянии. В связи с тем, что кишечнополостные были наиболее крупными животными в рассматриваемой биоте, они чаще сохраняются и соответственно чаще попадаются палеонтологу. Отсюда представления о большей их численности и видовом разнообразии.
Размеры тела животных и соответственно образовавшейся окаменелости имеют несомненное значение в практической работе палеонтологов. Отпечатки размеров менее 2-3 мм, которые нередко встречаются вместе с более крупными на плоскостях напластования мелкозернистых пород, практически очень трудно интерпретировать, поскольку тонкие детали их строения сравнимы с размером зерен породы. Как правило, в столь малых по размерам отпечатках не удается установить иных деталей строения, кроме очертаний тела. Сложно бывает обнаружить и целые отпечатки крупных животных, достигающих в длину метра и более, особенно если они имели гладкую поверхность.
Плотность тела животного могла влиять на характер прижизненного или посмертного переноса, хотя в последнем случае газообразные продукты бактериального разложения уменьшали плотность и увеличивали плавучесть тела. У морских бесскелетных животных плотность тканей определяется содержанием воды, количеством органического вещества и его структурированностью. Известно, что чем выше организация животных, тем ниже содержание воды в тканях, но даже и у таких примитивных форм, как кишечнополостные, разные ткани (клеточный слой, мезоглея, гонады) имеют разную плотность и поэтому разную вероятность сохраниться в ископаемом состоянии. Например, у мелких вендских медуз, обладавших тонким нежным зонтиком, лучше всего сохраняются гонады и каналы гастроваскулярной системы, т. е. органы, в состав которых входит сравнительно меньшее количество воды. Лучшая сохранность гонад и гастроваскулярных каналов может объясняться и тем, что в этих органах в результате деятельности бактерий после гибели организма накапливались газообразные продукты разложения, и за счет внутреннего давления некоторое время поддерживалась упругость этих органов. Если медуза была погребена живой, то мерцательный эпителий каналов не сразу прекращал функционировать, что, по сообщению Н. Н. Марфенина (МГУ), способствовало заполнению гастроваскулярной системы тонкими глинистыми частицами и органическим детритом. У крупных медузовидных полипов с мощной мезоглеей часто сохраняется лишь внешний контур тела. С плотностью тканей коррелирует их устойчивость к механическим воздействиям, позволяющая сохранять форму тела при переносе, а в процессе захоронения существенно деформировать вмещающий осадок и не утрачивать в течение некоторого времени свой объем.
Плотность и дифференциация тканей, по-видимому, должны коррелировать с повышением их устойчивости к химическим и биохимическим агентам деструкции, а также к воздействию бактерий. Поэтому логично предположить, что наиболее многочисленные окаменелости должны представлять наиболее высокоорганизованные группы животных. Но в венде все было иначе - большую часть окаменелостей всегда составляют Radialia. Здесь, по-видимому, сказывалось действие четвертого, пятого и шестого указанных выше факторов. Во-первых, кишечнополостные обладают стрекательными клетками, которые, как было сказано ранее, сохраняли активность даже некоторое время спустя после гибели животного. Эта особенность резко выделяла кишечнополостных среди других животных, поскольку предотвращала возможность быстрого поедания их трупоедами и оставляла время для захоронения в осадке. Свойство стрекательных клеток сохранять свою функцию некоторое время после гибели организма защищало также погребенное тело от падалеедов, которые могли довольствоваться другими организмами, изымая их из числа потенциальных окаменелостей. Во-вторых, среди вендских кишечнополостных известно большое количество разнообразных полипов, которые при жизни деформировали осадок, так как по крайней мере базальная часть была в него погружена. Поэтому они, как правило, не испытывали значительного переноса и захоронялись на месте, не претерпев заметных посмертных изменений. К сказанному надо добавить, что кишечнополостные в венде, вероятно, были столь же активными хищниками, какими они являются и сейчас, т. е. они замыкали пищевые цепи. Все это могло способствовать преимущественному захоронению и фоссилизации кишечнополостных.
Большую роль в формировании ископаемых ассоциаций бесскелетных метазоа играет подвижность воды, которая в значительной степени определяет скорость осадконакопления, состав и структуру осадка, в котором происходит захоронение животного. Скорость течений влияет на перенос живых и погибших организмов и может выступать в качестве фактора, искажающего общую картину сепарации отдельных форм в соответствии с особенностями их размеров, плотности, плавучести, механической устойчивости и т. д.
Наиболее благоприятны для захоронения остатков мягкотелых метазоа толщи, сложенные тонкопереслаивающимися алевролитами и аргиллитами или песчаниками и аргиллитами. При этом отпечатки и следы чаще располагаются на подошве более грубозернистых слоев и реже на их кровле. Это объясняется, по-видимому, тем, что псаммитовый и алевритовый материал накапливается быстрее, чем пелитовый, в связи с тем животные оказывались погребенными в осадке до того, как их тела успевали разложиться. К тому же относительно крупнозернистый осадок значительно облегчает выход продуктам распада тел, в том числе и газам, а это, естественно, способствовало лучшей сохранности остатков организмов в еще нелитифицированной породе. Подстилающая же глинистая порода благодаря большей пластичности запечатлевает тончайшие детали строения тела - многократно ветвящиеся каналы гастроваскулярной системы медуз и т. п.
Окаменелости могли формироваться в двух случаях. Во-первых, когда новая порция более крупнозернистого материала засыпала тело живого или погибшего животного, лежавшее на глинистом субстрате. Во-вторых, когда животное какое-то время пребывало на глинистом грунте, а затем оставленный им отпечаток или след перемещения заполнялся более грубозернистым осадком, игравшим роль скульптурного материала.
Довольно часты находки остатков бесскелетных в глинистых толщах, вмещающих линзы алевролитов. Следы жизнедеятельности и остатки мягкотелых метазоа, как и в предыдущем случае, прослеживаются на подошве алевролитов в виде форм позитивного и негативного гипорельефа. Приуроченность этих окаменелостей к линзам алевролитов объясняется прежде всего различием физико-механических свойств алевролитов и глин, что значительно повышает вероятность фоссилизации и, главное, последующего отчленения этих литологических разностей друг от друга в результате естественного или искусственного препарирования. Однако не исключено, что в ряде случаев накопление алевролитов в виде небольших линз могло быть обусловлено самим наличием на дне живого или погибшего организма: турбулентные вихри, образовывавшиеся вокруг тела в движущемся потоке воды, вымывали тонкие пелитовые частицы вокруг животного, а появившиеся затем углубления могли служить ловушкой для более крупных алевритовых частиц.
Рассматриваемый тип сохранности характерен для подавляющего большинства местонахождений вендских бесскелетных, хотя в общем спектр пород, в которых встречаются окаменелости, значительно шире. Большое количество отпечатков было обнаружено, в частности, в гомогенных неслоистых алевропелитовых породах. В качестве примеров сошлемся на многочисленные Pteridinium из сюзьминских слоев на Летнем берегу Белого моря [756, 1096]. Обширную коллекцию вендской фауны удалось собрать и из тонкослоистых доломитов хатыспытской свиты в долине р. Хорбусуонки на Оленекском поднятии [2303].
Нередко быстрому захоронению и фоссилизации остатков бесскелетных способствовали различные катастрофические явления. Ярким примером может служить фауна Мистекен Пойнт из серии Консепшен (п-ов Авалон, Ньюфаундленд), где на кровле пластов вендских кремнистых алевролитов сохранились многочисленные отпечатки древнейших бесскелетных кишечнополостных великолепной сохранности, Эти животные были практически мгновенно погребены под тонким слоем вулканического пепла, сохранившегося в виде прослоев туффитов. Подобных уровней в разрезе серии Консепнеш немало, что свидетельствует о частых извержениях прибрежных вулканов [1272].
Другое довольно распространенное явление катастрофического характера, способствующее захоронению бесскелетных, - турбидитные потоки и штормы. Последние в характерных для венда условиях мелководных эпиконтинентальных морей играли важную роль. В накопившихся при штормах породах, характеризующихся косой и подвернутой слоистостью, плохой сортированностью, следами эрозии и вмывания осадка в нижележащие слои, остатки бесскелетных животных встречаются довольно редко, особенно в грубозернистых разностях. В более тонких алевролитовых породах отпечатки можно обнаружить значительно чаще, причем их ориентировка может быть различной, во многих случаях не совпадающей с поверхностью напластования и даже не параллельной ей. Обычно в этих случаях отмечаются изогнутые "неправильные" отпечатки, свидетельствующие о деформации тела в процессе захоронения.
Сходная картина наблюдается и в случае захоронения бесскелетных турбидитными потоками, но последние более характерны для областей интенсивного прогибания земной коры, или для склоновых фаций палеоконтинентов. Именно турбидитной природой вмещающих пород объясняется изобилие и разнообразие скелетной и бесскелетной фауны в Берджесс Шейл Британской Колумбии [1452, 2489]. Турбидитный генезис этих сланцев подчеркивается не только структурным положением толщ и их текстурными особенностями, но и разнообразной ориентировкой окаменелостей, а также смешением представителей самых разных местообитаний в одном захоронении.
Наиболее бедные ассоциации окаменелостей характерны для существенно глинистых толщ. Очень малое количество окаменелостей встречается, как правило, в пелитовых темноцветных разностях пород, накопившихся в относительно глубоководных условиях эпиплатформенных морей, вероятно ниже уровня воздействия штормовых волн. В венде в таких толщах изредка попадаются однообразные следы Palaeopascichnus и мелкие седентарные формы медузовидного облика. Большое количество дисперсного органического вещества (скорее всего, растительного происхождения), которое влияет на окраску пород, свидетельствует, что оно лишь в незначительной степени утилизировалось бентосом и пелагическими организмами. Редкость находок остатков вендских метазоа в глинах отчасти связана и с тем, что гомогенные и даже слоистые разности этой породы довольно редко образуют при выветривании хорошо отпрепарированные поверхности напластования. Но главные причины редких находок вендских бесскелетных в тонких пелитовых разностях заключаются, по-видимому, в следующем. Глины являются очень пластичным материалом и в тех случаях, когда в иле погребалось бесскелетное животное (особенно с низким содержанием "сухого" органического вещества в тканях), после разложения которого почти ничего не оставалось, осадок заполнял объем, занимаемый трупом, поверхности подстилающего и перекрывающего его слоев смыкались и пелитовые частицы могли, вероятно, пересекать границу потенциального отпечатка в вертикальных направлениях, тем более, что глина сохраняла пластичность в течение длительного времени. Другой причиной может быть генезис этих отложений, которые во многих случаях были связаны с довольно глубоководными условиями морского бассейна, т. е. с зонами наименьшей биомассы метазоа в докембрии.
Иная картина предстает при исследованиях существенно глинистых, но относительно мелководных толщ с признаками подводного размыва и течений. В самих глинах, которые растрескиваются и рассыпаются в мелкую крошку или размокают под действием поверхностных и грунтовых вод, очень трудно обнаружить остатки бесскелетных или следы жизнедеятельности. Однако растворение глин в дистиллированной воде иногда позволяет выделять оттуда тонкие палочковидные образования, сложенные алевритовыми частицами. Эти образования скорее всего являются фрагментами тонких ходов илоедов. В сухих плотных глинах мелководного происхождения довольно часто встречаются тонкие пиритизированные ходы. Пирит, вероятно, развивался по органическому веществу, которым было обогащено заполнение норок (слизь, фекальный материал).
Существенно иной тип захоронения остатков бесскелетных метазоа приурочен к конкрециям. В определенных условиях, которые еще мало изучены, вокруг погребенного в толще осадка организма образуются конкреции, которые останавливают аэробное разложение тела и предотвращают полное сжатие породы вокруг него. Мягкие части животных в конкрециях сохраняются в виде пятен, цвет которых отличается от цвета окружающей породы, а также в виде тонких минеральных пленок и слепков; иногда сохраняется и слегка измененная кутикула.
Случаи нахождения остатков организмов в разнообразных конкрециях заслуживают особого внимания. Так, например, крупнейшее местонахождение рассматриваемого типа известно с конца прошлого века в пенсильванских отложениях Иллинойса (США). Эта богатейшая ассоциация окаменелостей, известная под названием "фауны Мэйсон Крик", включает представителей кишечнополостных, моллюсков, аннелид, приапулид, немертин и разнообразных проблематичных животных неясного систематического положения, причем в ней присутствуют как бесскелетные, так и скелетообразующие формы [1937]. Эти фоссилии, среди которых доминируют остатки бесскелетных кишечнополостных, сохраняются в сфероидальных и эллипсоидальных конкрециях сидерита размером от 1 до 30 см, залегающих в серых алевропелитовых сланцах. Сохранность окаменелостей в конкрециях такова, что мягкие ткани и даже внутренние органы большинства животных можно наблюдать в ископаемом состоянии. Вмещающие конкреции породы накопились в условиях крупной дельты при стабильном положении уровня моря [2281]. Формированию фоссилии способствовали здесь два процесса: 1) быстрое захоронение организмов, которое предотвращало аэробное разложение, и 2) сингенетическое образование конкреций, которые окружали и "консервировали" организм.
Основная часть хорошо сохранившихся образцов в описываемом местонахождении была найдена в конкрециях, расколовшихся естественным путем благодаря выветриванию и трещинам, которые проходят по самому отпечатку, отделяя его от вмещающей породы. Естественное выветривание вообще является одним из лучших способов препарирования окаменелостей. Хорошие результаты по вскрытию конкреций были получены также путем повторного замораживания и последующего оттаивания их в воде. Повторение этих операций (от двух до пятидесяти раз и более) дает значительно лучший эффект, чем применение механического способа.
Иную методику выделения окаменелостей применил К. Мюллер [1987, 1988]. Растворение в уксусной кислоте черных желваковых известняков из верхнего кембрия Южной Швеции позволило ему извлечь большое количество окаменелостей, в основном остатков членистоногих с хорошо сохранившимися мягкими частями, даже их личинок. Размеры последних не превышают 1 мм, тем не менее с помощью сканирующего микроскопа удалось рассмотреть не только их форму, но и тонкие детали строения придатков. Подобная сохранность и сама возможность химического препарирования организмов с сохранившимися мягкими частями стала возможной благодаря фосфатизации органического вещества в известковой матрице. Не исключено, что исследование конкреций и желваков, подобных описанным выше, позволит обнаружить новые местонахождения древнейших метазоа и создаст дополнительные возможности для изучения морфологии вендских бесскелетных, их стратиграфического и географического распространения.
Образование пирита в условиях разложения органического вещества заслуживает особого внимания. Вполне возможно, что это явление можно будет использовать для детального изучения строения остатков бесскелетных организмов, поскольку в участках наибольшей массы исходного органического вещества следует ожидать и наибольшее содержание пирита в породе.
Находки остатков бесскелетных метазоа в карбонатных толщах пока редки. Это объясняется отчасти тем, что процессы диа- и эпигенеза в карбонатных породах во многих случаях могут приводить к разрушению первичных структур, образованных живыми и погибшими организмами в карбонатном иле. Наиболее мощными деструктивными агентами здесь выступают процессы растворения и перекристаллизации. Тем не менее в тонкослоистых листоватых разностях черных битуминозных доломитов хатыспытской свиты на Оленекском поднятии нами было обнаружено большое количество отпечатков бесскелетных метазоа. Встречаются окаменелости и в более светлых плитчатых разностях вместе с ископаемыми следами жизнедеятельности вендских грунтоедов. Разрез хатыспытской свиты в целом характеризуется наличием большого количества туфогенного материала, который также мог играть существенную роль в захоронении остатков животных.
Мягкие ткани погребенного организма обычно довольно быстро разлагаются. Существуют веские доказательства, что большая часть бесскелетных испытывает определенное коллапсирование в результате разложения тканей до того, как начинается уплотнение и сжатие вмещающего осадка [1455, 1803]. Нередко вокруг остатков бесскелетных наблюдается темное пятно, которое интерпретируется как результат вытекания содержимого из разлагающегося тела в осадок [1451, 1453, 1454]. В большинстве случаев коллапсирование тела в результате разложения происходит по вертикали без заметных размеров по латерали, хотя иногда встречаются отпечатки с двойным контуром, отражающим некоторые боковые деформации [2192, 2443]. По мере разложения мягких тканей объем, занимаемый телом погребенного животного, постепенно заполняется осадком. Нередко на кровле слоя можно видеть пологие депрессии, располагающиеся прямо над слепком организма. Они образуются в результате проседания осадка, перекрывавшего погребенный организм. Реже встречаются случаи подтекания осадка из подстилающего слоя на место разлагающегося тела. Иногда это связано с тем, что продукты разложения удаляются вверх сквозь перекрывающий осадок через вертикальный канал и увлекают за собой пластичный материал подстилающего слоя.
Немалые трудности в интерпретации окаменелостей могут вызывать типичные и случайные деформации мягкого тела при захоронении, такие, например, как нерегулярные концентрические морщинки аборальной части зонтика медуз, которые довольно часто рассматриваются как очертания гастральной полости, кольцевой канал и тому подобное, или редкие радиальные бороздки, трактуемые как радиальные каналы гастроваскулярной системы; бороздки могут оказаться следами разрыва мезоглеи.
Кроме особенностей, связанных с захоронением организма и с процессом разложения тканей, в окаменелостях бесскелетных метазоа обычно запечатлены результаты уплотнения осадка и его последующей истории. Эти литологические процессы выражаются в многократном сжатии (нередко более чем в 10 раз) остатков животного по вертикали, в наличии складок, морщин, прожилков и трещин, изменяющих облик окаменелости. Нередко в тонкослоистых породах на подошве нижнего микрослоя наблюдается слепок лишь наиболее выступающей части организма, а на подошве более высоких микрослоев видны отпечатки других частей тела. Во многих же случаях в аналогичных породах на подошве более грубозернистых прослоев видны в основном слепки его базальных частей. Еще одно важное обстоятельство - в зависимости от характера породы, от плотности тканей и органов животного и от их устойчивости к факторам разложения в породе могут запечатлеться те или иные черты организма. Например, в одних случаях отпечатываются все особенности строения покровов организма, в других - только пищеварительной системы и т. д. Изучение полихет из среднекембрийских Берджесс Шейл Британской Колумбии показало, что совершенно различные части одного и того же организма могут сохраняться в тонкослоистой породе в виде отпечатков. Положение поверхности преимущественного расщепления сланцев в этом случае определяется "конкуренцией" между различными частями тела и контролируется тремя независимыми факторами: площадью поверхности структуры (ткани, органа и др.), толщиной структуры и углом наклона ее поверхности к плоскости напластования [1455]. Эти факторы определяют характер сохранности таким образом, что один экземпляр подчас может демонстрировать разные фрагменты структур в зависимости от того, какой микрослой породы обнажается на конкретном участке окаменелости. В зависимости от толщины, наклона или локальных изменений состава микрослоя даже подобные структуры могут иметь разную сохранность и размеры. Поэтому в большинстве случаев требуется детальное изучение микрорельефа отпечатков; особое внимание должно быть уделено микрослоистости и наклону запечатленных структур к плоскости напластования.
В отличие от фоссилизированных скелетов остатки бесскелетного организма обычно невозможно извлечь из породы, и в каждом конкретном случае можно видеть лишь одну сторону его отпечатка или слепка. Следует заметить, что для многих животных существуют характерные позиции при захоронении, в частности, некоторые современные виды медуз после гибели падают на дно преимущественно книзу аборальной стороной зонтика. Если бы эти медузы могли сохраниться в ископаемом состоянии, то это были бы почти сплошь отпечатки или слепки аборальной стороны зонтика. И в тех редких случаях, когда в осадке запечатлелась бы субумбрелла, она могла быть описана как другой таксон. Корреляция отпечатков и слепков обратных сторон тела одного и того же вида возможна не всегда.
За редчайшим исключением, ископаемые остатки бесскелетных метазоа представляют собой слепки или отпечатки. Случаи находок следов временного пребывания живых организмов на субстрате можно интерпретировать как следы жизнедеятельности, однако последние не всегда позволяют наблюдать все детали морфологии животного, тем более, особенности его внутреннего строения. Еще меньше информации в этом отношении дают следы временного пребывания на поверхности осадка трупов животных. После их удаления с поверхности дна (течением или в результате всплывания по мере накопления газообразных продуктов разложения тканей) оставленный отпечаток, как правило, оплывает, тонкие детали его рельефа исчезают, не говоря уже о том, что даже в случае его хорошей сохранности и фоссилизации отпечаток этот даст представление лишь о форме тела и рельефе одной из его сторон, обращенной вниз. В настоящей главе затронуты лишь некоторые аспекты тафономии бесскелетных организмов, но уже сейчас видно, что данные тафономического анализа имеют большое значение для понимания особенностей экологии, морфологии и уровня организации вымерших организмов [986].
Уникальность сохранности бесскелетной фауны венда определяется спецификой некоторых абиотических факторов и неповторимыми особенностями биоса (рис. 113). Среди абиотических факторов следует отметить существование обширных мелководных материковых морей, благоприятных для обитания растений и животных; протяженные плоские литорали, которые служили громадными естественными ловушками для большой массы организмов, выброшенных приливами и штормами. Вероятно, в таких условиях сформировались знаменитые Эдиакарское местонахождение в Южной Австралии [1089] и местонахождение в ломозовских слоях могилевской свиты Подолии [1096]. Интенсивный снос продуктов денудации с суши обеспечивал высокую скорость осадконакопления, способствовавшую быстрому захоронению животных, которое временами могло иметь катастрофический характер (штормы, турбидитные потоки и т. п.). Мелководность морей, существование обширных литоралей и снос громадного объема продуктов размыва в море - все это вместе обусловило поддержание повышенной мутности воды. Следствием этого было наличие узкой фотической зоны, ниже которой в относительно глубоких зонах могла существовать лишь очень бедная жизнь из-за резкого дефицита кислорода.
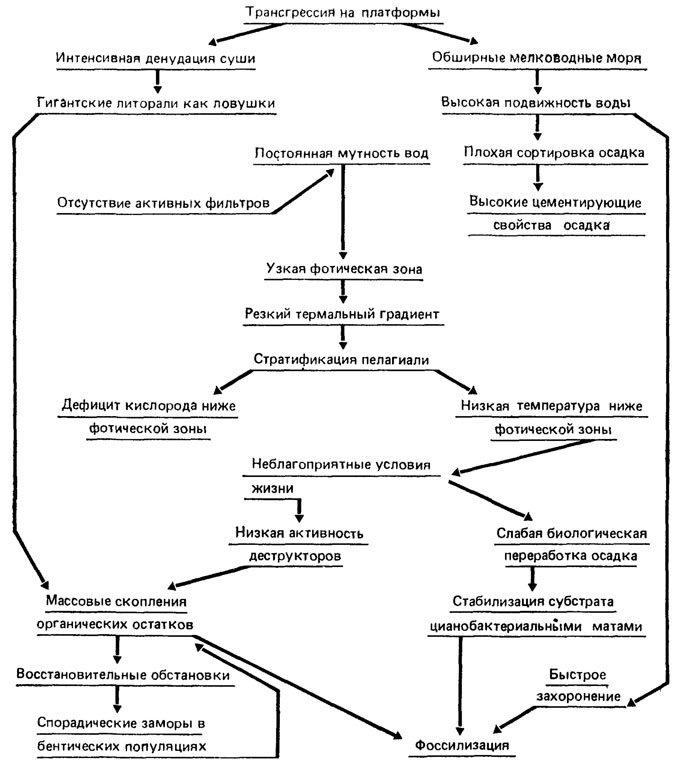
Рис. 113. Некоторые биотические и абиотические факторы среды, способствующие массовому захоронению и сохранности остатков вендских многоклеточных животных. По М. А. Федонкину, оригинал
Резкий температурный градиент, большая разница в температуре относительно тонкого верхнего слоя, хорошо прогреваемого и обогащенного кислородом в результате жизнедеятельности водорослей и растворения атмосферного кислорода, и холодного нижнего слоя морских вод препятствовали перемешиванию и аэрации природных слоев. Все это могло повлечь обеднение жизни в пелагиали и на дне относительно глубоководных зон и служить препятствием к их активной колонизации. Более того, пониженная деструктивная деятельность биоса в пелагиали и на дне из-за низкого содержания кислорода могла приводить к накоплению местами (даже на мелководье в понижениях рельефа дна) больших количеств остатков растений и животных и образованию восстановительной среды в результате медленного разложения органического вещества. Эти явления могли вызывать массовые заморы бентосных популяций, расположенных неподалеку, особенно в тех случаях, когда смена течений перемещала воды, отравленные продуктами разложения, в населенные зоны дна.
Среди важнейших биотических факторов, способствовавших массовому захоронению и сохранности вендских метазоа, можно выделить следующие:
1) высокую плотность популяций морских беспозвоночных в зонах с относительно большой скоростью осадконакопления, значительное количество прикрепленных организмов крупных размеров, в прижизненном состоянии создававших в поверхностном осадке такие структуры, которые потенциально могли запечатлеться в ископаемой летописи;
2) низкую активность плотоядных животных, прежде всего деструкторов-макрофагов, обусловливавшая длительную сохранность погибших животных, достаточную для погребения их под слоем осадка;
3) сравнительно невысокую степень биологической переработки осадка илоедами и другими активными представителями бентоса, способствовавшую ненарушенному состоянию структур, образованных живыми или погибшими животными в мягком осадке.
Следует учитывать также, что многие группы вендских многоклеточных уже обрели к этому времени разнообразные структуры, несшие опорную функцию, а у некоторых известных видов были довольно плотные кутикулярные покровы. Все это позволило им длительное время после захоронения сохранять устойчивость.
На границе докембрия и кембрия ситуация резко изменилась, главным образом благодаря резкой смене палеоэкологической и палеогеографической обстановки. В частности, сократилась площадь литорали, а также резко возросла интенсивность переработки осадков илоедами и другими представителями инфауны. И все же в фанерозойских отложениях встречается много остатков бесскелетных, особенно разнообразных червей. Известны и богатые их местонахождения, такие, как фауна среднекембрийских Берджесс Шейл Британской Колумбии [1454, 2488 и др.] и эссекская фауна из среднего Пенсильвания северного Иллинойса в США [1803, 2380].
Дальнейшее накопление фактического материала приведет, несомненно, к углублению имеющихся и, возможно, установлению новых закономерностей захоронения остатков этих организмов, а следовательно, к дальнейшему прогрессу рассматриваемого направления палеонтологических исследований.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'