
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Часть четвертая. Систематика и филогения
4.1. Принципы и методы палеонтологической систематики (С. В. Мейен)
Принципам и методам систематики, т. е. таксономии современных и ископаемых организмов, посвящена огромная литература. Она включает как сводные работы, касающиеся организмов вообще, так и более частные исследования, затрагивающие методы обработки и интерпретации материала (нумерическую систематику, кладистику и т. п.) конкретных групп организмов (например, аммоноидей) или определенную группу признаков (например, биохимическую систематику). Так как мнения исследователей по обсуждаемым вопросам сильно различаются, целесообразно ограничить круг разбираемых вопросов, сконцентрировав внимание на своеобразии систематики в палеонтологии.
Обычно отличия в таксономии современных и ископаемых форм видят в неполной сохранности последних (отсутствие многих частей, нарушение прижизненной их связи, посмертная деформация частей). Указывается на неприменимость в палеонтологии таких "биологических" признаков, как репродуктивная несовместимость, особенности этологии и прочих. Тем самым получается, что палеонтологическая систематика отличается от неонтологической только обедненным "признаковым пространством" (хотя иногда палеонтологу достается материал исключительно хорошей сохранности), а не какими-то иными принципами. Однако есть основания считать, что неполнота палеонтологических документов настолько велика, что приходится говорить о некоторых принципах и методах, с которыми неонтолог сталкивается лишь в исключительных случаях, т. е. опять же при неполноценном материале. Конечно, следовало бы говорить о различии таксономии не в палеонтологии и неонтологии, а в условиях полноценного и дефектного материала.
Главное отличие обоих видов материала не в том, сколько и каких признаков вовлекаются в анализ, а то, накладывает или нет данный материал окончательный запрет на таксономическое использование и принципиальную наблюдаемость некоторых классов признаков. В случае полноценного материала исследователь волен выбирать любые признаки, хотя практически может пользоваться только немногими из них. В случае же дефектного материала он заранее знает, что некоторые признаки принципиально недоступны, даже если их высокий вес вне сомнений (например, на палеонтологическом и плохо фиксированном неонтологическом музейном материале нельзя наблюдать признаки кариотипа). Поскольку с дефектным материалом палеонтолог сталкивается несравненно чаще, чем неонтолог, в дальнейшем мы будем условно рассматривать работу с дефектным материалом как специфику и прерогативу палеонтологии.
Остановимся на некоторых понятиях и терминах, которые потребуются для дальнейших рассуждений. Изучение разнообразия окружающего, в том числе органического мира идет двумя путями. С одной стороны, мы выделяем группы объектов, с другой - исследуем их свойства. Имеется в виду, что и группы, и свойства объективно существуют. В отношении групп надо различать понятия, соответствующие процессу их получения (1), достигнутому результату в виде списка, схемы или представленному иначе (2), самой объективной группировке (3), если таковую принимать существующей вне зависимости от субъекта и теории всей работы (4). Эти четыре понятия получили разное наименование в литературе. Мы примем для первого понятия термин систематика, для второго и третьего - система или классификация, для четвертого - таксономия. Будем считать, что система - частный случай классификации. Это - "привилегированная" классификация, отобранная исследователем в качестве опорной, базисной.
Остановимся на мерономизации систематики в палеонтологии самостоятельных и несамостоятельных признаках, типологических экстраполяциях, принципе монотопных выборок, проблеме возрастной множественности систем и проблеме свертки таксономического разнообразия. Вопросы систематики, связанные с кладистическим анализом (кладизмом, кладистикой), не рассматриваются (см. 4.3 и 4.4).
Мерономизация систематики в палеонтологии. Многочисленные авторы подчеркивали, что палеонтологическая систематика должна быть систематикой организмов, а не просто дисперсных частей. Особенно много об этом говорилось в связи с растительными остатками, в частности спорами и пыльцой [2238]. Выдвигалось мнение, что надо выделять не роды и виды оболочек спор и пыльцы, а роды и виды растений, представленных лишь этими микрофоссилиями. Некоторые остатки, например конодонты, не удавалось систематизировать при таком подходе, поскольку об общем облике конодонтоносителей известно крайне мало (см. 1.4 и 1.8). Соответственно появилась тенденция различать в палеонтологии таксоны, более или менее эквивалентные таксонам неонтологии (будем называть их эвтаксонами), и паратаксоны, соответствие которых неонтологическим таксонам необязательно.
Легко заметить, что выделение паратаксонов вообще не вполне сопоставимо с выделением эвтаксонов. Паратаксоны объединяют отдельные части организмов. В неонтологии для мысленной группировки частей не используется термин "таксон". Вместо этого говорят об органе определенного типа; соответственно употребляются понятия "правая створка", "опистоцельный позвонок", "перистый лист" и им подобные. Данные понятия несут некоторую таксономическую нагрузку (ясно, что опистоцельные позвонки могут иметь только определенные позвоночные), но в основе своей они морфологические, а не таксономические. Иногда палеонтолог может найти на дисперсных частях достаточно богатый комплекс признаков, который позволяет приблизить классификацию частей к системе соответствующих эвтаксонов. Если такой возможности нет, то классифицируют только дисперсные части и получают именно разные типы частей, а не таксоны организмов. Выделение же типов частей - прерогатива не систематики и не таксономии, а морфологии. По сути, здесь следует говорить не о морфологии, а о более общей дисциплине - мерономии, объединяющей морфологию, физиологию и экологию. Обращение к мерономии, а не просто к морфологии необходимо, так как фактически при выделении даже структурных компонентов организмов важна не только морфология. Ведь классификационными критериями при выделении частей служат также их физиологические и экологические функции [653]. Например, существует мнение, что цветок - не столько морфологическое, сколько функциональное понятие.
Те компоненты, которые выделяются мерономией у организмов данного таксона, называются меронами - органеллы, клетки, органы, отдельные функции, целые функциональные системы и др. Следовательно, распределяя дисперсные ископаемые части по определенным группам, мы получаем скорее мероны, а не таксоны. Чем сильнее при этом мы абстрагируемся от эвтаксонов, тем ближе наши паратаксоны к меронам. Происходит все большая мерономизация систематики.
В мерономии процедура получения меронов называется гомологизацией частей. Один из ведущих критериев гомологии - критерий положения части в целом (точнее, в архетипе). Классифицируя же дисперсные ископаемые части организмов, не всегда можно опереться на критерий положения. Поэтому нельзя отождествлять процедуру гомологизации (т. е. получения меронов) и процедуру получения паратаксонов. Выделяя паратаксоны, палеонтолог далеко не всегда может судить об отношениях сходства между однородными частями одного организма (например, судить об изменчивости листьев в пределах дерева), отличить части одного организма от частей, принадлежавших разным организмам (индивидам). Покровы, отделившиеся при линьке одного индивида, он может принять за независимые индивиды.
Провести строгую грань между эв- и паратаксонами нельзя. Некоторое сближение эвтаксонов с меронами (играющими роль паратаксонов), т. е. мерономизация, наблюдается в любой систематике, когда мы лишены возможности учесть какие-либо признаки или умышленно ими пренебрегаем. Известно много примеров выявления синонимии таксонов, оказывавшихся стадиями жизненного цикла или экологическими модификациями особей одного таксона. Например, "вид" диатомовых водорослей Moelleria antarctica Castr. оказался молодой стадией развития вида Eucampia balaustium Castr. [2336]. Установить подобные соотношения нелегко даже на современном материале, и здесь исследователю приходится опираться на косвенные соображения. Так, на основании числа попадающихся экземпляров, биометрических и гидростатических исследований было высказано предположение, что аммониты Otoscaphites puercuius (Jimbo) и Scaphites planus (Yabe) - особи разного пола одного вида [2353]. Статус обоих видов в отношении понятий эв- и паратаксон сложен и интерпретируется нелегко. Взятый порознь по отношению ко всем прочим аммонитам, каждый из видов может интерпретироваться как полноценный эвтаксон, но по отношению друг к другу эти виды - типичные паратаксоны. Следовательно, все зависит от того, в каком отношении рассматривается таксон. В одном отношении он выступает как типичный эвтаксон, в другом - как паратаксон. Эта относительность эв- и паратаксонов должна приниматься во внимание.
Палеонтологические таксоны расположатся довольно равномерно на воображаемой линии, соединяющей полноценные эвтаксоны с паратаксонами. Поэтому возьмем наиболее трудные для палеонтологии случаи, когда группировки фоссилий приближаются к паратаксономическому концу этой воображаемой линии.
Наиболее показательны палеоботанические материалы, где мерономизация таксономии была осознана еще в начале прошлого века и где отработана процедура обращения с паратаксонами, их выделения и наименования. На палеоботаническом материале особенно хорошо видны следующие основные отличия палеонтологической и неонтологической таксономии и номенклатуры [661].
1. У современных растений каждый индивид, будь то целый или представленный рассеянными частями (например, опавшими листьями и семенами), включается в один вид, род и т. д., т. е. в один таксон каждого ранга и получает единственное биноминальное название; исключения делаются лишь для гибридов.
У ископаемых растений различающиеся дисперсные части, даже первоначально принадлежавшие одному индивиду, если их прижизненная связь не наблюдается, а лишь предполагается, могут быть отнесены к разным таксонам одного ранга и получают самостоятельные биномиальные названия. Части выступают как независимые индивиды, классифицируемые независимо от их первичных взаимосвязей (табл. 7). Так, род Stigmaria объединяет ризофоры плауновидных, стволы которых относятся к родам Lepidodendron, Sigillaria и др., стробилы - к родам Flemingites, Lepidostrobus, Lepidocarpon и др. Упразднить род Stigmaria, даже зная прижизненное сочетание стволов и ризофоров, нельзя, так как невозможно поставить во взаимно-однозначное (изоморфное) соответствие виды, принадлежащие этим родам. В частности, неизвестно, с какими стволами и стробилами ассоциировали ризоформы вида Stigmaria stellata Goepp.
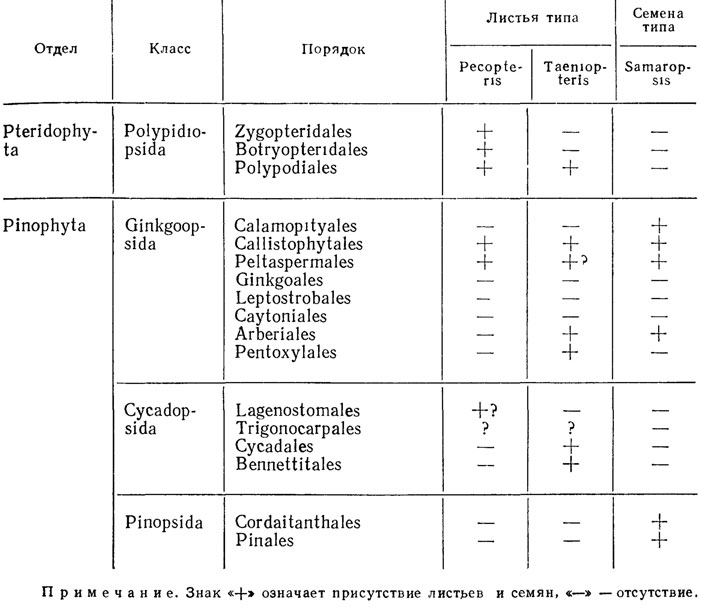
Таблица 7. Распределение дисперсных частей, относимых к форм-родам Pecopteris, Taeniopteris и Samaropsis, по надродовым таксонам высших растений
2. У современных растений все индивиды вида принадлежат одному роду, все виды рода одному семейству и т. д.
У ископаемых растений индивиды одного вида могут принадлежать разным родам и надродовым эвтаксонам (если бы растения встречались целиком), разные виды рода могут принадлежать разным семействам. Например, к виду Stigmaria ficoides (Stbg.) Brgt. относятся экземпляры, исходные растения которых могли относиться к разным семействам. Среди видов рода Pecopteris есть виды, заведомо относящиеся к разным семействам, и большое количество видов неопределенного систематического положения.
3. У современных растений каждый индивид должен быть включен в полную иерархию таксономических категорий от вида до отдела.
У ископаемых растений каждый экземпляр должен быть включен (если допускает сохранность) в вид и род; таксоны более высокого ранга могут быть неизвестны пли ими можно пренебречь. Например, вид Rhipidopsis ginkgoides Schm. иногда условно относят к порядку Ginkgoales без указания семейства, по это может быть и пельтаспермовое голосеменное пли представитель иной группы.
4. Если экземпляр современного растения не обнаруживает признаков, по которым нельзя определить его род и вид, то это не может служить основанием для выделения такого экземпляра в самостоятельный род и вид только в силу его дефектности.
Виды и роды ископаемых растений могут устанавливаться для различных степеней сохранности даже одной и той же части. Так, в зависимости от степени декортикации стволы лепидофитов, даже первично относившихся к одному виду, могут быть отнесены к разным родам (Lepidodendron, Aspidiaria, Bergeria, Knorria и др.; рис. 114).
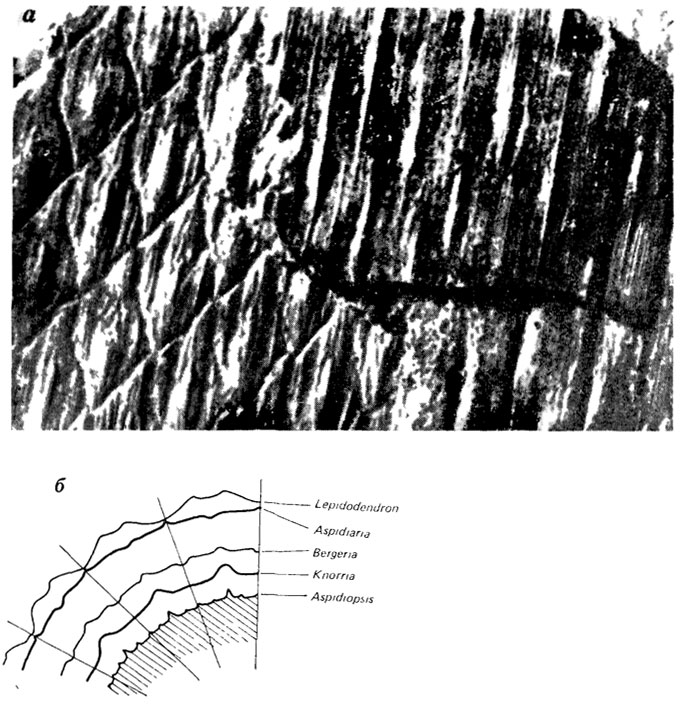
Рис. 114. Разные стадии декортикации ствола Lepidodendron и соответствующие им форм-роды. По В. Шусте. а - общин вид ствола с разными типами декортикации, соответствующими родам Aspidiaria(слева) и Knorria (справа); б - соотношение форм-подов схематически показанное па поперечном сечении ствола Lepidodendron
5. Разные онтогенетические фазы современных растений одного вида не могут выделяться в самостоятельные таксоны (кроме грибов с плеоморфным циклом).
Роды и виды ископаемых растений могут устанавливаться по разным фазам онтогенетического цикла даже одного вида. Достаточно вспомнить, что отдельные роды и виды выделяются для дисперсных спор, семян и плодов.
На основании некоторых из перечисленных пяти различий в таксономии и номенклатуре ископаемых и современных растений делались неоднократные попытки различать "полноценные" роды, орган-роды и форм-роды. Эти попытки нашли отражение в разных вариантах Международного кодекса ботанической номенклатуры. При этом не было принято во внимание, что перечисленные пять особенностей в большей или меньшей мере, явно или завуалированно, актуально или потенциально свойственны всем родам ископаемых растений. Каждый род выделяется по частям растений, всегда может обнаружиться, что разные виды одного рода относятся к разным семействам, всегда можно оспорить помещение вида в полную иерархию надродовых таксономических категорий, каждый род устанавливается для определенной онтогенетической фазы (полный онтогенетический цикл неизвестен ни у одного вымершего растения).
Перечисленные отличия могут служить для разделения эв- и паратаксонов по каждому из этих признаков и по любым их комбинациям. Для некоторого "абсолютного паратаксона" будут выполняться все вторые члены из перечисленных пар. Ясно, что палеонтология стремится приблизиться к эвтаксонам (для них нужно сочетание всех первых членов тех же пар), т. е. "демеронизировать" палеонтологическую систематику с тем, чтобы можно было интегрировать систематику всех организмов когда-либо заселявших Землю. С этой целью используется ряд приемов, на которых мы еще остановимся.
Самостоятельные и несамостоятельные признаки. Каждый палеонтолог должен знать, что конхостраки относятся к членистоногим, а двустворки - к моллюскам. Таксономическая дистанция между ними огромна. На практике же начинающий палеонтолог может легко ошибиться и принять раковины конхострак за двустворок. Опытный палеонтолог даже не станет смотреть, есть ли у данного экземпляра замок или иные признаки двустворок. Он обратит больше внимания на резкость линий нарастания, уплощенность створок, их вещество или просто будет исходить из того, что перед ним род Leaia, который, как известно, относится к конхостракам.
Палеонтологи очень часто опираются на комплекс характерных и заметных признаков, каждый из которых отдельно имеет небольшое значение, но все вместе эти признаки оказываются высоко скоррелированными с признаками очень высокого веса. Примерно то же происходит в случаях, когда палеонтолог определяет высшие таксоны, отталкиваясь от знания низших. Представим себе, что найден бивень формы и размеров, свойственных виду Elephas primigenius Blum. Достоверное определение, что это остаток плацентарного млекопитающего, а точнее, хоботного из рода Elephas, требует наблюдения широкого комплекса признаков, а не просто размеров и формы бивня. Все эти признаки могут лишь подразумеваться, "домысливаться", что и делается на основе допущения, что бивень принадлежит мамонту. Такую операцию палеонтолог проделывает постоянно. Род Pleuromeia без колебаний относится к плауновидным и вообще растениям, хотя никто никогда не видел у остатков этого рода важнейший признак растений - хлорофилл в фотосинтезирующих тканях.
"Домысливание" признаков высокого веса по наблюдению комплекса признаков невысокого веса широко применяется и при изучении современных организмов. Каждый биолог знает, что обладание клеточным ядром с ядерной мембраной и хромосомами - признак едва ли не высшего веса. Тем не менее на практических занятиях по систематике студент увидит ядро лишь у немногих организмов. Многие важнейшие таксономические признаки никогда не изучаются у большинства объектов и в то же время уверенно им приписываются. Такие постулируемые признаки назовем несамостоятельными, а действительно наблюдаемые - самостоятельными. По-видимому, не существует признаков, неизменно самостоятельных или несамостоятельных. Каждый признак может быть тем или другим в зависимости от материала, детальности и целей таксономического исследования. Казалось бы, можно хромосомы ископаемых организмов отнести к несамостоятельным признакам с большой уверенностью, однако недавно были описаны структуры, очень похожие на хромосомы, в микрогаметофитах карбонового вида рода Lepidostrobus [1368].
Возможность делать вывод о несамостоятельных признаках по самостоятельным свидетельствует о вхождении тех и других в достаточно прочный комплекс (синдром), являющийся неслучайным, т. е. закономерным. В то же время каждый признак может в отдельных случаях выходить из синдрома. Последний поэтому имеет не жесткий, а статистический характер. Отсюда и статистическая природа таксонов, различий между таксонами, а также неизбежно вероятностный характер всех таксономических утверждений [653].
Типологические экстраполяции. Любое типологическое (таксономическое или мерономическое) исследование основывается на определенных правилах выборки. Мы изучаем не всю популяцию, а только выборку, результаты исследования которой экстраполируются на всю популяцию. Из выборки первого порядка делаются выборки следующих порядков. Какие-то экземпляры подвергаются анатомическому изучению и из них могут быть изготовлены шлифы. Другие могут изучаться химическими методами. Не всегда можно применить широкий комплекс методов к одному экземпляру. Получив полную серию поперечных сечений одиночного коралла, необходимо изготовить полную серию продольных сечений из другого экземпляра.
Сложные технические приемы применяются не для отнесения результатов к единичным особям. Требуется экстраполировать на другие экземпляры изученные признаки, которые становятся для них несамостоятельными. Та же операция проделывается и в мерономических исследованиях. Обычно таксономические (с отдельных экземпляров на таксон) и мерономические (с отдельных частей на мерой) экстраполяции идут бок о бок. Поэтому разумно говорить об общем принципе типологических экстраполяции. Нетрудно видеть, что суждение о ненаблюдавшихся несамостоятельных признаках высокого веса по наблюдавшимся самостоятельным признакам малого веса - частный случай типологических экстраполяции.
Типологические экстраполяции - плавный инструмент, благодаря которому постепенно "демеронизируется" систематика, а паратаксоны приближаются к полноценным биологическим таксонам. Сходство частей в разных таксонах ограничивает типологические экстраполяции, которые неизбежно будут в разной степени вероятностными. Заметим еще, что правила и виды типологических экстраполяции остаются невыявленными и неформализованными. Поэтому они пока не поддаются математизации. Отсюда, в частности, невысокая продуктивность математических методов в палеонтологической систематике, в которой объем типологических экстраполяции особенно велик.
Принцип монотопных выборок. К числу важных несамостоятельных признаков почти всех ископаемых форм принадлежит интерфертильность внутри вида. Захороненных при копуляции животных до сих пор не обнаружено. Имеются данные о бесполом размножении (например, почковании у кораллов). Некоторые фазы полового процесса известны у ископаемых растений (проросшая пыльца в семезачатке каллистофитового Callospermarion, предполагаемое чередование поколений Zosterophyllum). Эти единичные сведения не заполняют огромного пробела в наших знаниях о репродуктивной совместимости или несовместимости организмов геологического прошлого.
Следует заметить, что и в неонтологии этот признак чаще всего относится к несамостоятельным. О нем обычно судят не по экспериментальным или наблюдаемым в природе скрещиваниям, а по косвенным свидетельствам. О межвидовой гибридизации судят по находкам особей с трансгрессирующими признаками двух видов (и делают при этом массу ошибок, принимая параллелизм в изменчивости за следы гибридизации). О репродуктивной изоляции судят по отсутствию переходных форм, размежеванию экологических ниш и (или) ареалов. Такие возможности, пусть более ограниченные, есть и у палеонтолога. В этом, в частности, и заключается смысл приложения географического и стратиграфического критериев видовой и надвидовой обособленности палеопопуляций.
Против обоих критериев было выдвинуто немало веских обвинений. Указывалось, что приложение их к таксономии приводит к логическому кругу, поскольку сначала мы обособляем таксоны по этим критериям, а потом используем те же таксоны для стратиграфических и биогеографических построений. Подобные нарекания во многом справедливы. Приходится постоянно сталкиваться с практикой, когда систематика фоссилий настолько тесно связана с их возрастом, что иной палеонтолог не берется за определение коллекции, пока геолог не укажет ему возраст вмещающих пород. Это, конечно, недопустимо.
В меньшем масштабе с тем же явлением приходится встречаться и в отношении биогеографического критерия в таксономии. Например, некоторые палеоботаники долго сохраняли независимыми роды Noeggerathiopsis и Cordaites только потому, что они приурочены к разным фитогеографическим областям.
Если разная стратиграфическая и биогеографическая приуроченность фоссилий подкрепляется пусть малозаметными, но выдерживающимися признаками, то нет оснований отвергать стратиграфическое и биогеографическое положение как дополнительный критерий самостоятельности таксонов. Более того, эти критерии должны применяться совместно, приводя в конечном счете к принципу монотопных выборок, ранее называвшихся монотопными рядами [638, 643]. Монотопные выборки применялись при изучении различных групп ископаемых растений [1076, 1709, 2250], двустворок [1934], строматолитов [541] и других фоссилий. Специфика монотопных выборок предусматривает следующее предположение: в одном захоронении или в стратиграфически и географически сближенных захоронениях больше вероятность того, что формы, близкие по самостоятельным признакам, представляют организмы, обладавшие общими несамостоятельными и весомыми признаками, в том числе и репродуктивной совместимостью. Устанавливая в монотопной выборке постепенные переходы между формами, резко различающимися по определенному признаку, мы интерпретируем эти различия как внутривидовые (рис. 115). Примерно такой же смысл имеют "биозаписи" (biorecords) Н. Хьюза [1761], нацеленные, правда, чуть ли не на полный отказ от традиционной видовой систематики, но с максимальным учетом полиморфизма в выборках.

Рис. 115. Внутривидовой полиморфизм листьев Rhaphidopteris praecursoria S. Meyen (Peltasmermales), выявленный по двум монотопным выборкам в казанских отложениях Притиманья. По С. В. Мейену. Листья изображены в разном масштабе, линейка всюду соответствует 1 см
Опытные палеонтологи всегда обращаются к монотопным выборкам, хотя приходится сталкиваться и с противоположным подходом, когда малейшие отличия служат для выделения (или определения) множества видов одного рода в одном захоронении, хотя бы эти виды и были связаны постепенными переходами. Такая практика при установлении видов ведет к их бесконечному дроблению, а при определении видов - к канонизации когда-то установленного диапазона внутривидовой изменчивости.
И то и другое нет оснований осуждать. Исследователь вправе выделять и определять таксоны, в том числе виды, так, как он считает правильным. Но все же заметим, что игнорирование принципа монотопных выборок лишает исследователя возможности выявлять внутривидовую изменчивость или корректировать ранее сложившиеся представления о таковой. Изучение современных организмов убеждает, что всем им свойственна большая или меньшая изменчивость по всем признакам. Нет оснований считать, что у организмов прошлого дело обстояло иначе. С другой стороны, имеющиеся взгляды на конкретный диапазон изменчивости могли сложиться на материале худшем, чем тот, который надо определить.
Можно назвать случаи, когда удавалось показать справедливость принципа монотопных выборок и несостоятельность пренебрежения им. Например, верхнепалеозойские кордаиты Сибири долго делились на виды по макроморфологическим признакам (пропорции, размеры, форма листа, густота жилкования и проч.). В одном захоронении нередко указывалось очень много видов одного рода. Изучение монотопных выборок приводило к иной систематике, основанной на микроструктуре листа, а из макроморфологических признаков - особенно на строении основания листа. Вывод по монотопным выборкам полностью подтвердился, когда были найдены облиственные побеги с весьма изменчивыми по форме листьями [638].
Известно, что перед палеонтологией традиционно стоит проблема восстановления прижизненной связи частей, встречающихся дисперсно. Анализ монотопных выборок помогает и в этом, в том числе в восстановлении жизненных форм растений.
Проблема возрастной множественности систем. Л. П. Татаринов [1031, с. 26] заметил: "Сам факт включения в систему позвоночных ископаемых форм сделал границы между их классами неопределенными и колеблющимися". То же можно сказать и про другие группы организмов. Так, открытие девонских прогимноспермов стерло грань между споровыми и голосеменными. Палеонтология знакомит нас с формами, связывающими не только современные, но и уже вымершие таксоны. Например, Asteroxylon связывает зостерофилловых проптеридофитов ("псилофитов") с примитивными вымершими плауновидными типа Drepanophycus. Трудность разграничения таксонов из-за таких переходных форм усугубляется тем, что в распоряжении палеонтолога часто есть лишь ограниченный круг признаков.
Решения давно возникшей проблемы, что делать с переходными формами, в которых трансгрессируют признаки разных таксонов, не существует. Неясно, возможно ли такое решение в принципе. Умозрительно проще всего проблема решается в кладистической систематике. В. Генниг [1724] отмечал, что если в ходе дифференциации вида со временем возникнут разные классы, то нет ничего страшного и противоречащего логике, если разные особи этого вида отойдут к разным классам. На практике такое, однако, не случается, поскольку с точностью до исходного вида (и тем более его части) монофилетическое происхождение высших таксонов реконструировать невозможно. Происхождение таксонов высокого ранга от единственного предкового вида лишь теоретически постулируется кладистической систематикой и никогда не было продемонстрировано на конкретном материале. По этой и ряду других причин "практика кладизма далеко не так радикальна, как его теория" [526, с. 123]; см. также [1036, 1037].
Другие таксономисты предлагают иной выход из положения, считая, что сам принцип филогенетической систематики, покоящейся на феномене возрастающей во времени дивергенции (постепенном расхождении таксонов в признаковом пространстве), требует смены ранга одного и того же таксона во времени. Имеется в виду, что, скажем, современные классы когда-то были более сходными и составляли лишь разные отряды, еще раньше - семейства и т. д. Соответственно предлагалось построение разных систем для разных эпох. Пример такого подхода на палеоботаническом материале показал Р. Дабер [1479]. Он предложил самостоятельные системы для эпох, которые он выделил по этапам эволюции растительного мира и назвал Psilophyticum (ранний и средний девон), Pterophyticum (поздний девон), Pteridospermophyticum (ранний карбон), Варисская эпоха (остальная часть карбона и ранняя пермь), Mesophyticum (поздняя пермь - ранний мел), Kanophyticum (альб - квартер). Пока никто не воспользовался предложенной системой.
Есть "за" и "против" такого подхода к систематике. "За" могут быть высказаны следующие соображения. Во-первых, мы действительно сталкиваемся в геологической летописи с таксонами, которые достаточно ясно отграничении друг от друга на определенном временном срезе, но совмещают признаки таксонов, появившихся раньше или позже. Некоторые таксоны в исторической перспективе приобретают статус переходных форм и не могут быть уложены в общую систему без серьезных натяжек. Строя системы для определенных временных интервалов, мы преодолеваем эти трудности. Во-вторых, как бы мы ни относились к историческим толкованиям системы организмов и даже ее отождествлению с филогенетическим древом (против этого обоснованно высказывались многие теоретики), огромная, если не ведущая роль унаследованных сходств в общей массе внутритаксономических сходств представляется очевидной. Признав это, мы вынуждены согласиться и с тем, что таксономические структуры не могут рассматриваться вне связи с исторической перспективой, а из этого следует принципиальная возможность значительной дивергенции и повышения ранга таксона в ходе его эволюции.
Соображения "против" тоже можно свести к двум положениям. Во-первых, неизвестно, как и на какие отрезки следует членить историю какой-то крупной группы или всего органического мира, чтобы затем для каждого отрезка строить самостоятельные системы. Стратиграфические подразделения связаны переходами (стратоэкотонами). Возникает вопрос, как быть с переходными зонами между отдельными этапами эволюции или седиментации. Вместо трудности с отражением в структуре системы форм, переходных в признаковом пространстве, мы получим не меньшие трудности в отражении переходных форм в земном пространстве - времени. Во-вторых, проведение общего принципа самостоятельных систем для разных отрезков геологической истории потребует аналогичного подхода и в мерономии (например, в морфологии). Тогда надо будет создавать и самостоятельный язык для описания организмов разных эпох. Вместо интеграции наших знаний об органическом разнообразии мы рискуем столкнуться с дезинтеграцией этих знаний по разным отрезкам времени. Возрастную множественность систем придется как-то снова интегрировать, а как это сделать, непонятно.
Противоположение аргументов "за" и "против" в конечном счете связано с недостаточностью существующих формальных средств для отражения того разнообразия организмов и той упорядоченности в нем, которые призвана отразить систематика (точнее, типология в оговоренном выше смысле). Система дискретных таксонов ограниченного числа рангов грубо упрощает то, что исследователь действительно видит в природе. Необходимы какие-то иные средства для адекватного отражения этого разнообразия в системе таксонов, меронов и архетипов. На поиск этих средств и должно быть направлено внимание исследователей. Отчасти к этому ведут и попытки ввести возрастную множественность систем. Насколько перспективны эти попытки - покажет будущее.
Сказанное о несоответствии формальных средств, используемых для выражения наблюдаемого разнообразия, самому этому разнообразию - старое противоречие действительности и образа, а также формы и содержания. Из этого же вытекают и прочие проблемы. С ними так или иначе связан любой систематик, будь то палеонтолог или неонтолог. В палеонтологии эти проблемы лишь ярче выступают, поскольку здесь приходится иметь дело с дефектным материалом и поскольку в палеонтологической таксономии разнообразие организмов с неизбежностью должно совмещаться с координатой времени. Поэтому обсуждение общих принципов палеонтологической систематики имеет большое методологическое значение для биологии в целом. Ископаемый органический мир - тот пробный камень, на котором можно успешно проверять принципы и методы на общебиологическую значимость. Ни один принцип или метод не может считаться универсальным и, следовательно, фундаментальным, если он неприложим к трудным случаям. Именно такой трудный случай и предоставляет теоретической биологии, в том числе и теоретической систематике, палеонтология.
Проблема свертки таксономического разнообразия. Задача каждой науки - свести все разнообразие изучаемых ей объектов в некую компактную и легко обозримую систему понятий, которые должны быть связаны друг с другом таким образом, чтобы одни понятия легко выводились из других по определенным правилам вывода. Связанные таким путем понятия не приходится запоминать поодиночке. Эту задачу преследует и систематика организмов, в том числе и ископаемых. Ее основные понятия - все те таксоны, которые она описывает. В некоторой степени систематика удовлетворяет указанному требованию. Зная соподчинение таксонов и признаки высших из них, мы уже можем не запоминать многих признаков низших таксонов. Так, нет необходимости помнить о наличии позвоночника у каждого вида позвоночных, поскольку наличие позвоночника - признак таксона высокого ранга. Классическая систематика, построенная по иерархическому принципу, всегда наделяет таксоны низшего ранга признаками таксонов более высокого ранга. Однако она не позволяет судить о признаках таксона того же ранга. Поэтому, зная признаки позвоночных и отвлекаясь от таксона более высокого ранга, мы ничего не можем сказать о признаках моллюсков. "Горизонтальное разнообразие" (в пределах таксонов одного ранга) необходимо просто фиксировать и запоминать, перебирая таксон за таксоном, каждый из которых получает независимую характеристику. Когда дело доходит до видов и разновидностей, приходится оперировать миллионами таксонов, запоминая их признаки порознь. Нечего и говорить о компактности и обозримости складывающейся структуры понятий.
Правда, систематики давно заметили, что сходство между таксонами не вполне соответствует иерархической древовидной структуре с независимыми конечными ветвями. В дереве сходств обнаруживаются поперечные связки, соединяющие порой весьма удаленные ветви. Еще К. Линней отмечал, что система организмов имеет форму сети, а не дерева. Впервые наиболее детально и на большом материале, включая палеонтологический, это явление подробно рассмотрел Э. Д. Коп [291]. Он описал параллельные изменения органов в разных таксонах, а сами параллельные ряды форм назвал "гомологическими группами". Соответственные члены разных рядов он назвал "гетерологическими". Опираясь на это явление, уже можно что-то сказать о таксоне того же ранга, если у этого таксона есть тот же тип органа, что и у изучаемого таксона.
Параллелизм в разнообразии разных таксонов отмечался многими исследователями и обычно использовался в борьбе с эволюционизмом дарвинистского толка. Упорядоченность в изменчивости выдвигалась в качестве опровержения тезиса случайной, ненаправленной изменчивости. Между тем на это же явление, хотя и на низком таксономическом уровне, обращал внимание и Ч. Дарвин, обозначив его термином "аналогическая изменчивость".
Ч. Дарвин и большинство его последователей трактовали параллелизм в изменчивости как дополнительное свидетельство филогенетического родства (чем ближе таксоны, тем полнее параллелизм в изменчивости; справедливым признавалось и обратное утверждение). Если связь с родством казалась сомнительной, то сходство в изменчивости связывалось с внешними факторами или сходством функционирования (конвергенция). Другие исследователи, преимущественно противники дарвинизма, видели в параллелизме свидетельство определенных закономерностей в изменчивости, которые могут проявляться и безотносительно к родству, внешним факторам и сходству в функции. Эти две точки зрения удержались до наших дней.
После работ Э. Д. Копа наиболее обстоятельно явления параллелизма были рассмотрены в работах Н. И. Вавилова [132]* и Н. П. Кренке [533], однако если первые получили широкую известность, то вторые, хотя явления параллелизма были в них проанализированы глубже, прошли почти незамеченными. Отчасти используя терминологию Э. Д. Копа, Н. И. Вавилов говорил о "законе гомологических рядов в наследственной изменчивости" и иллюстрировал его материалами преимущественно по культурным растениям. "Виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм и у других видов и родов. Чем ближе генетически расположены в общей системе роды и линнеоны, тем полнее сходство в рядах их изменчивости. Целые семейства растений в общем характеризуются определенным циклом изменчивости, проходящей через все роды и виды, составляющие семейство" [132, с. 35]. "Вместо случайных поисков исследователь ищет определенных форм, предвидя их существование на основании сходства изменчивости с ближайшими известными видами и родами" (там же, с. 51-52).
* (Первые работы Н. И. Вавилова на эту тему относятся к 1920 и 1922 гг. Ссылки на статьи Н. И. Вавилова даются по изданию 1967 г.)
Распределение признаков по родам и видам Н. И. Вавилов представил в виде наглядных формул:
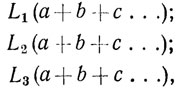
где L - радикалы, т. е. признаки, отличающие виды друг от друга, a, b, c... - признаки, одинаково варьирующие в разных видах. Количество признаков, которые можно выделить в радикал, обычно очень невелико.
В работах Н. И. Вавилова не получили должного внимания два обстоятельства, на которые обоснованно сделал упор Н. П. Кренке. Во-первых, признаки в скобках брались Н. И. Вавиловым в виде некоего списка, порядок членов в котором не имел существенного значения. Однако члены этого списка могут быть упорядочены по некоторому правилу преобразования данного признака. Этот упорядоченный набор признаков (или состояний одного признака) был назван рефреном [653, 1952]. Именно наличие рефренов и дает возможность тех прогнозов, о которых пишет Н. И. Вавилов. Во-вторых, признаки радикала тоже подвержены изменчивости и входят в рефрены. Правда, изменчивость радикала наблюдается редко, для этого нужны огромные выборки, иногда из десятков тысяч экземпляров. Поэтому изменчивость радикала трактуется как уродство и мало интересует систематиков. Будучи "уродством" в одном таксоне, признаки радикала обычно встречаются в качестве нормы в другом таксоне. На это явление, которое Н. П. Кренке назвал "законом родственных отклонений", а С. В. Мейен "правилом Кренке" [653, 1950], исследователи обращали внимание еще в XVIII в. [1135]. Как и в случае рефренов, диапазон родства, при котором проявляется правило Кренке, не имеет ограничений.
Подчиняя изменчивость признаков определенным правилам преобразования, рефрены позволяют "свертывать" разнообразие, наблюдаемое в пределах одного таксона и в разных таксонах. По существу, обращение к рефренам, их систематическое выведение на всем биологическом материале - пока единственный реальный путь номотетизации систематики, т. е. введения в систематику закономерностей, не отличающихся по статусу от тех, которые описываются точными науками. Иное дело, что пока преобразования меронов в рамках рефренов не часто удается описать с помощью математических средств.
Исследователи параллелизма на палеонтологическом материале вместо построения рефренов с ясно сформулированными правилами преобразования меронов чаще всего ограничиваются построением решеток комбинирующихся признаков. С помощью таких решеток иллюстрируются приложения "гомологических рядов" к ископаемому материалу. Наиболее обстоятельно это сделано в работах А. Ю. Розанова [873], который положил "гомологические ряды" в основу упорядочения систематики правильных археоциат (рис. 116). В некоторых случаях он показал последовательность форм, которая может рассматриваться как рефрен. Вслед за А. Ю. Розановым этот же подход был применен другими исследователями археоциат [372, 768, 1486, 1487], причем в методическом отношении следует выделить последнюю из указанных работ, в которой наряду с обычными для палеонтологии приемами использован математический метод распознавания образов - алгоритм "Форэль-1" (Формальный элемент-1). Известны работы по приложению гомологических рядов к фораминиферам [578, 839], кораллам и другим группам ископаемых. Рефрены, демонстрировались на палеоботаническом материале [1950, 1952].
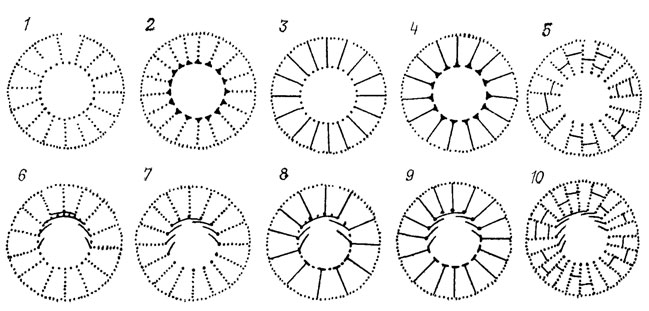
Рис. 116. Гомологические ряды археоциат семейств Ajacicyathidae и Cyclocyathidae. По А. Ю. Розанову. 1 - Aldanocyathus; 2 - Robustocyathus; 3 - Ajacicyathus; 4 - Mongolocyathus; 5 - Sibirecyathus; 6 - Kellericyathus; 7 - Gordonicyathus; 8 - Denaecyathus; 9 - Compositocyathus; 10 - Gordonifungia
С принципиальной точки зрения гомологические ряды могут быть составлены на всех систематических уровнях. Но в палеонтологии выбор уровня, на котором наиболее последовательно могут быть прослежены эти ряды, представляет собой немалую сложность, обусловленную возможными (практически довольно частыми) изменениями соподчинения выделяемых таксонов в зависимости от степени изученности данной группы ископаемых при отсутствии надежного обоснования их действительного ранга.
Выше уже говорилось, что вслед за Ч. Дарвином многие исследователи связывают параллелизм в изменчивости с единством происхождения соответствующих групп. Именно так трактовал свои ряды и Н. И. Вавилов: "Дело не только в параллелизме, во внешнем сходстве, а в более глубокой эволюционной сущности наследственной изменчивости у родственных организмов. Всеобщность этого явления прежде всего определяется генетическим единством эволюционного процесса и происхождения, родством" [132, с. 48].
Безусловно, в общем случае чем ближе филогенетическое родство между таксонами, тем полнее проявление параллелизма в изменчивости. Однако известны и обратные случаи [1950]. Хороший пример дают рефрены, построенные для листьев папоротников и тригонокарповых голосеменных. Эти рефрены несравненно полнее, чем те, которые можно построить для листьев тригонокарповых и пинопсидных голосеменных. Раньше полагали, что тригонокарповые и другие "птеридоспермы" произошли от папоротников; на основании этого можно было бы согласовать полноту рефренов с филогенетической близостью. Теперь известно, что папоротники и птеридоспермы не связаны прямым филогенетическим родством. Обе группы независимо произошли от тримерофитов, от которых взяли начало через прогимноспермов и прочие голосеменные. Хорошо известен параллелизм в поведении высших позвоночных и насекомых. Связывать этот параллелизм с общностью происхождения тех и других - значит, ничего не сказать. Попытки связать рефрены с общностью внешней среды или функционирования чаще всего тоже оказываются безуспешными, так как их невозможно конкретизировать.
Поведение признаков в эволюционирующих таксонах обычно описывается понятиями дивергенции, конвергенции и параллелизма. Привлечение понятий рефрена и правила Кренке существенно конкретизируют понятие параллелизма, поскольку в рассмотрение вводятся упорядоченность членов внутри параллельных множеств форм (рефренов) и частотные характеристики повторяющихся членов (правило Кренке). При конвергенции отдельные признаки таксонов сближаются в признаковом пространстве или же происходит сближение частот общих признаков (на эту разновидность конвергенции обычно не обращают внимания). Точно так же и дивергенцию можно представить как расхождение признаков таксонов в признаковом пространстве и как расхождение частот общих признаков. Имеются понятия и для различных сочетаний дивергенции, конвергенции и параллелизма - транзитивный полиморфизм, присутствие синтетических типов в основании филогенетических ветвей и др. [653, 1955].
В целом получается следующая модель соотношения таксонов и признакового пространства [661]. Можно представить признаковое пространство как многомерную панель, несущую лампочки-признаки, зажигающиеся при проявлении признака у индивидов данного таксона. Дискретные таксоны в той упрощенной характеристике, которую им обычно дают определители, будут представлены пятном постоянно горящих лампочек, а тераты (уродства) - лампочками, редко вспыхивающими вдали от основного пятна. Таксоны, детально изученные на огромных выборках (известны случаи, когда выборки исчислялись сотнями тысяч особей), обнаруживают изменчивость чуть ли не по всем признакам. Соответственно такие таксоны будут представлены более крупным пятном горящих лампочек, ни одна из которых не будет гореть постоянно, а лишь с большей или меньшей частотой (или с разной яркостью, если изменчивость континуальна). Переход от таксона к таксону (с учетом рефренов и правила Кренке) будет происходить за счет вспыхивания новых лампочек и за счет перераспределения частот вспыхиваний (или яркости свечения) в одном и том же множестве лампочек. Строго говоря, каждый рефрен скорее можно представить не набором лампочек, а нитью, у которой в одном таксоне светятся (причем с варьирующей яркостью) одни участки, а в другом - те же или другие участки.
Вполне допустимо предположение, что сложная картина изменчивости, наблюдающаяся у хорошо изученных таксонов, свойственна всем таксонам. Тогда типология организмов во всей ее полноте предстанет в виде сложной системы размытых и прихотливо переплетающихся пятен на нашей воображаемой панели. Таксономическая эволюция организмов будет выглядеть как перемещение пятен и как разрастание самой панели. Различить оба процесса на практике, видимо, можно лишь в масштабе геологической летописи. Так называемая микроэволюция демонстрирует лишь перераспределение частот в проявлении признаков и ничего не говорит об их появлении. Поэтому относить учение о микроэволюции к числу собственно эволюционных преждевременно.
Задачу свертки разнообразия в этой модели можно представить как умение управлять вспыхиванием и интенсивностью горения лампочек с помощью лаконичных команд, а также как умение описать и саму панель, и разнообразие световых пятен-таксонов на ней наиболее экономным способом. Пока мы не умеем делать ни того, ни другого, а лишь наметили отдельные регулярности в этой воображаемой очень сложной панели.
Свертка разнообразия организмов требует разработки соответствующих понятийных (семантических) и знаковых (семиотических) средств. Имеющиеся средства вроде филогенетических древес, дендрограмм, различных графиков, линиями изображающих соотношение таксонов или отдельных аспектов их преобразования, здесь явно недостаточны (рис. 117, 118).
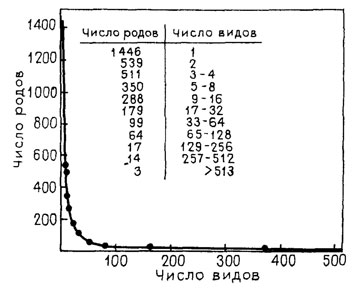
Рис. 117. Пример вогнутой кривой - число видов в 3510 родах долгоносиков семейства Curculionidae. По Киссингеру, из Э. Майра
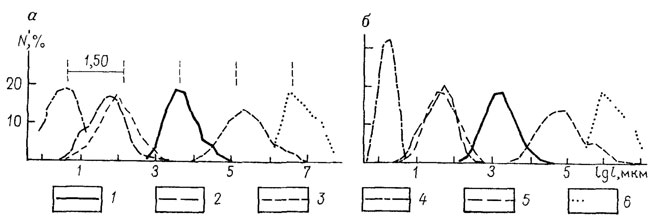
Рис. 118. Процентное распределение числа N видов главнейших групп организмов пелагиали Мирового океана в зависимости от максимальных линейных размеров. По Л. Л. Численко. (а) и приведенных линейных размеров l (б): 1 - Crustaceae; 2 - Flagellata; 3 - Pisces; 4 - Bacteriae; 5 - Diatomea; 6 - Cetacea
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'