
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
4.6. Политипическая концепция вида в палеонтологии (Л. А. Невесская)
Для теоретической палеонтологии, цель которой - изучение процессов видообразования, филогении и эволюции органического мира геологического прошлого, проблема вида как основной элементарной единицы филогенетической систематики и основного этапа эволюционного процесса, качественно отличного от других этапов, имеет огромное значение.
Практические задачи палеонтологии, тесно связанные с решением проблем стратиграфии, палеогеографии, палеобиогеографии и реконструкции условий образования полезных ископаемых, также требуют определения видов как естественных объективных единиц, характеризующих отдельные стратиграфические, палеогеографические и палеобиогеографические подразделения и определенные обстановки прошлого.
Существуют две основные концепции вида - монотипическая, или типологическая, и политипическая, или популяционная. Первая исходит из того, что сущность вида абсолютна и постоянна, а изменчивость - лишь несовершенное выражение этой сущности. При принятии этой концепции вид будет представлять собой случайную совокупность индивидов, имеющих основные свойства "типа" вида, т. е. экземпляра, выбранного в качестве стандарта. Степень близости или отличия от "типа" определяется исследователем субъективно уклоняющиеся от него экземпляры выделяются обычно в качестве разновидностей, которые при описании часто оказываются за пределами данного вида, или же в качестве других морфологических "видов" (морфовидов). В этом случае в одной популяции могут быть выделены несколько таких морфологических "видов", один из которых будет включать наиболее часто встречающиеся варианты вида, а другие - группы крайних экземпляров ряда изменчивости. Другая концепция - политипическая, или популяционная, исходит из того, что вид представляет собой генетически, экологически и физиологически единое целое и обладает общим генофондом, слагающимся из генофондов всех составляющих вид скрещивающихся популяций. Именно единство и взаимосвязь генофонда вида лежат в основе его объективности [1935]. Вид характеризуется определенными морфологическими, физиологическими, экологическими и другими признаками.
Каждый вид обладает определенными биологическими свойствами, которые обеспечивают адаптацию его к абиотическим условиям существования и возможность существовать с другими видами и особенно - с потенциальными конкурентами [1935]. Стабильность вида поддерживается сложной гомеостатистической системой, общей для всего вида и обеспечивающей, с одной стороны, единство вида, а с другой, - его дискретность. Эти механизмы определяют требования и выносливость вида в различных условиях среды.
В качестве критерия вида для организмов с половым размножением среди всех этих признаков и свойств выбрано наличие дискретности с другими видами, обусловленной репродуктивной изоляцией составляющих вид популяций от популяций всех других видов, т. е. наличие нескрещиваемости с другими видами [1933, 1934, 1935]*.
* (В настоящее время известно довольно много случаев гибридизации, в результате чего возникают диплонты, триплонты и так далее, причем некоторые из них способны к размножению. Тем не менее, вероятно, эти случаи представляют собой исключения и не обесценивают критерий репродуктивной изоляции.)
Проблема вида в биологии обсуждалась в последние десятилетия многими биологами [375, 376, 594, 596, 1008, 1054, 1058 и др.]. Каждый вид, как правило, состоит из многочисленных популяций, в определенной степени связанных между собой. Каждая такая местная популяция служит основной естественной единицей существования, приспособления и воспроизведения вида. Индивидуум содержит лишь небольшую часть генного фонда популяции в течение краткого периода времени и в силу этого не может быть основной единицей эволюции. Только популяции - воплощение фонда генов всех ее членов, единство которого обеспечивается свободным скрещиванием особей внутри популяции, может служить основой новых генных комбинаций и благодаря этому главной единицей эволюции [1189, 1191, 1409, 1935].
Одним из основных свойств каждой популяции является ее изменчивость, обеспечивающая исходный материал для формо- и видообразования, а следовательно, для эволюции. Поэтому изучение изменчивости популяций служит основой большинства эволюционных теорий и гипотез о видообразовании. Выделяются два основных типа изменчивости [594, 1105, 1935]: индивидуальная, или внутрипопуляционная, которая характеризует различия между индивидуумами одной популяции, и групповая, или межпопуляционная, отражающая различия между популяциями или группами популяций. Как индивидуальная, так и групповая изменчивость могут проявляться в качестве генетической и фенотипической изменчивости.
Палеонтолог (да и зоолог тоже) обычно имеет дело лишь с фенотипической (видимой) изменчивостью, которая тем не менее связана и в какой-то мере отражает генотипическую изменчивость.
Взаимоотношения гено- и фенотипической изменчивости, как и отношения генотипа и фенотипа достаточно сложны. Фенотип представляет собой совокупность признаков и свойств, являющуюся результатом проявления его наследственной конституции (генотипа) в конкретных условиях среды. Таким образом, фенотипическая изменчивость частично будет отражать имеющуюся в популяции генетипическую изменчивость, а частично - возникать за счет влияния среды обитания на фенотипы, составляющие популяцию; эта часть изменчивости не передается по наследству [2287]. Генотипическая изменчивость может быть значительно больше фенотипической, но может иметь место и обратное соотношение, особенно при разнообразных условиях существования популяции или популяций [1935]. Поэтому следует всегда иметь в виду существование потенциальной генотипической изменчивости, которая может проявиться в фенотипической изменчивости, особенно при изменении условий существования.
Как индивидуальная, так и групповая изменчивость могут иметь разные формы проявления. Рассматривая разнообразие форм изменчивости, Н. П. Парамонова [810] дала следующую их группировку: 1) по характеру исследуемых признаков - морфологическая (структурная), физиологическая (функциональная), этологическая (изменчивость поведения) и др.; 2) по характеру проявления признаков - непрерывная (количественная, рядовая) и прерывистая (качественная, альтернативная); 3) по связи с различными стадиями онтогенеза (возрастная) - изменчивость на ранних или поздних стадиях; 4) по наличию или отсутствию связи с полом; 5) по характеру связи между различными признаками - коррелятивная (корреляционная, соотносительная, связная, сопряженная, зависимая) и некоррелятивная; 6) по связи с условиями среды - экологическая (биотопическая) и географическая (топографическая); 7) по характеру проявления в пространстве - клинальная (непрерывная) и дискретная (прерывистая); 8) по характеру проявления во времени (хронографическая) - направленная и ненаправленная; 9) по связи с унаследованностью - наследственная и ненаследственная. В понятие изменчивости не должны включаться различия признаков или свойств, у различных индивидуумов одного вида, связанные с возрастными и сезонными изменениями и половым диморфизмом.
Особо необходимо отметить внутривидовую изменчивость, выражаемую через дискретные признаки, или фены. Фены - это генетически обусловленные отдельные признаки фенотипа, характеризующиеся простой элементарностью, т. е. неразложимостью на составные части (без потери качественного своеобразия), альтернативностью и дискретностью. Выделение фенов в общей фенотипической изменчивости вида и оценка их встречаемости в различных внутривидовых группировках помогают решению проблем внутривидовой систематики, морфологии, экологии и микроэволюции. По распространению отдельных редких фенов или их комбинаций можно прослеживать пути формообразования и миграций [1232, 1233]. При исследовании изменчивости, как правило, возникает вопрос о ее таксономическом значении, т. е. о ранге внутривидовых категорий.
Как уже говорилось, вид обычно состоит из многих популяций, которые могут группироваться во внутривидовые категории. Такой вид называется политипическим. Основа классификации внутривидовых категорий была заложена А. П. Семеновым-Тян-Шанским [942]. Первое место среди групповых (межпопуляционных) категорий занимает подвид (или географическая раса), который состоит из группы локальных популяций, имеет определенный ареал отличается от других подвидов относительно устойчивыми наследственными морфофизиологическими признаками [1935]. В качестве негеографической внутривидовой групповой категории была предложена морфа, или экологическая морфа, которая представляет собой локальную популяцию (или группу популяций), адаптированную к местным условиям и отличающуюся особыми морфологическими признаками. Для морф характерны отсутствие определенного ареала и нестойкость их отличительных морфофизиологических особенностей, поэтому при смене условий эти признаки могут быть легко потеряны или изменены. Подвиды и морфы охватывают целые популяции и группы популяций. Выделяются внутрипопуляционные единицы, например, формы, аберрации, уродства. Формой обычно называют морфологически различные группы особей внутри одной популяции, образование которых часто обусловлено полиморфизмом [1935], а значительные индивидуальные отклонения выделяются в качестве аберраций и уродств. Множественность выделяемых внутривидовых группировок свидетельствует о сложности структуры вида, которая неодинакова у разных видов.
Из всех внутривидовых категорий только подвид является систематической единицей, узаконенной правилами зоологической номенклатуры. Он имеет триноминальное название. В определении других внутривидовых категорий (морфа, форма и др.) имеются значительные расхождения.
Политипическая концепция, рассматривающая вид в динамике его развития и в конкретных формах его существования, в то же время способствует упорядочению систематики. Биометрические методы, которые применяются для изучения изменчивости, одновременно позволяют более объективно сравнивать популяции для установления их различий и таксономического ранга.
Что касается понимания вида в палеонтологии, то до 50-х годов нашего века в палеонтологических работах господствовала типологическая концепция, хотя уже в 40-х годах советские ученые указывали на необходимость отказа от выделения чисто морфологических видов [108, 1100]. Особенно очевидно неприемлемость типологической концепции была выявлена в рамках дискуссии о проблеме вида в 50-60-х годах. В ряде работ было продемонстрировано, что при использовании типологического метода скрывается одно из основных свойств организмов - их изменчивость, так как промежуточные формы отбрасываются или включаются в наиболее сходные с ними "виды". Выделенные типологическим методом "виды" искусственны и не дают возможности, с одной стороны, выявить ход эволюционного процесса, а с другой - не могут служить полноценными индикаторами обстановок прошлого, так как при их выделении не учитывается экологическая изменчивость [275, 2003 и др.]. С 60-х годов политипическая концепция вида начала завоевывать первенство в палеонтологии и в настоящее время может считаться господствующей.
При применении политипической концепции вида в палеонтологии исследователь прежде всего сталкивается с основным вопросом: может ли быть применим "критерий нескрещиваемости" к вымершим видам, т. е. можно ли считать ископаемые ("палеонтологические") виды равноценными современным ("неонтологическим") видам. Рассмотрению этой проблемы были посвящены многие работы, в которых высказывались различные точки зрения. Некоторые, хотя и немногие, исследователи защищали необходимость придерживаться в палеонтологии типологической концепции вида, в то же время подчеркивая произвольность и субъективность морфологических видов [55, 1281, 1829]. Ряд палеонтологов, принимая политипическую концепцию, в то же время указывает на специфичность палеонтологического материала и приходит к выводу о субъективности палеонтологического вида [1257, 1632, 1693 и др.]. В связи с этим делались попытки разделить биологический (неонтологический) и палеонтологический виды основываясь на том, что первый имеет распространение только в пространстве, а второй - также и во времени. Для палеонтологического вида были предложены даже особые названия: палеовид, или палеонтологический вид [555], хроновид [1632, 2154, 2376, 2483], сукцессиональный вид [1779], или фратрия [237, 1055], который объединяет популяции сегмента эволюционной линии, существовавшие в течение определенного промежутка времени, и состоит из ряда последовательных голоморфовидов; каждый из последних равен неонтологическому виду.
Против такого противопоставления палеонтологического вида биологическому активно выступали как биологи-неонтологи [1934], так и многие палеонтологи [275, 292, 604, 862, 887, 1211, 2154 и др.].
Система современных видов, как подчеркнули В. И. Громов [275] и В. Е. Руженцев [887], также учитывает фактор времени, так как большинство современных видов уходит корнями в геологическое прошлое, поэтому существуют достаточные основания считать, что вид в палеонтологии не имеет принципиальных отличий от вида в неонтологии.
О степени репродуктивной изоляции можно судить по степени морфологических отличий, так как в большинстве случаев существует определенная корреляция морфологических и генетических различий, причем первые, являясь следствием репродуктивной изоляции, как правило, вторичны [1935, 2287]. Морфологические отличия вида от других видов - продукт его биологической уникальности. В то же время чисто морфологический критерий неприменим из-за наличия морфологических различий экземпляров, относящихся к одному виду (возрастные различия, половой диморфизм, экологические морфы и пр.). Поэтому переоценка данного критерия может привести к выделению морфологических, "видов", которые часто оказываются внутрипопуляционными вариантами [275, 1015, 1632, 1779, 2003 и др.].
Применение морфологического критерия (включая изучение изменчивости) только совместно с биогеографическими, палеоэкологическими и хронологическими (стратиграфическими) данными делает решение вопроса о видовом статусе ископаемых форм достаточно достоверным.
Все изложенные положения о популяциях и их изменчивости для современных видов применимы и к вымершим видам. На палеонтологическом материале можно изучать индивидуальную и групповую изменчивость, различные формы изменчивости, а также возрастные и сезонные изменения и половые различия.
Выявление разных типов изменчивости имеет значение не только для решения теоретических проблем, но и для практических целей палеонтологии: при разработке детальной биостратиграфии, для палеогеографических и палеобиогеографических исследований, для реконструкции обстановок прошлого.
Для исследования изменчивости древних организмов необходимы достаточно хорошая сохранность материала, наличие выборок определенного объема и приуроченность этих выборок к слоям небольшой мощности, положение которых в разрезе точно установлено. Только если образцы относятся к одному и тому же маломощному слою одного или нескольких близко расположенных местонахождений, можно с достаточной долей условности говорить об ископаемой популяции.
Ископаемая популяция носит "усредненный" характер будучи представлена совокупностью экземпляров последовательных поколений [395].
Для прослеживания возможных изменений размаха и характера изменчивости во времени необходимо иметь выборки экземпляров одного вида из последовательных слоев непрерывного разреза, отвечающего достаточно длительному промежутку времени. Для изучения изменчивости предложен биометрических методов, основанных на исследовании распределения частот значения отдельных признаков или их отношений [1105, 1461, 1462, 1632, 1778, 1798, 2003, 2145, 2146, 2149, 2169, 2283, 2347].
Поскольку изменчивость признаков внутри популяции непрерывная, это распределение частот выражается одномодальной вариационной кривой (рис. 123) - одномодальные кривые 1-4 представляют собой каждая одну выборку. Поэтому наличие одномодальных кривых для всех подвергшихся изучению признаков для какой-либо выборки свидетельствует, что мы, скорее всего, имеем дело с одной популяцией.
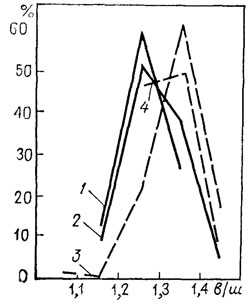
Рис. 123. Кривые изменчивости отношения высоты раковины к ее ширине для плиоценовых гастропод Viviparus abchasicus Tab. Эвксинского бассейна. По В. Я. Табояковой. 1 и 2 - две выборки раковин типового подвида; 3 и 4 - то же, для подвида V. abchasicus moquicus Tab.
При наличии морфологической прерывистости можно предполагать наличие в выборке представителей двух популяций (при исключении возможности внутрипопуляционной прерывистой изменчивости, полового диморфизма, наличия различных возрастных стадий), которые, будучи симпатрическими, скорее всего, должны относиться к различным видам [2003, 2346]. Нужно, однако, помнить, что одно статистическое различие не может служить основой для определения таксономического ранга популяции или группы популяций. Необходим также анализ географического и стратиграфического распределения изучаемой категории с привлечением палеоэкологических данных [395].
Определив границы изучаемого вида на основании имеющегося палеонтологического материала из одного слоя одного или близких местонахождений, можно судить о внутрипопуляционной (индивидуальной) его изменчивости; сравнивая же выборки из многих одновозрастных местонахождений или из последовательных стратиграфических подразделений, можно выявить межпопуляционную (групповую) изменчивость в пространстве или во времени.
Следует подчеркнуть, что выявление размаха изменчивости отдельных количественных признаков и дальнейшее сравнение амплитуды изменчивости у разных внутривидовых категорий одного вида и разных видов возможны только в случае, если выполнена хотя бы элементарная статистическая обработка полученных данных (вычисление средней величины, среднего квадратического отклонения, коэффициента вариации). Путем стандартных статистических методов можно по сравнительно небольшому числу экземпляров (выборке) оценить изменчивость многочисленных особей, составляющих популяцию, так как изменчивость не бесконечна [2003].
Изучая изменчивость, мы сталкиваемся, как и для современных видов, с вопросом о внутривидовых категориях и их ранге в каждом конкретном случае. Для ископаемых видов применяются те же категории, что и для современных, т. е. подвид и морфа для групповой изменчивости и форма, аберрация, уродство - для индивидуальной изменчивости [432, 604, 862, 1015, 1211]*.
* (Морфы, формы и другие внутривидовые категории можно обозначить латинскими названиями (longa, curta и проч.), буквами (α, β, a, b и проч.) или русскими названиями (длинная, короткая и проч.).)
Необходимо специально остановиться на категории подвида, в понимании которого среди палеонтологов существуют разногласия. Подвид в биологии - географическая категория, однако в палеонтологии он применяется и как хронологическая категория для группы популяций, имевших определенный временной "ареал" (рис. 124). В. Е. Руженцевым [887] было показано, что биологических различий между географическими и хронологическими подвидами не существует, и разграничение их обычно практически невозможно, если различные подвиды найдены в разных районах и нельзя точно определить их одновременность. Даже при наличии серии подвидов из последовательных слоев отдельные подвиды могли быть географическими расами, замещавшими друг друга из-за климатических или каких-либо других изменений условий существования [1806, 1935, 2376]. Поэтому в определение подвида следует включать фактор времени и тогда подвидом называть группу популяций, географически или хронологически обособленную от других аналогичных групп популяций того же вида, отличающуюся вследствие этого определенными, передающимися по наследству морфологическими особенностями, но связанную с другими подвидами переходными формами в местах контакта [604, 605, 862, 887].
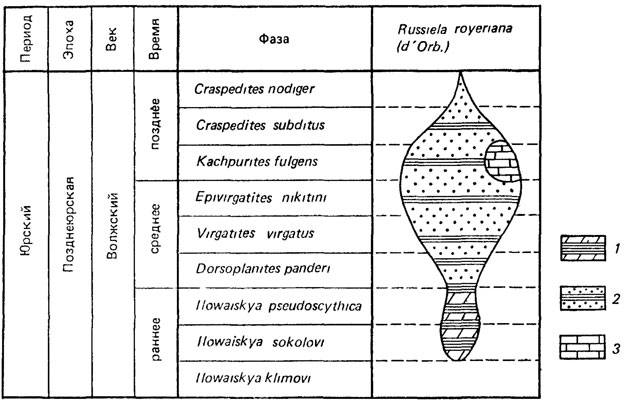
Рис. 124. Схема структуры полиморфного вида брахиопод Russiella-royeriana (d'Orb.). Волжский ярус Русской платформы. По В. П. Макридину с изменениями. 1 - Russiella roveriana undorae (Makrid.). Географический ареал Ульяновско-Саратовская синеклиза и Общий Сырт. Фации иловых впадин и тонкозернистых терригенно-карбонатных осадков относительно углубленной области шельфа. 2 - Russiella royeriana royeriana (d'Orb.). Географический ареал Московская и Ульяновско-Саратовская синеклиза. Фации песчаных и песчано-глинистых осадков сублиторали. 3 - Russiella royeriana tenuis Makrid. Географический ареал - Прикаспийская впадина; фации терригенных и Хемогенных карбонатных местами глауконитовых осадков относительно углубленной области шельфа
Естественно, что важное значение при обосновании выделения подвидов следует придавать картированию их географических ареалов и изучению их во времени (см. 5.1). Хронологический подвид не следует выделять в самостоятельную категорию, как это делают некоторые исследователи [555, 900, 2154, 2376], называя ее "транзитом" или "мутацией".
Необходимо подчеркнуть, что палеонтологи нередко разновидности (вариететы), выделенные в старых работах, без достаточных оснований переводят в подвиды. Это создает трудности для понимания истинной структуры вида, так как очень часто в качестве вариететов выделялись морфы, формы, аберрации, а не подвиды. Что касается других внутривидовых категорий, то они, как правило, не имеют принципиальных отличий от тех, которые применяются для современных видов.
Изучение внутривидовой изменчивости и структуры вида вымерших организмов проводилось для различных систематических групп, но особенно детально для различных групп беспозвоночных. Так, индивидуальная, или внутрипопуляционная (внутриколониальная у колониальных организмов), изменчивость исследовалась у фораминифер [259, 316, 599], кораллов [100, 103, 104, 323, 916, 1047], мшанок [710], брахиопод [271, 272, 422,960, 1011], остракод [265], двустворчатых [37, 64, 65, 300, 304, 364, 393, 437, 732, 810, 950, 951, 1138, 1205], брюхоногих [435, 437, 455, 456, 717, 821, 1027, 1028, 1029, 2002] и головоногих [89, 889, 1122, 1686] моллюсков. Эти исследования показали, что размах индивидуальной изменчивости у разных популяций одного и того же вида в различных условиях и у разных видов может весьма отличаться. Было замечено, что наиболее изменчивы эврибионтные виды, что может быть обусловлено, по Г. В. Никольскому [748], тем, что наличие широкой амплитуды изменчивости обусловлено приспособлением к изменчивой среде, в которой сформировался и существует тот или другой эврибионтный вид. Это положение подтверждается высокой изменчивостью эвригалинных морских видов, которые обычно были широко распространены в условиях отклоняющейся солености в полуморских бассейнах или в опресненных участках морей с нормальной соленостью. Примерами этого могут служить четвертичные моллюски Черноморского бассейна [435, 732] и сарматские и раннемэотические моллюски Понто-Эвксинской области [437]. Помимо адаптивного характера широкой изменчивости причиной этого явления могло быть уменьшение давления естественного отбора в связи с увеличением размеров популяций, существовавших в полуморских водоемах немногочисленных видов. То же объяснение, по-видимому, применимо и к солоновато-водным видам, высокая изменчивость которых многократно подчеркивалась Л. Ш. Давиташвили [292, 294, 296, 300]. Многочисленные конкретные примеры изменчивости солоноватоводных видов моллюсков приведены также в работах Л. А. Невесской и Л. Б. Ильиной [435, 437, 732, 735].
Наличие таких изменчивых видов в ряде случаев приводило к выделению морфологических "видов", которые при внимательном исследовании их изменчивости оказались лишь вариантами индивидуальной изменчивости [65, 810, 950, 1027, 1028, 1029]. К индивидуальной изменчивости относится также изменчивость отдельных стадий онтогенеза, отмечающаяся значительно реже, чем изменчивость взрослых организмов [265, 810, 1686].
Что касается групповой, или межпопуляционной (межколониальной для колониальных организмов), изменчивости, то для ископаемых видов (впрочем, часто и для современных) в ряде случаев трудно разграничивать географическую и экологическую изменчивость, так как пространственно разделенные популяции всегда существуют в какой-то степени в других экологических условиях, а экологически различные популяции обычно разделены и в пространстве, хотя и находятся в пределах одного ареала.
Как уже указывалось, представление об экологической и географической изменчивости древних видов дает сравнение выборок из многих местонахождений строго одновременных отложений. При этом основной трудностью является установление синхронности разделенных в пространстве ископаемых популяций, но при детальных биостратиграфических работах трудность устранима.
Экологическая или географическая изменчивость популяций одного вида отмечалась для многих групп организмов: фораминифер [316, 486, 599, 1333], кораллов [100, 103, 320, 1047], мшанок [506], брахиопод [117, 272, 422, 604, 906, 1011, 1211, 1236], двустворчатых [64, 65, 300, 393, 437, 732, 734, 735, 807, 809, 810, 899, 950, 951, 1714, 2348], брюхоногих [300, 435, 437, 717, 1027, 1028, 1029] и головоногих [89] моллюсков, млекопитающих [212, 213] и др.
Изучение групповой изменчивости часто приводит к необходимости выделения внутривидовых категории - подвидов (при географической, эколого-географической или хронологической изменчивости), пли морф (при экологической изменчивости). Так, ряд видов двустворчатых и брюхоногих моллюсков Черноморского бассейна, проникших сюда из Средиземного моря в начале голоцена, представлен эндемичными подвидами, возникновение которых было обязано, по-видимому, не только и не столько относительной географической изоляции, сколько другим гидрологическим условиям и, в первую очередь, понижению солености [435, 732].
В качестве примеров различных морф можно привести морфы бивальвии Mytilaster lineatus (Gm. in L.) и гастропод Rissoa membranacea (Adams) и R. parva (Costa) (рис. 125) со скульптированной раковиной, приуроченных к открытому мелководью плейстоценового и современного Черного моря, и морфы этих же видов с гладкой раковиной, характерные для опресненных районов [435, 732]. Скульптированная и гладкая морфы выделены также для мэотических гастропод Pircnella disjuncta disjunctoidies (Sinz.), причем их развитие тоже было обусловлено различной соленостью [437].
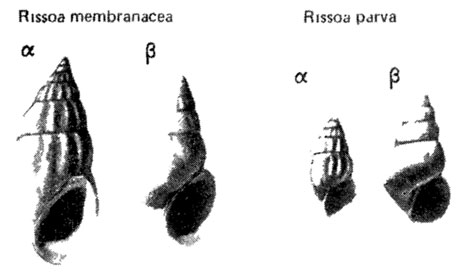
Рис. 125. Морфы Rtssoa parva (Costa) в голоценовом Черноморском бассейне. По Л. Б Ильиной. Морфа: α - скульптированная, β - гладкая
С жизнью на различных субстратах было связано возникновение двух морф бивальвии Ostrea edulis taurica Siem. в Черноморском бассейне: морфы с низкой связочной площадкой и умеренно толстостенной раковиной и морфы с высокой связочной площадкой и массивной раковиной. Первая из них связана с песчаноилистыми грунтами и характерна для устричников ("грядовая" морфа), а вторая - со скалистыми грунтами ("скалистая" морфа [732]). Аналогичные морфы описаны В. П. Макридиным [605] на примерах юрских брахиопод.
Во многих случаях, в том числе и в приведенных примерах, трудно различить подвид как географическую категорию и морф как экологическую категорию, так как приуроченность представителей того или иного подвида к разным участкам бассейна часто обусловлена различием грунтов, солености, температуры и других условий существования. Скорее, следует говорить не о географическом, а об "экологическом" подвиде. Изучению изменчивости популяций во времени с выделением в ряде случаев хронологических подвидов посвящены работы многих палеонтологов. Такого типа изменчивость также обусловлена главным образом влиянием смены экологических условий, т. е. может быть сведена к эколого-географической. Тем не менее ее следует рассматривать отдельно, так как прослеживание именно "хронологической" изменчивости дает возможность выявить пути видо- и формообразования. Примеры хронологической изменчивости, т. е. постепенного изменения признаков во времени, приведены в работах по фораминиферам [316, 599, 1333], кораллам [323, 1047], брахиоподам [885, 886], двустворчатым, [296, 300, 364, 393, 732], брюхоногим [300, 435, 436, 437, 821, 2002] и головоногим [722, 1400, 2384] моллюскам, пресмыкающимся [872] и др.
Изменчивость отдельных признаков во времени могла быть направленной или ненаправленной. Широкое распространение направленной изменчивости у моллюсков неогеновых бассейнов юга СССР отмечал Л. Ш. Давиташвили [296, 300], демонстрируя это на конкретных фактах. То же самое отмечалось Н. П. Парамоновой [807, 810] и Л. Д. Ильиной [437] на сарматских двустворчатых моллюсках, Л. А. Невесской [732, 734, 735] и Л. Д. Ильиной [435] - на четвертичных двустворчатых и брюхоногих моллюсках.
Рассмотрение амплитуды изменчивости (индивидуальной и групповой) у видов с различным эволюционным потенциалом показало, что наибольшая изменчивость отмечалась у видов, которые давали начало новым видам, Это было показано на видах одних и тех же родов двустворчатых моллюсков из неогеновых бассейнов юга СССР [734].
При рассмотрении конкретного случая симпатрического видообразования в голоценовом Черноморском бассейне оказалось, что наибольшая изменчивость предкового вида наблюдалась в период возникновения нового вида, а затем изменчивость всех признаков уменьшалась [734].
Исследование групповой изменчивости выявило ряд случаев параллельной изменчивости и гомеоморфии видов одного рода или близких родов, возникавших почти одновременно, но разобщенных пространственно, или же отмечавшихся в разные промежутки времени. Такие примеры проводились для кораллов [433], брахиопод [966, 1011], двустворчатых и брюхоногих моллюсков [40, 300, 437, 808, 810, 1029] и других групп.
Изменчивость вымерших форм можно изучать и методами фенетики [1232, 1233]. Впервые на необходимость при палеонтологических исследованиях изучать изменение отдельных дискретных признаков указали Г. Ф. Осборн [2047] и А. А. Борисяк [108]. Возможность применения методов фенетики в палеонтологии была отмечена Т. Г. Сарычевой и А. В. Яблоковым [914]. Впервые выделение фенов на ископаемом материале было проведено для ряда палеозойских брахиопод [272, 273], а также для сарматских гастропод [821]. Прослеживание отдельных фенов во времени и в пространстве показало, что некоторые из них, появляясь у немногих членов популяции, быстро исчезали, не повлияв на развитие той или другой группы, другие же - закреплялись в ходе исторического развития группы, приобретая таксономическое значение. Их изучение помогает прослеживать филогенетические связи и ход эволюции группы (рис. 126).
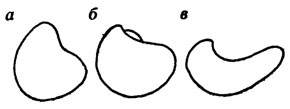
Рис. 126. Филогенетический ряд позднеюрских и раннемеловых Liostrea северо-сибирских морей. По В. А. Захарову. а - L. delta (Smith) из кимериджа; б - L. praeanabarensis (Zakh.) из волжского яруса; в - L. anabarensis Bod. из неокома
Изучение возрастных изменений у вымерших форм также имеет большое значение для решения проблем систематики, филогении и путей эволюции. Особенно детально эти изменения могут быть исследованы у тех групп, у представителей которых они прослеживаются на одном экземпляре, например, у археоциат [180], кораллов [322, 323, 439], головоногих моллюсков [89, 91, 348, 405, 887, 888, 889, 1160, 1686]. Однако и при отсутствии этих условий возрастные изменения, которые могут быть очень значительны, должны по возможности быть выяснены во избежание таксономических ошибок и неправильной интерпретации эволюционного развития. Изучение возрастных изменений в этих целях проводится на брахиоподах [117, 271, 960, 966], остракодах [265, 267], двустворчатых [38, 731, 732, 733, 734, 899, 951] и брюхоногих [16, 252] моллюсках и многих других группах, в том числе и позвоночных животных [871]. Во многих случаях изучение возрастных изменений помогает решить вопросы о возможных предках, путях формо- и видообразования и направлении эволюции. На ряде примеров показано, что новые признаки, возникновение которых отражало образование нового вида, появились на разных стадиях онтогенеза, т. е. развитие шло путем анаболии (геронтоморфоза), или же педоморфоза. Примеры анаболии с последующим ускорением онтогенеза известны для аммоноидей Medlicottia и Eumedlicottia [887]. Явления педоморфоза через девиацию также отмечены для аммоноидей Macrocephalus - Kepplerites - Cosmoceras, Orthosphinctes - Prorasenia - Rasenia [887, 1012]. Педоморфоз через фетализацию ("неотению") выявлен для ряда групп [25, 267, 734, 1012, 1013].
В отдельных работах приводились примеры полового диморфизма, отмечаемого для разных групп: остракод [265, 417], брахиопод [272], брюхоногих [315] и головоногих моллюсков [1915]. Выявление таких различий, связанных с полом, необходимо для выявления правильного систематического положения особей разного пола.
Следует сказать, что какие бы задачи ни ставил перед собой палеонтолог - исследование филогении, уточнение системы, выявление палеоэкологических особенностей той или другой группы организмов, проведение палеобиогеографического районирования, построение детальных стратиграфических схем - он сталкивается с необходимостью правильного определения объема вида и выяснения его изменчивости, что возможно только в рамках политипической концепции вида.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'