
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
4.5. Системные исследования в таксономии и филогенетике (В. М. Эпштейн)
Одна из важнейших проблем таксономии и филогенетики - проблема метода. В последние десятилетия эта проблема приобрела особую остроту в связи с развитием новых областей таксономии - числовой таксономии, геносистематики, кариосистематики, а также с дискуссиями, сопровождавшими обсуждение методов и результатов. Однако проблема метода в таксономии и филогенетике до сих пор остается неясной. Обычно, определяя методы этих наук, исходят из источника сведений для принятия таксономического и филогенетического решений. Такая трактовка метода рождена геккелевской триадой, рассматривавшейся в качестве метода филогенетики. Однако в литературе по мере вовлечения в сферу таксономии и филогенетики материалов из новых отраслей биологии многократно высказывалось мнение о недостатках такого "предметного" подхода, и его недостаточность становилась все более очевидной. Сейчас число групп признаков и соответственно методов их изучения очень велико. Так, Э. Майр [595] выделил пять типов таксономических признаков: морфологические, физиологические, экологические, этологические и географические, которые делятся на восемнадцать более мелких групп. Л. П. Татаринов [1033] разделил методы филогенетики на три группы: генотипические, фенотипические и экстрасоматические, которые делятся на четырнадцать более мелких групп.
Так как своеобразие исходного материала несомненно влияет на выводы о месте организма в системе и его филогении, эта классификация имеет определенную ценность. Однако на основании одних и тех же данных различные исследователи приходят к разным выводам. Отсюда следует, что характер материала не предопределяет решение. Недаром Д. М. Федотов [1101] отметил, что даже использование методов всего комплекса биологических наук не гарантирует успех реконструкции филогенеза. Недостатки "предметного" подхода состоят в следующем. 1) в число методов таксономии и филогенетики включаются любые методы и методики всего комплекса биологических наук, 2) обе науки не имеют своих, специфических для них методов и 3) не рассматривается процесс принятия таксономических и филогенетических решений.
Исходя из этих соображений, целесообразно дополнить этот подход другим - "функциональным", при котором методы систематики и филогенетики характеризуются как методы принятия систематического (точнее, таксономического) и филогенетического решений. При функциональном подходе выявляются специфические особенности метода каждой из этих наук; возникает возможность выделения и классификации подчиненных методов, с помощью которых решаются отдельные таксономические и филогенетические задачи; устанавливаются связи между методами этих наук. Классификация методов таксономии и филогенетики и связи между этими методами [1220] представлены на рис. 122.
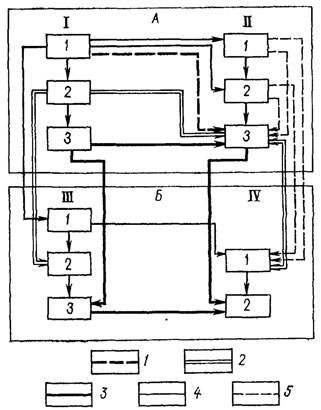
Рис. 122. Система методов таксономии и филогенетики. По В. М. Эпштеину. А - методы таксономии I - методы описания таксонов 1 - как совокупности отдельных систематических признаков, 2 - как целого, 3 - как системы; II - методы классификации: 1 - на основании уравновешивания систематических признаков, 2 - на основании априорной оценки одного признака или одной группы признаков, 3 - на основании апостериорной оценки признаков (филетического взвешивания). Б - методы филогенетики, III - методы реконструкции архетипов: 1 - как совокупности отдельных примитивных признаков, 2 - как целого, 3 - как системы; IV - методы реконструкции преобразований архетипов: 1 - реконструкция последовательного ряда архетипов; 2 - моделирование процесса преобразования архетипов, 1 - контур, соответствующий систематике XVII-XVIII вв. (описание по совокупности признаков и классифицирование); 2 - контур, соответствующий систематике и филогенетике XIX и первой половины XX в. (целостное описание, классифицирование, реконструкция архетипов и отдельных стадий филогенеза); 3 - контур, соответствующий систематике и филогенетике второй половины XX в. (системное описание, классифицирование, реконструкция архетипов и Динамики филогенеза); 4 - прочие связи между методами; 5 - предполагаемые связи между методами
Методы таксономии. Можно выделить две группы подчиненных методов (рис. 122): методы описания таксонов (I) и методы их классифицирования (II). Каждая из этих групп включает три метода (1, 2, 3), связи между которыми рассматриваются ниже.
Методы описания таксонов (I) разделяются на подчиненные методы по уровню описания организации таксона как целого.
Первый метод (I 1) характеризует организацию таксона как совокупность некоторого числа признаков. Этот метод в истории науки был первым, с него начинается изучение новых и малоисследованных таксонов, его использует систематик, приступая к освоению нового для него таксона. Этот метод применял К. Линней, хотя он отчетливо осознавал важность описания организма как целого. К. Линнею принадлежит известное изречение, что род определяет признаки. Однако К. Линней мог описать и классифицировать обрушившееся на него многообразие форм жизни только этим, наиболее простым способом.
Данный метод обеспечивает исходные функции систематики - диагностическую и классификационную, а также прикладную функцию в тех областях науки и практики, где можно достаточно надежно отличать один вид от другого. Естественность таких классификаций часто оказывается незначительной.
Второй метод (I 2) характеризует организацию таксона как целое. Этот метод впервые использовал Ж. Кювье - первым изучавший различные группы животных по всем системам органов и стремившийся объединять сравнительную анатомию и классификацию. Этот метод позволяет более обстоятельно описывать организацию таксона как уникальное явление природы, т. е. реализировать интегративную функцию систематики. Известно, что использование новых признаков и выяснение связей между признаками закономерно ведет к улучшению классификации. Таким образом, описание организации таксона способствует повышению естественности существующих классификаций.
Третий метод (I 3) - моделирование организации таксона как системы означает большой шаг вперед в реализации интегративной функции систематики, так как позволяет описывать целостность организации таксонов количественными методами. Кроме того, он позволяет улучшить классификацию путем строгой формализации понятий и стандартизации наборов признаков, которые должны быть необходимыми и достаточными для описания таксона как системы. Истоки этого подхода в трудах И. И. Шмальгаузена [1193] и А. А. Ляпунова [593]. Системные аспекты таксономии и филогенетики выражает мерономия, понятия и принципы которой разработаны С. В. Мейеном и Ю. А. Шрейдером [653, 667] и их последователями. Системный подход к описанию разных таксонов реализовали Д. Рауп и Ф. Стенли [864]. Этот подход используется и в наших исследованиях [1215, 1216, 1221].
Связи между этими методами таковы: вначале организация рассматривается в качестве совокупности некоторого числа признаков (I 1), затем в качестве целого (I 2), а далее в качестве системы (I 3). Вместе с тем необходимо отметить, что каждый из этих методов имеет область применения.
Методы классифицирования (II) выделены по их отношению к взвешиванию признаков. Изложению их сущности и оценке посвящена большая литература [595]. В данной работе они разделены на три группы: придание всем признакам равных весов- уравновешивание признаков (II 1); априорная оценка признаков (II 2); апостериорная оценка - филетическое взвешивание признаков (II 3). Взаимоотношения между ними таковы: придание всем признакам равных весов и априорная оценка признаков в исходных вариантах исключают филетическое взвешивание. Однако при филетическом взвешивании можно использовать некоторые идеи и методики, применяющиеся при уравновешивании признаков и их априорной оценке (поэтому на рис. 122 стрелки от II 1 и II 2 к II 3 показаны пунктиром).
Методы филогенетики. Методы филогенетики можно разделить на две группы подчиненных методов: методы реконструкции архетипов (III) и методы реконструкции их преобразований (IV). Первая группа включает три метода (1, 2, 3), вторая - два (1, 2).
Методы реконструкции архетипов (III) разделяются по уровню реконструкции архетипа как целого. Первый метод (III 1) позволяет представить архетип как совокупность примитивных признаков. Этим методом обычно пользуются систематики, реконструируя облик предков-основателей соподчиненных таксонов. Таким образом, нельзя создать целостное представление об архетипе, который к тому же реконструируется как неспециализированное существо, не располагающее адаптациями к конкретным жизненным условиям.
Второй метод (III 2) позволяет представить архетип в качестве целостного существа, обладающего комплексом адаптации к условиям среды обитания. Его применению должно предшествовать глубокое изучение таксона, для которого воссоздается архетип. Классическим примером использования этого метода являются реконструкции предковых форм в трудах А. Н. Северцова [926] и др.
Третий метод (III 3) позволяет представить организацию архетипа в качестве системы. Он использован в трудах Д. Раупа [864], и также был применен нами для реконструкции формы тела предковых форм пиявок. Необходимо подчеркнуть, что в упомянутых исследованиях подлежала изучению только форма тела, а моделирование внутренних органов - дело будущего.
Связи между этими методами (от III 1 к III 2, а от него к III 3) идентичны связям между методами описания таксонов.
Первый метод реконструкции преобразований (IV 1) - выяснение направления преобразований и сущности отдельных изменений. Этим методом построены все филогенетические деревья. Его отличают следующие недостатки: 1) он позволяет для каждых двух таксонов реконструировать только одну предковую форму, а не веера форм, которые соответственно теории дивергенции и палеонтологическим данным для хорошо изученных таксонов реализуются в процессе филогенеза; 2) пользуясь этим методом, нельзя представить филогенез в динамике.
Второй метод (IV 2) - моделирование филогенетических преобразований - позволяет реконструировать веера форм и с помощью мультипликации представить на экране преобразования этих форм в динамике [1215].
Взаимоотношения этих методов таковы: необходимо знать последовательность преобразований (IV 1) прежде, чем приступать к моделированию этого процесса (IV 2).
Связь между группами методов таксономии и филогенетики. Выявлена следующая связь между методами описания таксонов и методами классифицирования (группы I и II). При уравновешивании признаков (II 1) и априорной оценке признаков (II 2) нет необходимости в описании организации таксона как целого; отказ числовых таксономистов от филетического взвешивания разрушает представление об организме как исторически сложившейся многоуровневой системе; геносистематики, придавая показателям, характеризующим ДНК, большее значение, чем остальной организации таксона (которую они квалифицируют как фенотип), также не нуждаются в таком описании. Поэтому на рис. 122 показано, что оба метода используют описание таксона как совокупности признаков (I 1). При филетическом взвешивании (II 3) могут быть использованы (хотя и с разной степенью эффективности для классифицирования) все три метода описания таксонов.
Связи между методами описания видов и методами реконструкции архетипов (группы I и III) можно представить следующим образом. Из рис. 122 видно, что каждому методу описания соответствует метод реконструкции. Например, основываясь на наиболее примитивном методе - описании организации таксона как совокупности отдельных признаков, можно реконструировать архетип как совокупность отдельных примитивных признаков, однако нельзя воссоздать его облик в качестве единого целого. Исходя из наиболее продвинутого описания таксона как системы, можно реконструировать организацию архетипа как систему. Воссоздавать архетип как совокупность примитивных признаков в данной ситуации нецелесообразно, так как при такой реконструкции будет утрачена ценная информация о связях между признаками, содержащаяся в системном описании таксона.
Между методами реконструкции архетипов и методами реконструкции динамики филогенетических преобразований (группы III и IV) выявлена такая связь: реконструкции организации архетипа как совокупности примитивных признаков (III 1) и как целого (III 2) позволяют реконструировать только общее направление филогенеза и сущность отдельных преобразований (IV 1). Содержащейся в реконструкциях архетипов информации в данном случае недостаточно для описания филогенеза как процесса. Моделирование организации архетипа (III 3) позволяет представить филогенез в динамике (IV 2). Разумеется, этот метод описания архетипа дает возможность осуществить и более простую операцию (IV 1), однако такая операция нецелесообразна. Поэтому на рис. 122 связь между ними не показана.
Связь между методами классифицирования и методами реконструкции преобразований (группы II и IV) можно представить следующим образом. Придание всем признакам равных весов и априорная оценка признаков могут дать некоторое представление о направлении филогенеза и отдельных филогенетических преобразованиях. Поэтому связь между этими методами (II 1 и II 2 с IV 1) показана пунктиром. Филетическое взвешивание (II 3) дает надежное основание для реконструкции направления филогенеза и отдельных преобразований (IV 1), а следовательно, для моделирования процесса филогенеза (IV 2). Исторически обусловленные связи между методами таксономии и филогенетики можно описать с помощью трех контуров, охватывающих основные блоки, указанные на рис. 122.
Первый контур: использование методов I 1 - описание организации таксона как совокупности признаков, II 3 - взвешивание признаков без обращения к истории таксонов, III 1 и IV 1 соответствует особенностям систематики XVI-XVIII столетий. На рис. 122 этот контур показан пунктиром.
Второй контур: использование методов I 2 - описание организации как целого, II 3 - филетическое взвешивание III 2 - описание организации архетипа как целого, IV 1 - описание стадий филогенеза соответствует традиционному подходу, истоки которого в трудах Ж. Кювье, Ч. Дарвина, Э. Геккеля. На рис. 122 этот контур обозначен тонкой линией.
Третий контур: использование методов I 3 - системное описание таксонов, II 3 - филетическое взвешивание, III 3 - описание архетипа как системы, IV 2 - моделирование динамики филогенеза соответствует новым идеям и методам в современном естествознании. На рис. 122 этот контур обозначен толстой линией.
Принципы и методы биологической кибернетики в таксономии и филогенетике. Из анализа проблемы метода в таксономии и филогенетике следует, что перспективность конкретных методов и методик в этих науках зависит от того, насколько полноценно они позволяют описать целостность организма.
Проблема целостности - одна из "вечных" биологических проблем. Эта проблема в связи с классификацией животных была поставлена Аристотелем. Она стояла "за кадром" в "Системе природы" К. Линнея. Характеристика организма как живой системы, обменивающейся со средой веществом и энергией, была разработана в трудах Ж. Кювье, который на огромном материале по ныне живущим и ископаемым животным показал целостность организма во взаимодействии его частей ("принцип корреляций") и во взаимодействии со средой ("принцип условий существования"). Теория Ч. Дарвина объяснила целостность организма как результат эволюции путем естественного отбора и послужила основой для дальнейшей разработки проблемы, А. Н. Северцов и И. Шмальгаузен исследовали ее основные аспекты и дали материалистическую трактовку в противовес многочисленным теориям целостности, рассматривавшим это качество с механических и идеалистических позиций.
В 40-50-х годах развитие теории систем и кибернетики создало фундамент для принципиально нового подхода к проблеме целостности. Через десять лет после выхода в свет "Кибернетики" Н. Винера появились первые статьи И. И. Шмальгаузена, в которых эволюция описывалась в терминах и понятиях кибернетики. Этот синтез явился закономерным итогом исследований И. И. Шмальгаузена по проблеме "Организм как целое в индивидуальном и историческом развитии" [76, 193]. В серии кибернетических работ И. И. Шмальгаузен рассматривал эволюцию на популяционном уровне, в книге "Регуляция формообразования в индивидуальном развитии" - на организменном.
Большое значение для биологии (включая палеобиологию) имеют труды А. А. Ляпунова, в частности, его работа "Об управляющих системах живой природы и общем понимании жизненных процессов" [593], значение которых еще не оценено во всей их глубине. Труды И. И. Шмальгаузена и А. А. Ляпунова служат фундаментом для конкретных, заранее планируемых кибернетических исследований в таксономии и филогенетике. Анализ обобщений в этих областях науки свидетельствует о следующем: 1) организация таксона может рассматриваться как система; 2) основные области их исследований соответствуют основным областям биокибернетики - теории организации систем, теории процессов управления, теории информационных процессов; 3) конкретные количественные исследования закономерно связаны со свойственным кибернетике методом - математическим моделированием [634, 677, 864, 1215, 1216, 1217]. Исходя из указанных соображений, была высказана мысль о том, что эти работы находятся у истоков нового направления в науке - филогенетической кибернетики [1215, 1216]. Ее предмет - филогенез. Метод - математическое моделирование. Главные области исследований: системное изучение организации таксонов, процессов управления филогенетическими преобразованиями в пределах организации как сложной динамической системы, а также информационных аспектов филогенеза - эволюции генетического кода и эволюции многообразия различных групп организмов.
Системное изучение организации таксонов. Системный подход к описанию организации различных таксонов является логическим продолжением предшествующих исследований: описания организации таксона как совокупности признаков и последующего описания его организации как целого. Этот подход становится возможным после того, как биолог создает модель своего объекта как целого, используя для этого понятия и термины своей области науки. Системный подход представляет собой формализацию этой модели. Он осуществляется с помощью системных понятий и терминов, позволяющих произвести количественное описание целостности.
Таким образом, открываются новое, ранее недоступные возможности изучения органических форм и их преобразований в филогенезе. Для перехода от традиционных методов описания к системным большое значение имеет идентификация биологических и системных понятий.
При моделировании организации таксонов возникают большие трудности; оно может быть осуществлено в результате сложных и трудоемких исследований. Первые опыты системного моделирования посвящены описанию формы тела животных [864, 1215, 1216, 1221], так как форма тела выражает объем, внутри которого происходят взаимосвязанные изменения органов. Форма тела большинства животных может быть представлена в виде комплекса простых геометрических фигур. Форма тела - важнейший эктосоматический признак. Основополагающие работы Д. Раупа по моделированию спирально завитых раковин беспозвоночных показали, что этот метод в равной степени применим к описанию ныне живущих и ископаемых животных.
Системное описание организации в пределах одного из ее уровней осуществляется сходно, причем в этой работе можно выделить несколько этапов, в основном соответствующих обычно выделяемым этапам математического моделирования [601], но имеющих некоторые специфические особенности.
На основании изучения различных видов современных и ископаемых животных ("реальная ситуация") формируются их целостные описания. На этой основе возникает необходимость использования системного подхода в их организации ("постановка задачи") и построения соответствующих моделей.
Построение и описание модели. Модель организации в упомянутых ранее работах строилась в виде зрительного образа, отражающего наиболее существенные особенности объекта. Обычно систематик выделяет в качестве систематических признаков лишь отдельные особенности организации. При системном подходе к организации используются все признаки, необходимые и достаточные для построения изображения. Поэтому прежде всего составляется комплекс необходимых для построения изображения параметров, а для сравнения получаемых изображений - комплекс безразмерных величин (индексов) - отношений параметров. Изображения представляют объект как целое, а безразмерные величины выступают в качестве систематических признаков. Их набор используется как стандарт для описания любого нового или недостаточно изученного члена таксона.
Системный анализ модели. После изучения определенного числа реальных объектов выясняются черты их организации, которые являются инвариантными, и те, которые варьируют у разных объектов. Уже в начале работы объект рассматривается как система, и в нем можно выделить наиболее важные части, рассматриваемые в качестве подсистем. Затем на основании описания объектов с помощью модели можно выделить основную совокупность инвариант, образующую структуру модели и совокупность варьирующих особенностей, образующую ее программу.
Структура выражает самые существенные свойства модели. Наиболее важным аналогом структуры в биологии является "план строения", хотя с понятием структуры ассоциируются и другие важнейшие понятия - такие, как гомология, коннексия и гомеостаз. Наиболее важные аспекты, охватываемые структурой в моделях рассматриваемого типа, - постоянство частей, постоянство их положения, постоянство связей между ними. Наиболее важные и заметные (ключевые) признаки, образующие структуру таксона, используются в качестве его диагноза.
Программа таксона представляет собой совокупность его свойств, варьирующих в пределах, допускаемых структурой. Программа таксонов соответствует понятию пространства систематических признаков. Соотношение между структурой и программой таксона выражает степень его пластичности.
Построение пространства логических возможностей ("прогноз"). Совокупность теоретически допустимых сочетаний систематических признаков соответствует пространству логических возможностей [382], которые в свою очередь отвечает пространству теоретически допустимых таксонов. Значение прогноза теоретически возможных форм организации для функционального анализа существующих форм, образующих пространство реализации, со всей ясностью показал Д. Рауп. Основанные на этом подходе широкие возможности функционального анализа представлены в работах И. С. Барскова (см. 1.3).
При моделировании организации на всех его этапах проверяется ее соответствие реальным объектам. Изложенные методы позволяют осуществлять традиционное классифицирование, используя методы теории распознавания образов, которые позволяют избавиться от субъективизма, обусловленного произвольным выбором систематических признаков и их произвольной оценкой. Кроме того, они дают возможность сравнивать классификации для одного и того же таксона с целью выбора самой системной из них. Последняя оказывается и наиболее естественной. На этой основе был построен машинный определитель для рыбьих пиявок Палеарктики и проверена классификация этих животных на ЭВМ. Использование ЭВМ дает возможность вводить при построении классификации большое число признаков и совершенствовать ранее созданные классификации, применяя новую информацию, в том числе сведения о кариотипах и ДНК классифицируемых объектов. Осуществляемый интуитивно или сознательно системный подход способствует созданию матричных классификаций, позволяющих предсказывать облик теоретически допустимых таксонов. Теория и практика этих исследований разработаны Б. В. Преображенским [842] на примере ископаемых и современных табулятоморфных кораллов.
Перспективы системного моделирования тесно связаны с возможностями, представляемыми ЭВМ с изображающими устройствами, которые использовались в упомянутых работах. Они создают реальные возможности описания филогенеза с помощью зрительных образов, реконструируемых на основе точных, количественных исследований. Необходимо отметить, что в отличие от традиционных методов системный подход позволяет реконструировать в виде направлений в пространстве логических возможностей "веера форм" для каждого архетипа. Получив серии промежуточных изображений между двумя формами в пространстве логических возможностей, одна из которых рассматривается как предковая, а другая как производная, можно с помощью мультипликаций представить данный этап филогенеза в его динамике [1215].
Процессы управления филогенетическими преобразованиями. Обратные связи между органами в филогенезе. Современные представления об адаптациях свидетельствуют, что внешняя среда ставит определенные "проблемы", которые организм должен "решать" и что эти "проблемы" решаются в процессе естественного отбора [569]. И. И. Шмальгаузен [1193] отметил, что важнейшим аспектом процесса эволюции наряду с адаптацией, прогрессивной дифференцировкой, повышением организации и специализацией является его регулируемость. В процессе филогенеза происходят сложные изменения частей организма, связанные регуляторными взаимодействиями. Качества этой системы управления на уровне организма как целого контролируются естественным отбором. Таким образом, направленность эволюции, взаимосвязь между частями организма, относительная независимость частей, выражающаяся в "мозаичной эволюции", и ряд смежных проблем могут интерпретироваться в качестве различных аспектов управления филогенетическими преобразованиями организма как сложной развивающейся системы. К их числу принадлежит проблема координации.
Классификация координации, разработанная А. Н. Северцовым [926] и И. И. Шмальгаузеном [1187], основана на анализе причин, обусловливающих эти связи. Осуществляя причинное изучение организации, важно выяснить, являются ли установленные связи топографическими, динамическими или биологическими. Однако для системного описания организации таксона и ее филогенетических преобразований необходима количественная характеристика координации. Поэтому целесообразно в дополнение к существующей классификации координации по причинам вести их классификацию на иных основаниях: по степени детерминированности - детерминированные и стохастические (вероятностные) координации; по направлению взаимодействия - положительные и отрицательные координации; по месту в системе управления - координации, выражающие прямые и обратные связи между органами [1217].
В литературе содержится, начиная с классических трудов Ж. Кювье, В. О. Ковалевского, А. Н. Северцева и И. И. Шмальгаузена, немало примеров перечисленных типов координации. Однако в общих схемах филогенетического взаимодействия органов и развития различных групп организмов до сих пор принимались во внимание преобразования, представляющие собой прямую последовательность событий. Однако естественно предположить, что в процессе филогенеза, регулируемого естественным отбором преобразования частей организма как сложной биологической системы должны основываться на принципе обратной связи.
Определяя положение вида в системе, систематики выясняют последовательность появления систематических признаков (чем выше ранг таксона, тем древнее характеризующие его признаки). Затем, реконструируя ряд архетипов, устанавливают последовательность преобразований, связь между которыми является аналогом прямой связи. Так как соподчиненные таксоны характеризуются разными признаками, следует сказать, что в филогенезе мероны, обеспечивающие адаптациогенез ("лидеры"), непрерывно менялись. Естественно предположить, что мероны, ранее игравшие эту роль, на последующих этапах эволюции изменялись соответственно новому "лидеру", и эти изменения выражают обратные связи между ними [1214, 1219].
Для выяснения теоретически допустимых обратных связей между органами в процессе филогенеза с целью их выявления и изучения в организации исследуемого таксона необходимо построить ее модель.
Такая модель была использована для описания филогенетических преобразований организации пиявок, в частности их центральной нервной системы в ее взаимосвязях с другими системами органов [1219]. В соответствии с этими взглядами и на основе палеонтологических данных рассмотрим некоторые важные филогенетические преобразования у головоногих моллюсков (теутид). Этот пример любезно предоставил нам В. Н. Шиманский.
В настоящее время известны две группы головоногих моллюсков. Первая, включающая теутид, септид, октопод и вампиротеутид, имеет внутреннюю раковину то более развитую, то очень сильно редуцированную или вовсе ее не имеет. Вторая, куда входит один-единственный род Nautilus, характеризуется хорошо развитой наружной раковиной. У первых мантийный комплекс мономерный, есть крупные хорошо развитые руки и высокоорганизованная нервная система и органы чувства. У наутилуса мантийный комплекс димерный, имеются многочисленные небольшие щупальца, нервная система представлена нервными тяжами и расположенными на них ганглиозными скоплениями. У внутреннераковинных гомологом ноги является воронка - особое образование, через которую животное с силой выталкивает воду и благодаря этому, подобно ракете, быстро движется. У примитивных наутилид развита нога в виде двух лопастей, способных свертываться и также образовывать воронку.
Хорошо известно значительное число групп вымерших головоногих, от которых, как правило, сохраняется только раковина, значительно реже другие части организма (отпечатки рук, радула, клювы, чернильные мешки).
Исходя из этих материалов, не так трудно представить себе архетипы современных теутоидей, рассматривая указанные особенности с точки зрения их плезиоморфности и апоморфности (табл. 8).

Таблица 8. История развития отряда Teuthida
Дадим к табл. 8 некоторые пояснения. Совершенно очевидно, что нервная система в виде тяжей - признак плезиоморфный. В равной степени это касается и ноги, которая, вероятно, напоминала ногу гастропод. Небольшие щупальца, во-первых, также более древний признак, чем мощные длинные, во-вторых, коррелятивно они связаны с наружной раковиной. Моллюск при опасности прятался в жилую камеру, при больших же руках это невозможно. Они останутся добычей нападающего. Появление мантии, облекающей раковину, во-первых, снизило ее защитное значение, во-вторых, сделало животное более приспособленным к скольжению в воде, более маневренным. Приобретают очень большое значение мощные руки и прекрасный двигательный аппарат. Усложнение этих элементов было возможно только при значительной концентрации нервной системы.
Информационные аспекты филогенеза. И. И. Шмальгаузен рассмотрел основные проблемы эволюции в свете теории информации. Он подверг анализу биологический код, применив к его характеристике такие понятия, как количество информации, емкость канала связи, помехи, помехоустойчивость, надежность. Значение этих идей и разработок И. И. Шмальгаузена многократно рассматривалось в литературе [32, 688 и др.]. Анализ сведений о генетическом коде на молекулярном и клеточном уровнях с этих позиций, по-видимому, открывает широкие перспективы новых исследований. Он придает необходимую полноту изучению филогенеза в качестве процесса сопряженной эволюции организации и биологического кода как программы, соответственно которой происходит ее развитие. Изучение филогенеза с этих позиций методом математического моделирования начато работами В. В. Меншуткина и Б. М. Медникова [634, 677]. Такой подход, отдавая должное исследованиям в области гено- и кариосистематики, одновременно снимает претензии некоторых исследователей на исключительную роль сведений о ДНК в таксономии и филогенетике.
В последние годы развитие теории информации привело к решению задач, стоящих весьма далеко от проблемы передачи сообщений. В результате сложилась прикладная теория информации, становление которой связано с использованием теоретико-информационных представлений как средства для решения задач в других областях науки [497]. Философское осмысливание этих представлений привело к определению информации как отраженного в сознании многообразия. Если систематику можно рассматривать как науку о многообразии живой природы [595], то филогенетику можно определить как науку о происхождении и развитии этого многообразия. Таким образом, связь между этими областями биологии и прикладной теорией информации представляется естественной.
О значении многообразия как фундаментального показателя организации различных таксонов и его значения в эволюции свидетельствуют некоторые важные закономерности филогенеза: 1) новые группы происходят от сравнительно неспециализированных форм (правило происхождения от неспециализированных предков - правило Э. Д. Копа); 2) специализированные группы имеют тенденцию и далее двигаться по пути специализации (правило прогрессивной специализации - правило Ш. Депере); 3) организм не может вернуться к строению, ранее осуществленному в ряду предков (правило необратимости эволюции Л. Долло)*. Отсюда следует, что только пластичная организация, способная предстать в различных вариантах в зависимости от условий окружающей среды, имеет широкие филогенетические перспективы. Поэтому совершенство организации живых существ целесообразно рассматривать как единство трех составляющих: степени сложности (морфофизиологическое совершенство - морфофизиологический прогресс, по А. Н. Северцову), степени приспособленности (биологическое совершенство - биологический прогресс, по А. Н. Северцову) и степени многообразия.
* (Названия и формулировки правил даны по А. В. Яблокову и А. Г. Юсуфову [1234].)
Для различных таксонов количественная оценка степени морфофизиологического и биологического совершенства может быть получена с помощью балльных оценок; показателем степени многообразия является число видов [1218]. А. Мюллер [1980] показал, что процесс филогенеза всех крупных таксонов сопровождался пульсациями многообразия, изображения которых могут быть сведены к нескольким основным типам. Обычно предполагают, что эти фигуры обусловлены давлением среды, определяющим их контуры. Обычно форма упомянутых фигур зависит и от способности предковых форм производить многообразие или, используя обычную биологическую терминологию, от их пластичности. О том, что эти способности действительно играют очень важную роль, свидетельствуют факты освоения обширных зон жизни различными группами организмов через миллионы лет после вымирания их предшественников [952]. По-видимому, в данном случае в процессе естественного отбора одна группа животных исчезает в то время, когда организация другой еще не приобрела комплекса приспособлений, необходимого для освоения освободившейся экологической зоны.
Внешние причины, обусловливающие положение группы организмов в природе, относительно ясны - это преобразования рельефа, трансгрессии моря, изменения климата, исчезновение пищевых ресурсов, конкуренция и т. д. Оценка внутренних потенций предковых форм до сих пор, представлялась лишь в самой общей форме, наибольшими потенциями обладают неспециализированные, экологически пластичные формы; их эволюция идет по пути возрастания специализации; эти изменения на уровне организации как целевого необратимы. И. И. Шмальгаузен описывает этот закономерный ход событий как переход от ароморфозов к алломорфозам, а от них - к теломорфозам. Однако для количественной характеристики филогенеза необходимо изучение различных таксонов методом математического моделирования из организации и ее преобразований. Для этого необходимо рассмотреть филогенез в виде серии архетипов. Каждый из них должен быть представлен в виде модели, в организации которой выделяетя структура и программа. Количество независимо изменяющихся особенностей организации, выступающих в таксономии в качестве систематических признаков, определяет объем пространства логических возможностей, т. е. способность архетипа производить многообразие или его экологическою и морфофизиологическую пластичность. Эти потенции можно оценить в битах, если рассчитать теоретически возможные варианты организации как итог равновероятных двоичных выборов. Следует отметить, что предлагаемый подход не чужд традиционной таксономии, так как практически вся классификация построена с использованием дихотомических ключей, в которых систематические признаки представлены в виде иерархии двоичных выборов.
Таким образом, системный подход к изучению организации различных групп организмов и их исторического развития в дополнение к возможностям традиционных методов позволяет.
1. Осуществить количественное описание организации таксона как целого.
2. Воссоздать теоретически возможные варианты организации.
3. Улучшить систему исследуемого таксона путем усовершенствования описаний, методов классифицирования и усиления функционального анализа организации.
4. Облегчить и усовершенствовать работу "традиционного" систематика, осуществляя обработку данных и получение изображений на ЭВМ с дисплеями.
5. Исследовать динамику филогенеза, используя построение пространств логических возможностей, получение изображений серий переходных форм (веера форм) и их мультипликацию.
Новые возможности, представляемые системным подходом к проблемам таксономии и филогенетики, по-видимому, могут быть использованы также при решении некоторых других теоретических и практических задач палеонтологии.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'