
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
5.3. Методы палеогеоботанических реконструкций (С. В. Мейен)
Многие принятые в ботанике понятия имеют эквиваленты в зоологии. Для обозначения этих парных понятий иногда употребляются терминоэлементы "фито" и "зоо" (фитохория и зоохория; фитогеография и зоогеография). Но некоторые понятия специфичны для ботаники, например понятие растительность, являющееся основным для геоботаники или фитоценологии. Полного эквивалента этим дисциплинам в зоологии тоже нет. Может быть, поэтому понятие растительность иногда с трудом воспринимается биологами, получившими преимущественно зоологическую подготовку. При этом обычно смешивают понятия флора и растительность.
Чтобы довольно полно представить животное население, территории, достаточно привести список таксонов, распределение в пространстве и частоту встречаемости каждого из них.
Дополнительно к этим сведениям указывать, какой облик имеют представители того или иного таксона, обычно нет необходимости. Самое большее, что требуется еще сообщить, не являются ли особи, скажем, необычно крупными или, наоборот, мелкими, угнетенными. Остальное ясно из диагнозов таксонов. У растений дело обстоит иначе. Один и тот же вид может быть представлен совершенно разными модификациями, и для характеристики растительного покрова нередко важнее знать модификацию, чем точную видовую принадлежность растения. Например, если ограничиться словами, что доминантом растительного покрова (главным эдификатором) является сибирский кедр (Pinus sibirica), то нельзя понять, о чем идет речь - о лесе из высокоствольных деревьев или низком кедровом стланике. Помимо P. sibirica стланиковые заросли образуют и многие другие растения, известные также в виде обычных по облику деревьев. Некоторые виды родов Ficas и Microsideros в лесу растут, как лианы, а на открытых местах - как кустарники [520]. В зависимости от условий происходит смена так называемой жизненной формы.
Прекрасный анализ этого понятия и современные представления о жизненных формах дала Т. И. Серебрякова [947], есть много сводных работ по жизненным формам водорослей и грибов [143, 600, 819 и др.]. Понятие жизненной формы иногда употребляется и в зоологии [679], хотя имеет там несколько иной смысл, обозначая морфологический тип неродственных форм, заселяющих определенные местообитания. Сходные по биотопам и жизненным формам сообщества животных называют изобиоценозами или параллельными сообществами. Разнообразие жизненных форм растений свойственно таксонам всех рангов. Нередко в разных родах, семействах, порядках можно видеть полный параллелизм жизненных форм. Поэтому по жизненным формам можно устанавливать рефрены. Это дает возможность одним таксонам подменять другие в сообществах без ярко выраженных изменений в общем облике растительного покрова. Одинаковые с виду луга могут быть составлены самыми разными таксонами. Травянистая растительность обычно образована покрытосеменными, но в хр. Восточный Танну-Ола и в ряде мест МНР эдификатором травяного покрова стали селягинелли [276, 1120].
Иногда подбор жизненных форм явно связан с климатическими факторами. Например, в горных областях множество форм принимает облик подушек; для Средиземноморья характерны конвергентные формы жестколистных ксероморфных форм вечнозеленых кустарников [1968], образующих специфический тип растительности - чаппараль; некоторые растения мангровых зарослей, даже относящихся к разным семействам, настолько сходны друг с другом по общему габитусу и характеру листвы, что часто без специальной подготовки их трудно отличить друг от друга, если они не цветут. В других случаях местный параллелизм жизненных форм не удается связать с конкретными экологическими факторами. Это явление иногда называют географическим параллелизмом [2479]. Например, в Новой Зеландии 50 видов, относящихся к 21 семейству, принадлежат к диварикатным кустарникам (длинные междоузлия, ветвление под прямым углом, мелколистность). Для юго-восточной части о. Мадагаскара характерны растения с высокой степенью ветвистости и растения с укороченными побегами [2378]. Связь географического параллелизма с экологическими факторами совершенно неясна. Его предполагалось объяснить неполовым переносом генетической информации, а именно вирусами [2479] и другими агентами [648]. Столь же загадочный параллелизм известен и у многих животных [74, 594, 648].
Итак, для представления о растительном покрове знание жизненных форм совершенно необходимо и не может быть заменено знанием таксономического состава растений, но с другой стороны, оно и не может его заменить. Таксономический же состав растений данной территории отвечает понятию флора. Не обязательно под флорой подразумеваются все растения всей территории. Можно ограничиться определенными крупными таксонами (флора покрытосеменных, флора мхов или бриофлора) или определенной частью ландшафта (водная флора, высокогорная флора).
Помимо набора жизненных форм, понятие растительности включает частоту встречаемости как таксонов, так и жизненных форм, типы образующихся сообществ и взаимосвязи между сообществами. Геоботаника (или фитоценология) изучает не только саму растительность, но и ее взаимоотношения с экологическими (климатическими, эдафическими) факторами, динамику растительного покрова. Историческая геоботаника (палеогеоботаника или палеофитоценология) изучает историю растительности. Самостоятельность палеогеоботаники относительна, поскольку при реконструкции растительности прошлого приходится постоянно обращаться к современному растительному покрову. Наоборот, многие особенности современной растительности не могут быть поняты без учета ее истории.
Обсуждая реконструкции растительности прошлого, можно оставить в стороне реконструкцию растительности, непосредственно предшествующей нынешней и реконструируемой без обращения к палеонтологическим данным, а судить о ней лишь путем экстраполяции в прошлое наблюдающихся сейчас тенденций или путем исторического истолкования пространственной структуры растительности. Например, о сукцессии судят путем сравнения смежных участков, но этот путь чреват серьезными просчетами. Нас интересует реконструкция растительности достаточно далекого прошлого, когда приходится обращаться к палеоботаническим документам. Как и во всех исторических реконструкциях [654], будем считать идеалом не абсолютно полное знание об интересующем нас ископаемом объекте, а такую степень знания, которая возможна для еще существующего объекта (см. 1.4).
Для реконструкции растительности прошлого надо восстановить в статике и динамике следующее: 1) жизненные формы растений, 2) участие в растительном покрове разных таксонов и разных жизненных форм, 3) прижизненные сообщества растений, 4) взаимоотношения сообществ.
Общий смысл понятия жизненной формы уже изложен. Перечислить комплекс признаков, по которым устанавливаются жизненные формы, трудно. Обычно указывают признаки, имеющие прямое или косвенное отношение к общему облику растения. Это - форма кроны, наличие или отсутствие главной оси, размер осей, их ориентировка по отношению к субстрату, характер ветвления, пространственное распределение вегетативных и генеративных частей, динамика нарастания осей, положение почек возобновления, характер облиствления и др. [217, 946, 247]. Эти признаки устанавливаются по дефинитивным стадиям или по типу онтогенеза (например, различить истинную дихотомию оси от ложной можно, лишь проследив онтогенез оси). Для разграничения жизненных форм используются и анатомические признаки, такие, как количество вторичной древесины в осевых частях. Нетрудно видеть, что все это - обычные таксономические признаки. Можно сказать, что все признаки форм одновременно являются и таксономическими признаками (тем более, что набор жизненных форм входит в характеристику таксона), но обратное утверждение будет неверным. Например, в число признаков жизненной формы не войдут многие микроморфологические особенности и те из макроморфологических (эпиморфологических) особенностей, изменчивость которых мало сказывается на общем облике растения как компонента растительности.
Трудности реконструкции жизненных форм ископаемых растений очень велики и связаны не только с фрагментарностью материала, но и с отсутствием такой типологии жизненных форм современных растений, которая была бы вполне пригодна для типологических реконструкций. В геоботанике до сих пор идет оживленная дискуссия, что следует считать травой и что деревом. Как показано Л. Е. Гатцук [217] на примере понятия травянистое растение, в классификации жизненных форм до сих пор сказывается то обстоятельство, что первоначально на представления о жизненных формах большое влияние оказало лучшее знакомство с растениями умеренной зоны, когда необычные жизненные формы тропических растений представлялись как исключения из правил. Хотя в геоботанике предложено множество классификаций жизненных форм (в классификации А Гумбольдта их было 19, а в классификации А Гризебаха - 54), жизненные формы выделялись по положению почек возобновления, по экологическим факторам и экологическим нишам, по продолжительности жизни и др. [520, 946, 947].
Палеоботаники обычно обходятся небольшим числом понятий, употребляемых без должного внимания к их содержанию и объему. Чаще всего, как правило, ограничиваются указанием, что данное растение было деревом, кустарником или имело травянистый облик, особо выделяются эпифиты и лианы, иногда говорят о пахикаульных или лептокаульных деревьях. Во многих случаях трудно понять, на каком основании ископаемому растению приписывается некоторая жизненная форма. Например, в "Основах палеонтологии" [788, с. 436] написано о плауновидном Sublepidodendron, известном лишь по обломкам осей и фрагментам коры: "Преимущественно небольшие деревья, частью, вероятно, кустарники". Есть ли смысл в понятии кустарник в приложении к плауновидным, неясно.
Невозможно отобрать такую из имеющихся классификаций жизненных форм, которая была бы наиболее пригодна для ископаемых растений. Вероятно, палеоботаника нуждается в собственной классификации жизненных форм в силу как специфики некоторых ископаемых растений, так и дефектности ископаемого материала. Рассмотрим некоторые показательные примеры Мы не будем останавливаться на методах реконструкции общего облика растений, о них см. 1.4. Нас будет интересовать соотношение таксонов и отвечающих им жизненных форм у вымерших растений. В целом, при обращении к палеоботаническому материалу получается, что в прошлом во многих таксонах были жизненные формы, несвойственные современным представителям. Классическими примерами можно считать обнаружение древовидных членистостебельных (каламостахиевых) и плауновидных (лепидокарповых) позднего палеозоя. Древовидный облик этих растений был установлен еще в прошлом веке. Хотя реконструкции были получены лишь для единичных представителей, сложилось представление, что древовидные формы характеризовали таксоны в целом (рис. 5). В действительности же и каламостахиевым, и лепидокарповым были свойственны существенно разные жизненные формы. Среди лепидокарповых есть вид Lepidodendron serratum, у которого стебли тонкие, стела расположена асимметрично, а листовые следы сосредоточены на одной стороне. По-видимому, это было не древовидное растение, а лиана. О травянистости каламостахиевых Ангариды можно заключить из того, что в захоронениях с облиственными побегами и стробилами никогда не встречаются крупные стволы, а фитолейма на стеблях тонкая, что трудно ожидать у растений с мощной вторичной древесиной. Среди кордаитантовых помимо высокоствольных деревьев, вероятно, были низкорослые формы с ходульными корнями, обитатели мангр [1464]. Не менее неожиданным было обнаружение лептокаульных форм среди цикадовых мезозоя [1494]. Эти формы в отличие от обычных пахикаульных (с толстыми неветвящимися стволами) цикадовых имели тонкие стройные стволы с кроной из повторно делящихся ветвей.
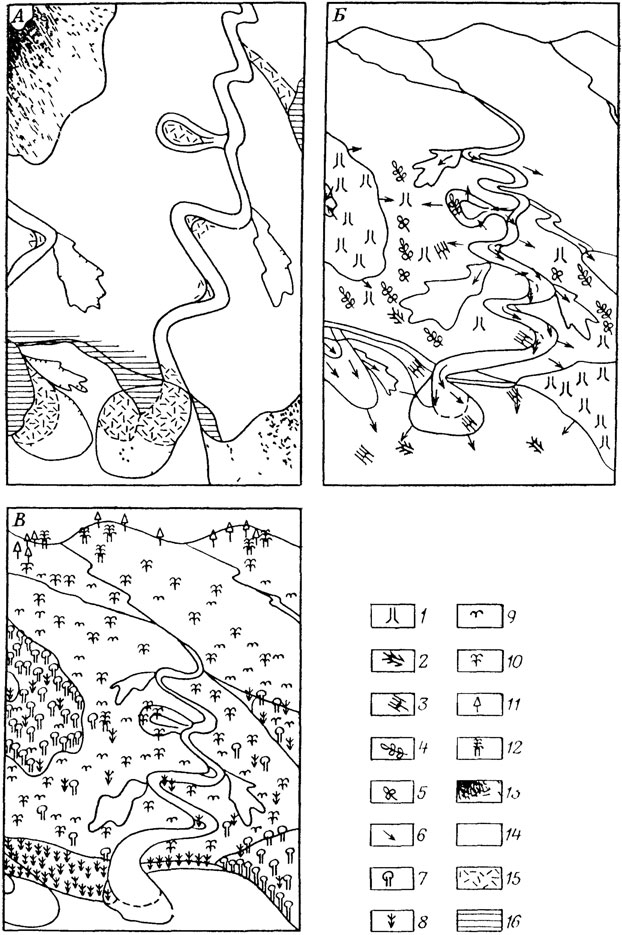
Рис. 5. Реконструирование растительных сообществ (на примере среднего карбона Западной Европы). По А. Скотту. А - реконструкция обстановок осадконакопления, Б - распределение ископаемых остатков растении, В - первичные сообщества растений, 1, 7 - плауновидные, 2 - облиственные побеги плауновидных, 3 - каламиты, 4 - птеридоспермы, 5, 9 - папоротники, 6 - возможные источники поступления остатков, 8 - каламиты, 10 - птеридоспермы, 11 - хвойные, 12 - кордаиты, 13 - 'болота', 14 - алевриты, 15 - пески, 16 - 'тростниковые' болота
О динамике роста растений можно судить по чередованию удлиненных и укороченных побегов [520, 636], по слоям прироста в древесине и вторичной коре, по сближению и разрежению листовых рубцов (характерно для карбоновых лепидофитов и других растений [1912, 1951]).
Для восстановления доли участия таксонов и свойственных им жизненных форм в растительности прошлого приходится обращаться к учету численности остатков и тафономическим наблюдениям, а затем опять же к косвенным аналогиям. После того как такой учет сделан, палеоботанику предстоит решить две проблемы, иногда неразрешимые: 1) в какой мере доля остатков соответствует доле прежнего покрытия площади растениями данного таксона, 2) как в тафоценозах представлены растения, относящиеся к разным участкам первичной растительности.
Особенно большую роль здесь играет сопоставление данных по макрофоссилиям с палинологическими материалами. Такое сопоставление позволяет в какой-то мере снять влияние "гипсометрического отбора" (см. 3.3). Взятые в отдельности макрофоссилии будут отражать лишь ту растительность, которая непосредственно примыкала к водоему, а также водные, полуводные и болотные сообщества. Нелистопадные травянистые растения, даже живущие на высокой пойме, если она не затопляется, как правило, не могут дать макрофоссилий в захоронениях. Взятые независимо от макрофоссилий палинологические данные неполноценны, так как систематическая принадлежность дисперсных миоспор часто неизвестна. В более древних отложениях для установления систематической принадлежности миоспор требуется контроль со стороны макрофоссилий. Особенно это касается палеозоя. В палинологической литературе, в частности отечественной, до сих пор широко практикуется отнесение палеозойских и мезозойских дисперсных миоспор к таксонам, установленным по макрофоссилиям, или даже к современным таксонам только на основании внешнего сходства с миоспорами известной систематической принадлежности.
Можно встретить указания на большой процент пыльцы гинкговых, цикадовых и беннеттитовых в пермских палинокомплексах Сибири. В макрофоссилиях из тех же слоев эти группы растений не представлены и поэтому считается, что названные выше голосеменные составляли растительность склонов и возвышенностей. В верхней перми Печорского бассейна отмечали большое количество пыльцы Ullmannia и Lebachia, хотя опять же побегов ни тех, ни других не находили несмотря на то, что комплексы макрофоссилий были богатыми и разнообразными. Этими хвойными "населяли" склоны и возвышенности.
Более детальное исследование ситуации и, в частности, изучение спор из спорангиев и микропиле семезачатков, сопоставление доминантов в комплексах миоспор и макрофоссилий [665, 1914] показали ошибочность всех этих заключений. Оказалось, что пыльца, сближавшаяся с гинкговыми, беннеттитовыми и цикадовыми, принадлежит растениям с листвой Rufloria и микростробилам Cladostrobus. Пыльца, прямо отнесенная к Ullmannia и Lebachia принадлежит (хотя бы частично) к птеридоспермам семейства Cardiolepiaceae. Противоречия между палинологическими комплексами и составом макрофоссилий исчезли. Их доминанты оказались одними и теми же, что позволяет считать представленные в них растения доминантами одной и той же растительности, но пока не ясно, какой, поскольку жизненные формы растений неизвестны. Аналогичные примеры по соотношению свидетельств миоспор и макрофоссилий можно привести и для мезозоя [154, 515].
Еще более трудной задачей является восстановление структуры сообществ, особенно палеозойских и мезозойских. Легче других можно представить себе монодоминантные сообщества маршевой, болотной или рипарийной (приуроченной к берегам водотоков) растительности (рис. 6). Кажутся правдоподобными реконструкции сообществ карбоновой болотной растительности, когда привлекаются данные одновременно по угольным почкам, по макрофоссилиям из подошвы и кровли угольного пласта и из пропластков пустой породы, по миоспорам из углей и вмещающих пород [2085, 2252]. Но и здесь можно говорить лишь об основных древесных эдификаторах, некоторых лианах и эпифитах, но не о сколько-нибудь достоверной синузиальной структуре. Например, до сих пор непонятно, какое место в структуре еврамерийской растительности карбона занимали многочисленные папоротники и сфенофиллы, большинство птеридоспермов. В литературе сложились канонические представления на этот счет, исходящие из реконструкций единичных форм, но основания этих представлений никогда как следует не анализировались. Часто эти представления не выдерживают первой же внимательной проверки. Например, долго считалось что птеридоспермы с листвой Sphenopteris hoeninghausii и стеблями Liginopteris были лазающими. Но известны местонахождения, где встречены только эти растения и не найдено никаких других, за которые они могли цепляться [1273]. Нередко палеоботаники опираются на характерные экологические признаки некоторых форм, подыскивают современный тип растительности, в котором распространены те же признаки, и используют полученную таким образом модель для реконструкции. Например, большое значение придается таким признакам растений, как резко выраженная гетерофиллия. По аналогии с водным лютиком или разными видами череды эти растения помещаются в воду, хотя гетерофиллия, причем очень резко выраженная, свойственна не только водным растениям, но и многим эпифитным папоротникам. Для отнесения растений, а с ними и всего сообщества к водно-болотному типу, часто используется наличие полостей и аэренхимных тяжей в стеблях и листьях. Однако у современного банана, плохо переносящего избыточное увлажнение, имеются полости в корнях и воздушные каналы в листьях [1285] Палеоботанику трудно, а то и нельзя учесть возможность широкой экологической амплитуды одного растения и далеко не прямую связь морфолого-анатомических признаков с определенными экологическими факторами. Есть растения вроде Phragmites communis L., который сейчас растет на пяти континентах - от Белого моря до дельты Ганга и болот в Уганде - и в зависимости от климатической зоны бывает летне-, зимне- и вечнозеленым. Никто не смог бы, основываясь на одних лишь морфолого-анатомических признаках, предсказать, что именно этот вид, а не какой-нибудь другой, будет столь безразличен к климатам и соседям по сообществу. Известны и другие виды с широкой экологической амплитудой, но весьма стабильными морфологическими, физиологическими, цитологическими и биохимическими свойствами [2206].

Рис. 6. Реконструкция растительности речной долины (на примере среднего карбона Западной Европы). По А. Скотту. На заднем плане заболоченный лес с доминирующими плауновидными на переднем плане - птеридоспермы у кромки воды - каламитовые
Более частные соображения, на которые опираются реконструкции отдельных черт растительности, приведенные в соответствующих разделах сводки В. А. Красилова [520] по палеоэкологии растений, здесь опущены В этой работе В. А. Красилова можно найти и соответствующие примеры. В целом следует сказать, что восстановить растительность с детальностью, позволяющей включить ее в современную классификацию, невозможно Чаще всего палеоботаник идет на это не потому, что ему удалось восстановить все черты, свойственные данному типу современной растительности, а лишь из-за того, что этот тип служит ему моделью.
Неслучайно поэтому в палеоботанической литературе почти нет сведений о "вымерших" типах растительности, хотя таковые вполне можно ожидать ввиду специфики как растений прошлого, так и комбинаций физико-географических условий. В качестве примера "вымершего" типа растительности можно привести данные из нижнего карбона Ангариды. Основными компонентами в комплексах макрофоссилий здесь являются лепидофиты, большей частью с неветвящимися стволами, и папоротниковидные (прапапоротники или птеридоспермы) типа Cardiopteridium - Angaropteridium. Часто в захоронениях встречаются только лепидофиты. Лепидофитовые же микроспоры преобладают в миоспоровых комплексах. Такие ассоциации проходят далеко на север, в те места, где, согласно палеомагнитным реконструкциям, должен был поблизости располагаться полюс. Здесь, в высоких широтах, должно было проявляться нынешнее чередование полярного дня и полярной ночи. Но особенность раннего карбона состояла в том, что световой режим сочетался с безморозным климатом. В Северо-Восточной Сибири в турнейском ярусе известны эвапоритовые породы, т. е можно предполагать аридный климат. Трудно представить себе лепидофитовую растительность этих мест - щетка из прямых, как палки, неветвящихся стволов разного размера по берегам водоемов, на несколько месяцев в году погружавшаяся в полярную ночь. Мы не знаем сейчас ничего похожего (рис. 7).

Рис. 7. Реконструкция раннекаменноугольного ландшафта Сибири
Не исключено, что в те эпохи, когда климат всей Земли был безморозным, существовали и иные "вымершие" типы растительности, особенно в высоких широтах. Кроме того, мы плохо представляем себе способность иных групп помимо покрытосеменных образовывать сложные синузиальные структуры. Существует мнение, что голосеменные не образуют, в отличие от покрытосеменных, сложных многоярусных сообществ [1042, с. 53]. Сейчас это действительно так, хотя среди голосеменных и в настоящее время есть разнообразные жизненные формы. Вполне вероятно, что в прошлом голосеменные демонстрировали гораздо более широкий спектр жизненных форм. В местах, где они доминировали, они вполне могли образовывать совместно с другими растениями не менее сложные сообщества, чем нынешние покрытосеменные [1764]. В. А. Красилов [520] считает, что по разнообразию жизненных форм покрытосеменные контрастируют лишь с хвойными. Не приходится забывать и о разнообразии жизненных форм у споровых растений.
Трудности реконструкции растительности прошлого и особенно возможность былого существования типов растительности, неизвестных в нынешних ландшафтах, заставляют палеоботаников прибегать к выделению очень обобщенных типов растительности, различающихся прежде всего по предполагаемой влажности биотопа. Таковы, например, типы растительности (гигрофильная и др.), выделяемые О. П. Фисуненко [1108], А. К. Щеголевым [1203], В. Гавленой [1712], М. В. Ошурковой [791] и другими палеоботаниками в пределах Еврамерийской области и Казахстанской провинции. Столь же обобщенными являются типы растительности, тафоценозы которых связаны (антракофильные) или не связаны (антракофобные) с угольными пластами. Эти типы первоначально были установлены для палеозоя [643], а затем и для мезозоя [520]. Остановимся теперь на значении реконструкций растительности прошлого. Они представляют собой самостоятельный интерес, так как вскрывают одну из существенных сторон эволюции органического мира. Без представлений о растительности, пусть очень гипотетических, невозможно осмысленно воспринимать органический мир континентов, обсуждать вопросы эволюции его в целом и отдельными компонентами. Даже гипотетические представления о характере растительности важны для обсуждения вопросов регионального и глобального литогенеза, реконструкции ландшафтов, обсуждения генезиса каустобиолитов и решения других вопросов. Как уже отмечалось (см. 5.2), реконструкции растительности неотделимы от флористических исследований, а следовательно, от фитогеографии и фитостратиграфии.
На значении палеогеоботанических наблюдений для стратиграфии остановимся отдельно. Первоначально для стратиграфии использовался главным образом списочный состав ископаемых растений. Правда, и тогда учитывалась частота встречаемости отдельных форм, т. е. геоботанический, а не флористический параметр (рис. 8). Еще в прошлом веке была замечена выдержанность некоторых ассоциаций в разных местонахождениях и их смена рассматривалась как смена типов растительности. Это служило основанием для выделения стратиграфических подразделений (например, в стефане Западной Европы). В течение первых 30 лет нашего века стал распространяться палинологический метод для разработки климатостратиграфии антропогена. Для более древних отложений климатическая интерпретация смены фитостратиграфических комплексов до сих пор используется довольно редко, хотя интерес к климатостратиграфии дочетвертичных отложений возрастает. Некоторые флористические смены, одновременно отражающие и коренные изменения растительности, возможно, служат показателем глобальных климатических перестроек. Так, на границе намюра А и В Западной Европы еще в прошлом веке было отмечено крупное изменение в составе комплексов растений. Примерно тогда же произошла радикальная смена комплексов в пределах Ангариды, а может быть, и Гондваны. Вполне вероятно, что эта палеоботаническая граница соответствует крупнейшему похолоданию, охватившему всю Землю и приведшему к исчезновению безморозного климата в высоких широтах [640]. Такой же глобальный характер, возможно, имело и похолодание, предполагающееся на границе мела и палеогена и отмеченное в перестройке растительных группировок в разных частях Земли.
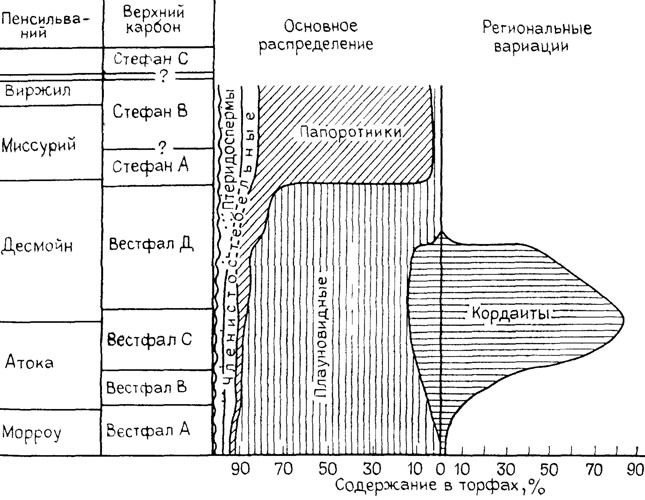
Рис. 8. Изменение процентного состава растительных остатков в окаменелом торфе на рубеже среднего и позднего карбона в Еврамерийской палеофлористической области. По Т. Л. Филипсу
Реконструкция последовательных смен растительности (палеосукцессий) позволяет выявить серию последовательных похолоданий и потеплений, нередко подтверждаемых изотопными палеотемпературными измерениями, и использовать эти данные для расчленения и корреляции толщ даже в тех случаях, когда только по списочному составу ископаемых растений этого сделать не удается. Примеры такого подхода имеются в работах В. А. Красилова [520, 522], детально рассмотревшего и теоретические предпосылки подобных климатостратиграфических построений. Можно надеяться, что именно на климатостратиграфической основе удастся снять противоречия между различными стратиграфическими методами, более точно сопоставить морские и континентальные отложения. Палеогеоботанические исследования при этом приобретут первостепенное практическое значение.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'