
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
5.2. Методы палеофлористических исследований и проблемы флорогенеза (С. В. Мейен)
Понятия "флора" и "растительность" четко различаются лишь в отношении современного растительного мира. Уже отмечалось [520, 642], что, говоря о флорах прошлого, палеоботаник в действительности подразумевает, скорее, геоботанические, чем флористические понятия. Приводится пример со стигмариевыми горизонтами (почвами), когда обнаружение таксона Stigmaria ficoides, если только он представлен в слое, важнее в геоботаническом (это показатель мангровой растительности), чем в собственно флористическом отношении. Некоторые таксоны ископаемых растений (изолированные стволы древесины, сердцевинные отливы каламитов и др.)" даже если им присвоены видовые эпитеты, отражают не столько виды растений как таковые, сколько жизненные формы, входящие в таксон неопределенного ранга.
Полное противопоставление понятий флора и растительность неправильно и по другой причине. Хотя, зная флористическую характеристику района, еще нельзя с уверенностью сказать о его растительности (как трудно сделать и обратное заключение), между составом флоры и характером растительности связи существуют и иногда очень тесные. Поэтому при характеристике растительности приходится давать флористический состав, а флористические сведения обычно даются по регионам, выделяемым с учетом геоботанических характеристик. И. Шмитхюзен [1198] вводит понятие дивизионы растительности для таких единиц растительного покрова, в которых одновременно учитываются флористические и геоботанические критерии. Он считает, что совпадают геоботанические и флористические единицы по крайней мере наиболее крупного ранга, поскольку те и другие в конечном счете контролируются климатом; см. также [520]. Однако дело не в ранге. Можно привести сколько угодно примеров как совпадения, так и несовпадения обоих типов границ любого ранга. Скорее, несовпадение можно связать с тем, что при районировании не всегда учитывалась возможность и целесообразность комплексирования геоботанических и флористических критериев, взвешивания одних с учетом других.
Приходится помнить, что без специальных и весьма трудоёмких исследований, как и без весьма гипотетических допущений (см. 5.3), палеоботаник не может опираться на геоботанические данные. Он чаще всего оперирует списками растений, привлекая в своих построениях в качестве непременного лишь такой геоботанический критерий, как частоту встречаемости отдельных форм. Геоботаническое насыщение палеоботаники - дело будущего. Но и в будущем палеофлористика (в узком смысле) сохранит самостоятельность как более объективная основа при реконструировании растительного покрова прошлого.
В науках, имеющих дело с историческими реконструкциями, нередко можно встретить разграничение объективных описательных процедур и субъективных интерпретаций, относящихся собственно к историческим реконструкциям. Особенно широко такое разграничение проводится в некоторых исследованиях по методологии геологии (в частности, в связи с обсуждением возможностей формализации и математизации исследований). Как в геологии, так и в других науках мы видим (в который уже раз) возрождение старого позитивистского мифа, разграничивающего объективные факты и субъективные интерпретации и неоднократно опровергавшегося историей науки. Тем не менее, по-видимому, можно говорить о некоей шкале, на одном конце которой будет располагаться абстрактная "абсолютная объективность" (т. е. сама природа в своей неисчерпаемости), а на другом - столь же абстрактная "абсолютная субъективность" (её даже невозможно представить), а в промежутке - рассеянные вперемежку протоколы наблюдений, эмпирические обобщения, гипотезы, теории и просто домыслы. Палеофлористические данные будут в своей массе ближе к объективному концу этой воображаемой шкалы, чем палеогеоботанические. Поэтому есть смысл прежде всего разобраться в источниках субъективности тех или иных палеофлористических данных (гипотетические моменты палеогеоботаники специально рассматриваются в 5.3). Главными источниками субъективности можно считать: 1) случайность выбора того или иного захоронения для детального изучения; 2) отбор самих растительных остатков в захоронении; 3) выбор техники для обогащения остатков (в случае мезо- и микрофоссилий); 4) неравномерное изучение собранных остатков, в частности, из-за того, что есть объекты, больше привлекающие всех исследователей, и объекты, на которые редко кто обращает внимание; 5) необходимость восстановления недостающих частей; 6) возможность по-разному взвешивать признаки и, следовательно, по-разному классифицировать объекты; 7) номенклатурные противоречия.
Остановимся подробнее на некоторых из этих пунктов. Два первых не нуждаются в пояснениях. Здесь мы часто следуем некоторым традициям, снова и снова возвращаемся к не раз изучавшимся местонахождениям и не обращаем внимания на другие, может быть, не менее важные. Традиции и привычка приводят к большему вниманию к одним группам растений, чем к другим, начиная со сборов их остатков на обнажения и кончая выбором для чтения уже опубликованных работ. Субъективен и выбор техники исследования. Часто очень трудно оценить эффективность того или иного метода. В течение многих лет палинологи применяют тяжелые жидкости для сепарации миоспор. Оказалось, что при обработке тяжелой жидкостью плохо сепарируются сильно скульптированные миоспоры [1369]. Можно изменить методы сепарации, но заранее невозможно сказать, какие дефекты будут у каждого из методов. Уже в том, что мы все же полагаемся на одни из этих методов в ущерб другим, проявляется субъективность.
Очень трудно, а иногда и просто невозможно, устранить субъективность, связанную с необходимостью реконструировать недостающие части объекта. Уже говорилось о трудностях при реконструкции жизненных форм и о субъективности выбора модели, служащей для типологической экстраполяции (см. 1.4). То же можно сказать о реконструкции всех недостающих частей. Распространенность самой процедуры реконструкции трудно осознать. Очевидно, что когда мы находим лист с оборванным краем и мысленно дорисовываем контур листа, то это - реконструкция. Менее очевидно, что когда мы находим отпечаток листа со всеми характерными признаками листа дуба и отмечаем этот экземпляр в каталоге как Quercus, мы тоже производим реконструкцию, домысливая (с помощью типологической экстраполяции) прижизненную ассоциацию этого листа со всем комплексом органов, свойственных роду Quercus (строение цветка, древесины и проч.).
Поэтому каждый раз, когда мы используем для районирования паратаксоны, установленные по разрозненным частям, вместо эвтаксонов (о различии паратаксонов и эвтаксонов, см. 4.1) мы вносим в палеофлористику субъективность. Более того, распределение паратаксонов в пространстве иногда отражает распределение не только эвтаксонов, но и отдельных их признаков. Хотя география признаков сама по себе интересна, ее нельзя отождествлять с палеофлористикой.
Наконец, очень важным источником субъективности в палеофлористике можно считать выбор ведущих систематических признаков, даже если остатки хорошо сохранились. Там, где один исследователь выделяет роды, другой ограничивается видами. Есть таксоны, удовлетворяющие всех палеоботаников и не подвергающиеся ревизии, и можно назвать множество таксонов, явно нуждающихся в ревизии. В этом смысле палеоботаническая систематика подобна любой другой.
Палеофлористические задачи можно условно разбить на стратиграфические и географические. Соответственно говорят о двух разделах палеофлористики - фитостратиграфии и палеофитогеографии. Эти два раздела тесно взаимодействуют и объединяются, когда речь идет об изучении становления и преемственности флор, т. е. флорогенезе. На общих принципах фитостратиграфии можно не останавливаться, поскольку они не отличаются от принципов, используемых в других разделах биостратиграфии.
Принципы палеофлористического районирования рассмотрены вместе с принципами палеобиогеографических исследований (см. 5.1). Поэтому в данном разделе ограничимся знакомством с тем, как общие принципы реализуются в палеофлористической практике, в частности при обосновании палеофитохорий, рассмотрим проблемы флорогенеза, а также остановимся на некоторых понятиях палеофлористики и на ее приложениях.
Целесообразно начать с рассмотрения группы понятий, относящихся в равной мере ко всем разделам палеофлористики. Для обозначения не самих палеофлористических единиц (фитохорий), а населявших их флор, по мнению некоторых палеоботаников, нужны какие-то понятия. Правда, можно отметить закономерность: подобные понятия время от времени появляются в статьях и докладах, но полностью отсутствуют в повседневном личном общении палеоботаников, в котором они обходятся просто понятием флора. Это, разумеется, недостаточный аргумент против введения понятий в теории. В литературе употреблялись понятия ценоквант [141], конкретная флора (Tolmatschew, 1971), тафофлора, стратофлора [910], геофлора [Chaney, 1959; цит. по 1145], полихронная флора [535] и др. Первые два понятия чисто ботанические, причем ценоквант - геоботаническое понятие, соответствующее наименьшему участку сообщества, а понятие конкретная флора - наименьшая флористическая единица. Тафофлорой считают "территориальную или стратиграфическую совокупность тафоценозов" [520]. Примерно такой же смысл имеет понятие стратофлора. Геофлора отвечает флоре крупной и устойчивой фитохорий (области, царству). Наконец, полихронная флора - хронологическое, а не географическое (хорологическое) понятие и отвечает наиболее крупным этапам развития растительного покрова.
Трудно сказать, какие из этих понятий действительно полезны, а какие появились как дань терминологическому пуризму. В палеофлористике, по-видимому, нужны понятия, в которых сочетаются собственно флористические и геоботанические критерии. Нужно какое-то понятие, обозначающее элементарную ассоциацию растений. В такой роли выступает тафоценоз. Известны группы сопряженных тафоценозов, повторяющиеся в разрезе, т. е. стратиграфически обратимые. Это палеофлористический аналог литологического цикла. Для него пока нет подходящего термина. Далее следуют комплексы тафоценозов, отвечающие в разрезе местным или региональным стратиграфическим единицам, а в хорологическом плане - фитохориям низкого ранга. Именно такой смысл имеют понятия тафофлора и стратофлора. Геофлора охватывает группы сопряженных тафофлор.
Не вполне ясна необходимость понятия еще более высокого ранга - полихронные флоры. Если исходить из определения, данного А. Н. Криштофовичем, который понимал под ними устойчивые великие флоры прошлого, охватывающие почти всю земную поверхность, то возникает вопрос - были ли на Земле такие флоры (более крупные, чем геофлоры). То, что мы сейчас знаем о флорогенезе палеозоя и мезозоя, указывает на длительное, преимущественно автохтонное развитие наиболее крупных геофлор, причем нет никаких данных, что иммигранты из соседних областей нивелировали региональную специфику геофлор. Флорогенез представляется как серия параллельно развивающихся стволов (геофлор), а полихронные флоры тогда надо рассматривать как объединение участков разных флорогенетических стволов по стратиграфической горизонтали. Смысл такого объединения непонятен. Правда, можно несколько видоизменить первоначальное представление о полихронных флорах и понимать под ними наиболее крупные этапы в развитии растительного покрова суши, отличающиеся степенью хорологической дифференциации. Тогда можно будет выделить всего четыре полихронные флоры: девонскую, каменно-угольно-пермскую (антракофитовую), триасовую и юрско-антропогеновую. Наконец, можно понимать под полихронными флорами лишь те этапы флорогенеза, когда дифференциация флор в пространстве минимальна (девон, средний - поздний триас). Думается, что эти преобразования понятия полихронная флора также не нужны как и само это понятие.
Наиболее детальные схемы районирования флор от карбона до неогена составлены для Евразии [796, 2063]. Схемы районирования для карбона, перми, юры и мела были составлены в середине 60-х годов и за прошедшие годы не потребовали серьезных изменений. Были уничтожены некоторые границы, закрыт ряд белых пятен, предложено несколько новых фитохорий (главным образом провинций), повышен ранг отдельных фитохорий, но в целом можно сказать, что схемы выдержали экзамен на стабильность, предъявленный большим новым фактическим материалом. Они оказались удобными для стратиграфических построений, не вступили в противоречия с палеоклиматическими и палеомагнитными реконструкциями, а также с результатами зоогеографического районирования. Поэтому накопленный опыт можно попытаться обобщить.
Главными критериями для выделения фитохорий служат различия флор по доминирующим таксонам. Ясной зависимости между рангом фитохорий и рангом доминирующих в них таксонов указать нельзя. Отчасти это связано с неустойчивостью самих таксонов у растений. Например, одни исследователи считают гинкговых и лептостробовых (чекановскиевых) порядками, а другие - классами. Цикадовые, беннеттитовые, гинкговые и лептостробовые слишком неуверенно разделяются на семейства, а среди палеозойских папоротников, плауновидных и членистостебельных весьма дискуссионно выделение и порядков. Поэтому при палеофлористическом районировании палеоботаники чаще всего оперируют родами и такими надродовыми группами, таксономический ранг которых не фиксируется. Говорят о доминировании кордаитов в верхнем палеозое Ангариды, глоссоптерид в одновозрастных толщах Гондваны, гинкговых и чекановскиевых в юре Сибири, а ранг этих групп не оговаривается. Систематическое положение некоторых групп, имеющих первостепенное фитогеографическое значение, иногда совершенно неясно с точностью даже до класса. Так, флора Катазиатского царства перми нередко называется гигантоптериевой по большой роли в ней гигантоптерид. Возможно, это голосеменные, родственные каллистофитовым и пельтаспермовым, но аргументировать это предположение можно лишь очень косвенными соображениями. В то же время принято считать, что гигантоптериды Катазии - единая крупная группа, весьма важная для палеофлористики перми.
При обосновании фитохорий важную роль играют и геоботанические критерии. Известно, что мангровая растительность свойственна не всем, а лишь тем фитохориям, которые расположены в низких широтах. Возможно, что турнейская флора Ангариды лишена эндемичных семейств и даже родов, но ее бедность по сравнению с еврамерийской и мелкорослость растений подтверждают самостоятельность Ангарской области.
При оконтуривании фитохорий приходится обращаться к общим экосистемным данным. Например, выделение в Индо-Европейской области поздней юры отдельных провинций [332] основано на немногих местонахождениях (Шотландская провинция - одним местонахождением). Тем не менее и само выделение провинций и проведение границ между ними подкрепляются картиной распределения ландшафтных зон, выделяемых по широкому комплексу палеогеографических признаков. Так, верхнеюрские местонахождения Франции, ФРГ и Польши приурочены к рифовым островам и берегам лагун аридного пояса, находящихся за рифами. Эти местонахождения входят в Южно-Европейскую провинцию. Шотландская провинция уже не относится к аридному поясу.
В отечественной литературе принята следующая иерархия фитохорий: царство, область, подобласть, провинция, округ. И. А. Добрускина [324] выделяет еще секторы, т. е. фитохорий с меридиональными границами в пределах широтных фитохорий (по аналогии с некоторыми схемами биогеографического районирования современной Земли). Если широтные фитохорий отражают прежде всего температурный и фотопериодический градиенты, то секторы отвечают таким факторам дифференциации растительного покрова, как климатические (влажность, континентальность), и флорогенетические. Последние иллюстрируются И. А. Добрускиной распределением пельтаспермовых в среднетриасовых флорах. Считая центром расселения пельтаспермовых районы, соседние с Уралом, она связывает картину их распространения в триасе с унаследованностью. Хотя секториальность широтных зон заметна в районировании разных эпох, специалисты обычно не выделяют секторов, а ограничиваются перечисленными категориями фитохорий. При этом иногда стираются меридиональные связи между фитохориями.
В литературе не раз обсуждалась номенклатура фитохорий [535, 796, 1041, 1043, 1846, 2063 и др.]. Обычно фитохорий именуются по названиям современных (Сибирская область) или прежних территорий (Ангарское царство). Предлагалось вводить в названия фитохорий указание их возраста (Бореально-меловая область и т. п.). Однако это возможно лишь в тех случаях, когда время существования фитохорий точно совпадает с геохронологическими единицами. В противном случае названия станут чрезмерно громоздкими. Порой вовсе невозможно ввести указание возраста в название фитохорий. Например, Субангарская область, если ее выделять по макрофоссилиям растений, появляется неодновременно в пределах Евразии - в интервале от артинского до татарского веков. Время формирования Гондванского царства остается крайне дискуссионным, поскольку границу карбона и перми, а также границы отделов карбона и перми провести в гондванских отложениях не удается.
При обосновании фитохорий учитывается их предшествующая история и последующая судьба. Так, своеобразию турнейской флоры Сибири может быть и не стоило бы придавать существенного значения, если бы мы не знали, что с турнейского века начинается тот процесс обособления Ангарской области, который продолжался до конца палеозоя. Кроме того, опираясь на выявленную систему фитохорий, можно перейти к выявлению того, как формировались и расселялись флоры в планетарном масштабе, т. е. к реконструированию флорогенеза и его причин. Соответствующий раздел флористики можно назвать флорогенетикой.
Флорогенетические исследования включают наложение филогенезов конкретных таксонов на систему фитохорий, прослеживание эволюции сообществ, прохореза сообществ и таксонов. Соответственно требуется знание филогении хотя бы доминирующих таксонов, структуры сообществ (см. 4.4) и точная датировка региональных флор. Связь флорогенетики с филогенетикой и стратиграфией обратная, поскольку филогенетический анализ должен осуществляться на конкретном биогеографическом фоне (см. 5.1), а знание основных черт флорогенеза позволяет корректировать стратиграфические сопоставления. Например, известно, что еврамерийский вид Sphenophyllum oblongifolium (Germ, et Kaulf.) Unger появляется в Кузбассе в середине алыкаевского горизонта [1659, 2341]. Один вид в чуждой флоре не дает оснований для уверенного стратиграфического сопоставления тех уровней, на которых он появляется в разных фитохориях. Однако если учесть, что эволюция сфенофиллов происходила преимущественно в экваториальном поясе, а это уже флорогенетическое наблюдение, то едва ли можно предполагать появление Sph. oblongifolium в Ангарском царстве и его миграцию отсюда в Еврамерийскую область. Это значит, что уровень появления Sph. oblongifolium в Кузбассе не может быть ниже уровня появления этого вида в Еврамерийской области (основание стефана). Отсюда следует, что верхняя часть алыкаевского горизонта не может относиться к среднему карбону (высказывалось мнение, что вся алыкаевская флора средне-карбоновая).
До сих пор специальные флорогенетические исследования касались становления лишь современных фитохорий. При этом обращение к палеоботаническому материалу было спорадическим. Такова, например, известная сводка Е. В. Вульфа [199] по исторической географии растений. На анализе современных растений построено учение Н. И. Вавилова о центрах формообразования. Участники дискуссии о месте происхождения и путях расселения покрытосеменных стали систематически обращаться к палеоботаническому материалу лишь с конца 60-х годов.
Попытки резюмировать хотя бы основные черты флорогенеза в планетарном масштабе по уже накопленным материалам пока не предпринимались. В палеофлористических сводках делается упор на различия и сходства флор в синхронических срезах, а не на их преемственность. Общие флорогенетические представления сводятся к указаниям на происхождение фитохорий, относящихся к эпохам высокой флористической дифференциации, от материнских космополитных или очень широко распространенных флор, занимавших континенты в эпохи низкой дифференциации. Так, указывается, что отчетливая дифференциация флор карбона пришла на смену космополитной флоре позднего девона. Известно, что сложная картина фитохорий Ангариды в поздней перми развилась на месте более однородной раннепермской флоры. Эти данные указывают лишь на одну сторону флорогенеза, а именно: на территориальную преемственность фитохорий, на усиление или спад общей палеофлористической дифференциации. Однако они не указывают, за счет чего возникли те или иные фитохорий - за счет иммиграции растений, локального возникновения или вымирания групп растений или перераспределения доминант по площади фитохорий.
Во флорогенезе протекают все три процесса. Например, позднепермские провинции по периферии Ангарской области выделяются (помимо прочего) по присутствию группы Callipteris - Compsopteris - Comia, скорее всего, пришедшей сюда из Субангарской области, т. е. с юга и запада. В перми пельтаспермовые шире всего распространены и наиболее разнообразны на северо-западе Субангарской области и в Печорской провинции, т. е. на Русской платформе и в Приуралье. Как считает И. А. Добрускина [324], именно это и определило различие средне-позднетриасовых флор западной и восточной частей Евразии. В позднем мелу Тихоокеанская провинция выделяется более длительным выживанием нильссоний и некоторых других растений. Перераспределение доминант под влиянием чередующихся похолоданий и потеплений можно видеть на севере Евразии в течение антропогена (по палинологическим и палеокарпологическим данным).
В приведенных примерах, количество которых легко умножить, флорогенетические события рассматриваются без учета филогенетических реконструкций. Прослеживается расселение уже сформировавшихся таксонов или выживание таксонов, ранее бывших убиквистами, в отдельных районах. При этом не освещается главный вопрос флорогенеза - место происхождения растений, заселяющих фитохории. Таких данных флорогенетика собрала немного прежде всего из-за того, что филогенетическое древо высших растений реконструировано еще очень плохо, а попытки совместить филогенетическое древо с системой фитохории можно пересчитать по пальцам. Результаты некоторых таких попыток поучительны.
Интенсивные палинологические исследования [1277, 1521, 1736, 1762, 1763 и др.] позволили проследить последовательную смену типов пыльцы раннемеловых покрытосеменных. Монокольпатная пыльца вверх по разрезу сменяется трехбороздной, трехборозднопоровой и, наконец, трехпоровой. Параллельно растет общее разнообразие пыльцы. То, что древнейшая пыльца принадлежит одному морфологическому типу (род Clavatipollenites), рассматривается как свидетельство монофилии покрытосеменных. Эта пыльца раньше всего появляется и быстрее всего достигает заметного процента в палинологических комплексах стран, лежащих по обе стороны от современной Атлантики, - в широтной полосе от севера Аргентины до севера США и от Габона до Англии.
В других местах Земли пыльца покрытосеменных появляется позже. В альбе род Clavatipollenites и близкие к нему роды встречаются почти повсеместно. Таким образом, перечисленные приатлантические районы были областью возникновения покрытосеменных. При этом там, где их пыльца появляется раньше всего и в наибольшем количестве, в палинологических комплексах доминирует пыльца хвойных, в том числе хейролепидиевых, а литологические признаки вмещающих толщ указывают на аридные условия. По-видимому, близок к истине Г. Л. Стеббинс [2322], указывающий на наибольшую вероятность возникновения покрытосеменных в обстановке сезонного дефицита влаги, и неправы те, кто считал местом возникновения покрытосеменных Юго-Восточную Азию и примыкающие к ней архипелаги, где сейчас наблюдается наибольшая концентрация архаичных покрытосеменных, но где пыльца покрытосеменных появляется в разрезе нижнего мела лишь с альба.
Этот пример поучителен в двух отношениях. Во-первых, без прослеживания палеонтологически документированной филогении можно легко принять рефугиумы и вторичные центры формообразования за место происхождения таксона. Во-вторых, важно привлекать для выявления истории групп растений палинологические материалы. Если бы палеоботаники опирались лишь на макрофоссилии, то они бы не включили в район возникновения покрытосеменных Габон, Бразилию и другие места, где макрофоссилии раннемеловых покрытосеменных пока не находили. Более того, первые макрофоссилии покрытосеменных появляются на ярус выше, чем пыльца.
Необходимость привлечения к проблемам флорогенеза палинологических данных выдвигает еще одну сложную проблему: какие споры или пыльцу продуцировали растения, которые определяли облик и специфику фитохории. Решение этой задачи усложняется тем, что с течением времени тип спор и пыльцы, продуцировавшийся растениями определенного таксона, мог сильно меняться. Например, в позднем триасе голосеменные семейства Peltaspermaceae продуцировали монокольпатную безмешковую гладкую пыльцу, т. е. пыльцу того же типа, что и пыльца гинкговых, цикадовых и беннетитовых. В поздней перми растения того же семейства продуцировали двумешковую (протосаккатную) ребристую и безмешковую ребристую пыльцу, которую пока невозможно отделить от пыльцы арбериевых (глоссоптерид). Поскольку пределы полиморфизма спор и пыльцы в таксоне заранее неизвестны, а параллелизм в многообразии спор и пыльцы между таксонами весьма обычен, мы всегда рискуем, интерпретируя палинологические данные, принять одни растения за другие.
Таким образом, флорогенетические исследования оказываются несравненно более трудоемкими и требуют гораздо более детальных палеоботанических исследований, чем просто палеофлористическое районирование. Правда, некоторые общие флорогенетические вопросы можно решать косвенными, менее трудоемкими методами. Так, в задачу флорогенетики входит выявление не только центров формообразования отдельных таксонов, но и мест наиболее интенсивного образования новых таксонов вообще. Эти места можно выявлять и не зная конкретных филогенезов путем анализа суммарного разнообразия растений разных фитохории во времени.
При изучении позднепалеозойских флор Сибири было замечено, что некоторые растения гораздо дольше, чем в экваториальной (еврамерийской) флоре, сохраняют архаичный облик. Лепидофиты до среднего карбона включительно сохраняют черты, свойственные экваториальным лепидофитам в позднем девоне - турне. Членистостебельные удерживают свойственную раннекарбоновым формам организацию фруктификаций до поздней перми. Значительная часть пермских папоротников соответствует по уровню продвинутости среднекарбоновым формам. Если для каждой группы, присутствующей в обеих флорах, выделить ведущие таксономические признаки и проследить на геохронологической шкале распространение не самих групп, а только общих признаков, то получается, что ни один из них не появляется у сибирских растений раньше, чем у еврамерийских; многие появляются одновременно или они сначала отмечаются у еврамерийских растений.
Около половины признаков удерживается у сибирских растений намного дольше, чем у еврамерийских. Очень многие признаки (синангии у папоротников, интегументоподобные спорофиллы и галониальные рубцы у лепидофитов, мощная вторичная древесина у членистостебельных, сетчатое жилкование у папоротников и птеридоспермов, крупная монолетная пыльца у птеридоспермов и др.) известны лишь у еврамерийских растений, а признаки, известные лишь у ангарских растений (дорсальные желобки, кистевидные полиспермы и эндоретикулоидная экзина у руфлориевых, редукция числа спор до 32 у Geperapteris), немногочисленны и устанавливаются с трудом. Показательно и то, что общее разнообразие сибирской флоры по числу видов и родов раз в пять ниже, чем в еврамерийской.
Таким образом, в карбоне и перми бореальная флора Сибири продуцировала гораздо меньшее разнообразие растений, "новой продукцией" она обеспечивала лишь себя и ничего не дала еврамерийской флоре, хотя многие ангарские растения, видимо, имеют еврамерийское происхождение.
Аналогичного анализа по другим флорам пока не выполнялось. Отдельные наблюдения показывают, что подобное соотношение вкладов экваториальных и бореальных флор в суммарное разнообразие растений сохранилось в мезозое и кайнозое. Это видно по распространению родов цикадофитов в юрских флорах на диаграмме, составленной М. П. Долуденко для других целей [332]. Количество общих родов между Сибирской и Индо-Европейской областями довольно велико, но ни один род не появился в Сибири раньше, немногие роды появились в обеих областях одновременно и несколько родов жили в Сибири дольше.
Предварительный анализ разнообразия растений в фитохориях палеозоя и мезозоя показывает, что внеэкваториальные флоры (не относящиеся к тропикам и субтропикам) были слабыми генераторами принципиально новых признаков растений и соответственно новых таксонов высокого ранга (от порядка и выше). С другой стороны, формы, появившиеся в экваториальных флорах и затем замещенные там более продвинутыми растениями, порой длительно удерживались (персистировали) во внеэкваториальных флорах - явление внеэкваториального персистирования [1954].
Анализируя надвидовую эволюцию покрытосеменных, Г. Л. Стеббинс [2322] привел много доводов за то, что главные центры генерации таксонов высокого ранга у этих растений располагались по периферии влажных тропических лесов, в районах с сильными сезонными колебаниями в количестве осадков. Данные о происхождении и начальном расселении покрытосеменных в раннем мелу хорошо согласуются с гипотезой Стеббинса. Пока трудно сказать, насколько приложима она к другим эпохам и другим группам растений. Есть данные, указывающие на то, что Субангарская область перми, расположенная в зоне аридного или семиаридного климата с высокими среднегодовыми температурами, была важным центром формообразования. К сожалению, территории, удовлетворяющие намеченным Г. Л. Стеббинсом критериям, хуже изучены в палеоботаническом отношении, чем области с более влажным климатом, способствовавшим угленакоплению. Поэтому одна из главных задач флорогенетики - выявление и детальное изучение флор аридных поясов, особенно их периферийных районов, обращенных к экватору.
Хотя флорогенетические исследования трудоемки и длительны, только с обращением к ним палеофлористика переходит от формального анализа ареалов и выявления отдельных совпадений в распределении растений и факторов среды к познанию закономерностей в эволюции растительного покрова, а это - необходимое условие методически более совершенного приложения палеоботаники для решения вопросов стратиграфии, литологии, тектоники и общей биологии.
По Евразии и другим материкам накопилось достаточно данных об изменении растительного покрова Земли от девона до антропогена. Можно проследить становление и исчезновение фитохорий, их расщепление и слияние. Общая схема флорогенеза, построенная для Евразии (рис. 3, 4), показывает максимум флористической дифференциации, приходящийся на пермь. Затем следует смягчение границ между фитохориями и сокращение числа фитохорий. Антропоген - новая кульминация в дифференциации флор.

Рис. 3. Географическое распределение главных филогенетических ветвей; на врезке показано распространение основных типов флор в позднем палеозое, времени главной дивергенции голосеменных. По С. В. Мейену. Области. 1 - нотальные (гондванские) внеэкваториальные; 2 - экваториальные и прилежащие к ним экотонные. 3 - бореальные, 4 - неопределенная фитогеографическая ситуация девона Clad - Cladoxylales, Calaraopit - Calamopityales, Callist - Calistophytales, D1 - Dicranophyllales; F - Fedekurtzia и близкие к ней растения, G - Gnetales; Ib - Ibykales (Iridopteridales); Ms - Marsiliales; Sl - Salviniales; - W - Welwitschales
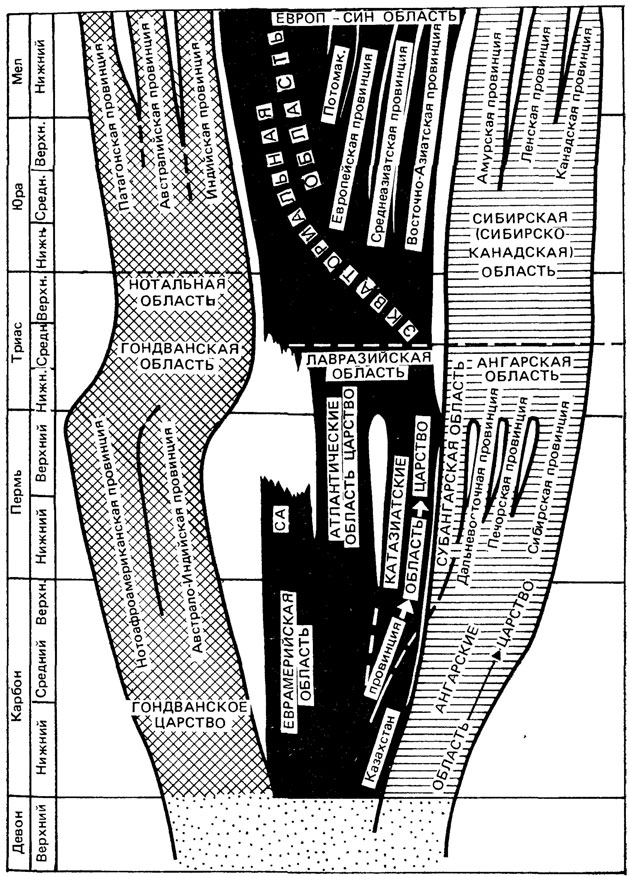
Рис. 4. Эволюция фитохорий в позднем палеозое и мезозое. По С. В. Мейену. СА - Северо-Американская область, Потомак - Потомакская провинция; Казахст. - Казахстанская провинция, Европ.-Син - Европейско-Синийская область
В целом ясно, что основными факторами, контролирующими флористическую дифференциацию, были климат, рельеф суши (особенно такие мощные складчатые пояса, как Урало-Монгольский в позднем палеозое или Альпийский, или Тихоокеанский в кайнозое) и крупные акватории, из которых в позднем палеозое и триасе главную роль играл Тетис, а начиная с середины мела - Атлантический океан. Пока невозможно дать интерпретацию каждой границы с указанием факторов, обусловивших ее появление и резкость. Границы высшего ранга, оконтуривавшие области безморозного климата с севера и юга, контролировались прежде всего астрономическим фактором - режимом инсоляции [641, 656]. Местные палеогеографические факторы оказывали на нее второстепенное влияние. Крупные и устойчивые перемещения границ такого ранга на зоне современной градусной сетки можно интерпретировать, скорее, как перемещение материков, чем самих границ, по поверхности геоида.
Карты палеофлористического районирования служат важным подспорьем палеогеографических, палеоклиматических и палеотектонических исследований, связь которых с палеофлористическим районированием обратная. При районировании учитывается общая палеогеографическая и палеоклиматическая ситуация, реконструированная по непалеоботаническим данным (см. ранее приведенный пример с выделением позднеюрских провинций). Хотя палеоботанические наблюдения сыграли немалую роль в появлении и обосновании идеи перемещения материков, сами палеоботаники используют мобилистские реконструкции, выполненные по широкому комплексу геолого-геофизических данных [749, 1417, 2130 и др.]. Более подробно о значении палеофлористических (как и других палеонтологических) данных для реконструкции палеоклиматов, ландшафтных обстановок, тектонических событий и стратиграфических исследований см. в 5.1, 5.3, 8.1 и 8.4.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'