
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
7.3. Эволюция как эпигенетический процесс (М. А. Шишкин)
В основе непротиворечивой эволюционной теории должен лежать общий исходный принцип, позволяющий объяснить все многообразие биологических закономерностей. Ни одна из них (в том числе и наследственность) не может быть противопоставлена остальным как нечто независимое от эволюционного процесса. Единственно приемлемой основой для такого общего объяснения представляется дарвиновская идея естественного отбора.
В понимании Ч. Дарвина субстрат естественного отбора составляют целостные организмы, конкурирующие в борьбе за существование и возможность оставить потомство. Результат отбора - возникновение все новых устойчивых и дискретных типов организации, которые регистрируются исследователями как ныне живущие или вымершие биологические виды. Именно высокая устойчивость этих типов, или адаптивных норм [1184], в пространстве и во времени создает возможность их сравнительного изучения и классифицирования и делает их следы, сохраняемые в геологической летописи, надежным средством для возрастных и биогеографических сопоставлений.
Говоря об этом свойстве адаптивных норм, мы в действительности подразумеваем, что их историческое существование поддерживается в поколениях непрерывной сменой особей, надежно реализующих в своем индивидуальном развитии данную видоспецифичную организацию. Каким образом создается эта устойчивость онтогенетического осуществления? На этот вопрос и должна ответить эволюционная теория. Объяснить эволюцию на основе естественного отбора - значит показать, что именно он ответствен за создание механизмов индивидуального развития, ведущих к детерминированному конечному результату (т. е. к норме). В классическом дарвинизме эта проблема не разработана, но необходимость ее решения очевидна. Поэтому любая концепция, претендующая на роль нового этапа в развитии дарвинизма, должна оцениваться прежде всего по тому, насколько она способна объяснить происхождение устойчивости нормального онтогенеза.
Первую попытку вскрыть соотношения между естественным отбором и индивидуальным развитием представляет теория зародышевой плазмы А. Вейсмана, в которой организм как объект отбора был заменен совокупностью наследственных зачатков (детерминантов), передаваемых в поколениях независимо от онтогенеза, через зародышевый путь. Развитие рассматривалось как пассивный результат действия этих зачатков, разделяющихся в ходе клеточных делений и однозначно детерминирующих по отдельности все части организма (что и объясняет его устойчивость). Причины возникновения этих предопределяющих свойств зачатков теория не объясняла. Эта попытка отделить развитие как область "соматогенных" (ненаследственных) изменений от процесса передачи половых клеток как сферы действия отбора оказалась в решительном противоречии с фактами. Во-первых, зародышевый путь во многих случаях не обособляется от соматического развития или же является его итогом (вегетативное размножение, чередование половых и бесполых поколений). Во-вторых, очевидно, что между структурой зародышевой клетки и исходом нормального развития нет однозначного соответствия. Удаление частей развивающегося зародыша вместе с их предполагаемыми детерминантами может не мешать регуляции развития к норме и, наоборот, одна и та же клетка способна дать начало разным устойчивым типам организации в случаях адаптивного полиморфизма. Необходимость обойти эти противоречия, сохраняя принцип мозаичной детерминации, заставила ввести в теорию дополнительные допущения: 1) об избыточности зародышевой плазмы по отношению к фактическому исходу развития; 2) о переносе части ее (резервной идиоплазмы) вместе с соматическими клетками в ходе развития; 3) об изменении ее состава с помощью отбора (борьбы) детерминантов под действием внешних факторов [159, 2471]. Таким образом, вопреки исходному пункту своей концепции А. Вейсману фактически пришлось признать, что именно индивидуальное развитие является источником изменений, вызываемых естественным отбором.
Эти трудности в понимании эволюционной роли онтогенеза, по существу, разрушившие теорию А. Вейсмана, перестали, однако, казаться актуальными на следующем этапе развития идеи дискретной наследственности. После переоткрытия менделевских законов, казалось бы, все стало ясно. Если все признаки независимо определяются генами, то онтогенез можно рассматривать как несущественную промежуточную инстанцию между теми и другими, не представляющую специального интереса. Чтобы понять историческую роль отбора, достаточно оценить его действие непосредственно на гены.
Оценка этой роли, характеризующая представления раннего менделизма, может быть суммирована в нескольких общеизвестных положениях: 1) наследственная изменчивость подчиняется исключительно правилам менделевских расщеплений; 2) новые варианты могут вноситься в нее лишь за счет мутаций (эта возможность, впрочем, также оспаривалась); 3) естественный отбор способен лишь отсеивать наследственные факторы, уменьшая разнообразие форм и, таким образом, не может выполнять той созидательной функции, которая приписывается ему дарвинизмом.
Принято считать, что эти упрощенные воззрения были преодолены в ходе развития моргановской хромосомной генетики и что ее объединение с дарвинизмом привело к созданию новой синтетической теории, реабилитировавшей идею отбора. Для оценки сущности этой теории необходимо прежде всего установить, какие принципы лежат в ее основе и как она объясняет главную эволюционную проблему - устойчивость нормального индивидуального развития.
Очевидно, что ее исходным пунктом, как и у А. Вейсмана, служит принцип отбора наследственных факторов (в данном случае - менделевских аллелей). Хотя уже с 40-х годов в изложениях теории одновременно подчеркивается, что объектами отбора являются фенотипы, тем не менее "ее идеалы остались прежними" [750]. Движущий отбор и сегодня определяется в ней как процесс замещения аллелей, а скорость эволюции - как скорость этого замещения [269]. В результате отбора ожидается увеличение однородности генетического состава популяций [999, 1164], а наличие в них высокой гетерогенности, несмотря на все попытки альтернативных объяснений (частотозависимый отбор и т. п.), приравнивается прежде всего к вмешательству факторов, действующих "несмотря на силы отбора" [7, с. 51]. Все это равносильно признанию, что подлинными единицами отбора предполагаются именно гены.
С этим принципом неразрывно связано принятие теорией и другого вейсмановского представления - об однозначных соотношениях гена и признака (хотя формально оно отрицается моргановской генетикой). Гены здесь обычно наделяются постоянными маркерами в виде коэффициентов приспособленности, сохраняемых ими во всех рекомбинациях. Кроме того, поскольку давление отбора оценивается только по изменению частоты форм в поколениях [1164], то ясно, что судить таким путем об изменениях генного состава можно лишь при наличии прямой связи между признаками и генами.
Следующий важный момент в оценке синтетической теории касается ее представлений о взаимоотношениях мутационного процесса и отбора, а также о причинах фенотипической устойчивости. Позиция теории по всем этим вопросам однозначно вытекает из отождествления ею менделевских факторов с хромосомными локусами*, в силу которого мутация рассматривается как акт создания элементарного менделевского различия. А поскольку такое различие должно контролировать пару альтернативных (аллельных) наследственных признаков, то мутация приравнивается к появлению нового устойчивого фенотипа. Хотя сторонники теории часто не замечают ее сальтационистской природы [594, 898], в действительности она очевидна и имеет ограничения лишь технического характера. Там, где две устойчивых морфы или расы различаются только по одному менделевскому фактору, это считается прямым указанием на происхождение одной формы из другой путем единичной мутации [1509, 269]; если же предполагается множество таких различий (например, у преемственных видов), то путь их приобретения представляется как цепь мутаций [1509, с. 52].
* (Хотя это отождествление и принято в моргановской генетике, оно не раз вызывало возражения [462, 848]. Т. Морган также признал его не без колебаний, допуская вначале, что менделеевские факторы суть лишь абстрактные свойства, каким-то образом связанные с локусами [708]. С развитием биохимических методов генетического анализа, где локус выступает как единица матричного синтеза, несоответствие указанных понятий вновь становится для генетиков все очевиднее [269, с. 290; 256].)
Это означает, что для синтетической теории вопрос об устойчивости нормального фенотипа и реализующего его онтогенеза вообще не является предметом рассмотрения; эффект всякого генетического изменения по определению считается ею наследственным, т. е. устойчивым. Хотя и признается влияние на него внешней и внутренней среды, сам факт его существования в виде особого маркера полагается очевидным и не зависящим от отбора (последний лишь оперирует такими наследственными признаками). Подлинной причиной появления нового признака считается мутация, а не отбор.
Следовательно, в основе синтетической теории лежит не столько реальный мир фактов хромосомной генетики, сколько редукционистские упрощения, унаследованные от А. Вейсмана и ранних менделистов (отбор генов, линейные связи генов и признаков, появление устойчивых новшеств путем мутаций). Такие важные эмпирические обобщения генетики, как устойчивость нормы ("дикого типа"), по сравнению с мутационными аномалиями и тенденция к их поглощению в скрещиваниях; нарушения менделевского наследования, выражаемые в понятиях экспрессивности и пенетрантности; гетерогенность однотипных природных аномалий; зависимость признаков от генома в целом; лабильность доминирования - не находят самостоятельного выражения на языке синтетической теории и низводятся ею на уровень несущественных помех, затемняющих правильную картину менделевских расщеплений.
Эти выводы вполне согласуются с сегодняшней тенденцией к переоценке синтетической теории в глазах генетиков. Ее все чаще характеризуют лишь как удобную абстракцию [465] или временное упрощение [999], а само определение эволюции как изменения генетического состава популяции признается редукционистским [597]. Иногда прямо указывают, что видеть здесь синтез генетики и дарвинизма - значит "выдавать желаемое за действительность" [39, с. 413]. К числу "вынужденных упрощений" теории относят сегодня наиболее существенные ее принципы: отбор аллелей, оценку их по вкладу в приспособленность, однозначное соответствие генотипа и фенотипа, возможность суждения о генном составе без учета закономерностей онтогенеза; используемые теорией модели действия отбора на частоты локуса с двумя аллелями характеризуются как "совершенно нереалистичные" [999].
Формальное признание фенотипа объектом отбора неизбежно заставляет признавать и эволюционную роль модификационной изменчивости [596], что на самом деле равносильно "пересмотру основных положений генетической концепции эволюции популяций" [353, с. 373] и означает введение в синтетическую теорию чужеродной пристройки ad hoc [487]. Характерно также нынешнее стремление теории отказаться от идеи генетического груза [269, 596, 669, 214], вызванное трудностями объяснения генетического полиморфизма, который обнаруживается внутри нормальных популяций. Но вместе с концепцией груза исчезает и неотъемлемое для синтетической теории объяснение движущего отбора как уменьшения генетической дисперсии популяции по приспособленности. Понятие отбора аллелей становится бессодержательным. Одним из косвенных и независимых признаний этого факта является предложение о замене генов хромосомными континуумами в качестве будущей основы популяционной генетики и эволюционных построений [1879].
Все эти попытки модернизировать синтетическую теорию объективно отражают невозможность совместить дарвиновское представление об отборе целых организмов (и их онтогенезов) с редукционистскими принципами ортодоксального менделизма и указывают на стремление ограничить роль последних. Но иногда возникшее противоречие пытаются решить и противоположным путем - за счет дарвинизма. Так, М. Д. Голубовский [255], почти повторяя слова В. Иогансена*, считает, что развитие учения о наследственности привело к последовательному ограничению постулатов селектогенеза. Место дарвиновской неопределенной изменчивости заняла строгая упорядоченность расщеплений, ограниченная менделевскими правилами и не зависящая от отбора. Поэтому созидательной роли отбора не существует - но сохраняет лишь то, что создается мутациями. А значит, нет и постепенности эволюции - ее суть составляют мутационные скачки, оцениваемые отбором. Этот призыв вернуться к представлениям Г. де Фриза и В. Иогансена неслучаен: он отражает ясное понимание несовместимости принципов, положенных в основу существующего "синтеза".
* ("Генетика вполне устранила основу дарвиновской теории подбора" (Johannsen [1915], цит. по [1104, с. 191]).)
Все это вновь заставляет нас обратиться к поискам теоретического обобщения, которое позволяло бы решить главную проблему дарвиновского учения об отборе - проблему возникновения устойчивости индивидуального развития. Одновременно оно давало бы возможность уяснить смысл эмпирических закономерностей, вскрываемых генетикой. Ибо явления наследственности, составляющие ее предмет, означают передачу признаков [310], т. е. не что иное, как их реализацию в онтогенезе потомков. Правильное наследование есть осуществление типичного (стабильного) развития [354, 670, 1194, 1196, 1684], тогда как нарушения наследования суть нарушения онтогенетической устойчивости. Поэтому наследственность и способность к устойчивому развитию составляют одну и ту же проблему [280, 922], и у нее может быть только одно решение, которое должно выражаться в понятиях, пригодных для описания онтогенеза.
Теория, удовлетворяющая в своей основе указанным требованиям, существует. Это - учение о стабилизирующем (канализирующем) отборе И. И. Шмальгаузена - К. Х. Уоддингтона, которое исходит из представления, что отбор по фенотипам ведет к созданию помехоустойчивого развития, реализующего эти фенотипы. Устойчивость признаков рассматривается здесь "не как свойство генов, а как выражение взаимозависимости частей в корреляционных системах развивающегося организма" [1196, с. 174]. Поскольку источником эволюционных изменений здесь признаются уклонения самого процесса развития, эта теория заслуживает названия эпигенетической. "Не изменения генотипа определяют эволюцию и ее направление. Наоборот, эволюция организма определяет изменение его генотипа" [1185, с. 57]. Несмотря на неизжитость в языке теории ряда чуждых или избыточных понятий, она в принципе позволяет раскрыть онтогенетический смысл фактов генетики.
Если стабильность итога развития представляет собой продукт эволюции, то же самое касается и наследственности. Это - одно из центральных положений эпигенетической теории. Создание наследственного признака описывается ею как накопление генетических изменений в пределах, допускающих осуществление отбираемого фенотипа [1186, 1196], или как генетическая ассимиляция последнего [2440, 2441]. В соответствии с этим сырой материал эволюции должны составлять неустойчиво наследуемые изменения, т. е. такие, которые могут быть охарактеризованы как дарвиновская неопределенная изменчивость. Согласно И. И. Шмальгаузену [1192], это лабильные неадаптивные морфозы (модификации) нормы, реализуемые на гетерогенной основе. Следовательно, созидательная роль отбора заключается в стабилизации первично неустойчивых типов онтогенетических реакций. Чтобы понять механизм этого процесса, необходимо сначала охарактеризовать главные свойства стабильного и нестабильного типов развития, найдя им адекватное выражение на языке генетики.
Прежде чем сделать это, суммируем сущность различий в понимании природы наследственности (устойчивости), разделяющих синтетическую и эпигенетическую теории. Для первой наследственность есть свойство особых элементарных носителей, не требующее причинного объяснения в рамках самой теории. Законы наследственности здесь действуют независимо от естественного отбора [255], т. е. эти два фактора рассматриваются как самостоятельные партнеры в осуществлении эволюционного процесса. Напротив, для эпигенетической теории наследственность есть выражение стабильности целостного индивидуального развития, создаваемой только отбором. Полагать, что эволюционно значимые наследственные свойства возникают помимо отбора - значит, принимать продукты эволюции за ее сырой материал.
Устойчивое (типичное) развитие. Теория устойчивого развития (или детерминации нормального фенотипа) составляет традиционную задачу экспериментальной эмбриологии. Свои попытки в этой области она начинала с уже упомянутой ранее преформистской (мозаичной) модели, предполагающей, что свойства взрослого организма могут быть сведены к сумме независимых линейных следствий из такой же суммы начальных причин (одними авторами они локализовались в ядре зиготы, другими - в цитоплазме). Эта механистическая модель, более всего связанная в эмбриологии с именем В. Ру, не выдержала экспериментальной проверки; явления регуляции при нарушениях эмбриогенеза показали, что итог развития до известной степени не зависит от вариаций составляющих его процессов [2184]. Что касается явлений цитоплазматической прелокализации зачатков, характеризующих "мозаичный" тип дробления, то они оказываются результатом предшествующих эпигенетических взаимодействий в эмбриогенезе и могут вновь сменяться в дальнейшем развитии регуляционными процессами [357, 922, 1199].
Все это привело к представлению о существовании целостной причины развития - сначала в виде энтелехии Г. Дриша [1524], а затем в форме концепции биологического поля [280]. Признание такого механизма детерминации означает, что признаки организма не имеют специфических коррелятов в зародышевой клетке и определяются взаимодействием всех ее частей; каждому частному акту дифференцировки предшествует детерминация целого. Отсюда следует, что предопределение целостных свойств зачатка должно происходить при неопределенном состоянии его элементов, как это и было впервые показано А. Г. Гурвичем [1683]. Это же логически вытекает из чисто сравнительного обобщения, сделанного столетием раньше и известного как одна из формулировок закона К. М. Бэра: "Общее в развитии предшествует специальному". Этот принцип определяющей роли целого - основа современного учения об эмбриональной детерминации [920, 922].
Если для эмбриологии приемлемо только такое решение, то оно не может быть иным и для генетики. Это выразилось в сходной эволюции ее представлений, приведших к возникновению гипотезы генного баланса [709, 1377], согласно которой каждый признак определяется всем геномом. Идея детерминации признака отдельным геном характеризуется поэтому как "ошибка Вейсмана" [353, с. 238], или как "генетика горохового мешка", отражающая воззрения раннего менделизма [594, с. 216], или же просто как некомпетентное мнение, ошибочно приписываемое генетике [1057].
Обусловленность нормального развития целостным начальным фактором, не сводимым к простой сумме его элементов, означает, что итог всего процесса является устойчивым, или эквифинальным по отношению к вариациям этих элементов. Последние детерминированы лишь статистически как совокупность, или, по А. Г. Гурвичу [280], "нормированы". Многочисленные проявления онтогенетической эквифинальности хорошо знакомы эмбриологам как в нормальном развитии (включая случаи разных путей размножения, дробления, гаструляции и т. п. при одинаковой взрослой организации у одних и тех же видов), так и при экспериментальных нарушениях, вплоть до регенерации целых особей из фрагментов специализированных тканей или самосборки зародышей из разделенных клеток (например, воссоединение клеток гаструлы морских ежей, ведущее к формированию нормальных личинок - плютеусов). Но обычно упускается из виду, что эта закономерность неизбежно должна иметь и генетическое выражение. Если устойчивое развитие определяется в конечном счете целостными свойствами зародышевой клетки, основанными на неопределенности (вариабельности) состояния ее частей, включая и строение ее хромосомного аппарата, то следует ожидать, что нормальный фенотип будет осуществляться в пределах широкого спектра геномных вариаций.
Остановимся на фактах, подтверждающих этот вывод. Общеизвестна генетическая гетерогенность ("насыщенность мутациями") природных амфимиктических популяций, скрытая под покровом адаптивного фенотипа и выявляемая инбридингом [1141]; непрерывная рекомбинация индивидуальных геномов в ходе смены поколений не меняет этих соотношений. Это означает, что отбор стремится преобразовать индивидуальные циклы развития в популяции таким образом, чтобы независимо от различий в их стартовых точках (зиготах) привести их к одному финалу - адаптивной норме [1178, см. рис. 23]. Но такие же соотношения должны существовать и там, где они не могут быть выявлены менделевским анализом, т. е. внутри автогамных и клональных популяций, которые на уровне разрешающей способности этого метода представляются генетически однородными. Это действительно доказывается многочисленными опытами по выращиванию жестко отселектированных сортов самоопылителей (злаков, бобовых) и партеногенетически размножающихся насекомых (например, тлей) в экстремальных условиях. Здесь неизменно обнаруживается разнообразие индивидуальных физиологических реакций, среди которых наиболее жизнеспособные варианты поддаются закреплению отбором [5, 909, 1149]. Хотя эти опыты истолковываются по-разному, вполне очевидно, что речь идет о генетической неоднородности, в норме скрытой канализирующими механизмами развития и получающей фенотипическое выражение в экстремальных условиях, где эти механизмы теряют эффективность. Результаты этих опытов принципиально ничем не отличаются от результатов, полученных при селективной стабилизации структурных или физиологических модификаций у ксеногамных организмов, например у дрозофилы [464, 2441], где они заведомо основаны на гетерогенности исходных линий. Все это позволяет сделать следующие выводы.
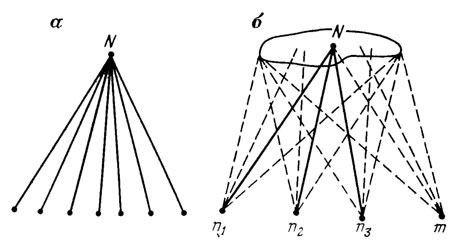
Рис. 23. Соотношения между зиготами и типами развития. По М. А. Шишкину. а - эквифинальность типичного развития нормальных гетерогенных зигот; б - ограниченность видоспецифичного пространства возможностей развития при любых вариациях зародышевого генома; сплошные линии - стабильный, прерывистые - лабильные пути развития; n1-n3 - нормальные зиготы; m - аномальная зигота; N - адаптивная норма
1. Создавая адаптивную норму, т. е. стабильный итог развития, естественный отбор неизбежно создает в ее основе генетическую вариабельность. Последняя является выражением помехоустойчивости нормального развития и не требует для своего объяснения дополнительных допущений - о силах, противодействующих отбору или о каких-то особых формах отбора (разнонаправленной, частотозависимой и т. п.), к которым вынуждена прибегать синтетическая теория, исходящая из селекции генов [7, 999, 1839]. Итог отбора при формировании адаптивной нормы - не сохранение фиксированной совокупности состояний геномных локусов, а создание новой организации генотипа, в рамках которой эти состояния непрерывно варьируют от особи к особи, не нарушая исхода развития.
2. Понятие нормального видоспецифичного генотипа не может быть описано в терминах генов, подобно тому, как свойства целого не сводимы к характеристикам его элементов [443]. Сущность генотипа как функциональной системы выражается лишь через характер контролируемого им индивидуального развития [461]. Такое определение по смыслу соответствует первичному иогансеновскому пониманию генотипа как "нормы реакции", или совокупности возможностей развития особи [443, с. 124].
Неустойчивое (аберрантное) развитие и эпигенетическая система. Вся область изменчивости, лежащая за пределами нормы и составляющая потенциальный материал отбора, представляет собой совокупность аберраций нормального развития. Теоретически для отдельного онтогенеза возможны два их источника - изменение условий развития и нарушение структуры самой зародышевой клетки, в первую очередь ее генома. В обоих случаях возникающее пространство аберраций не беспредельно; в частности, любой мутант остается особью своего вида [594]. Другой общей чертой подобных нарушений является снижение их устойчивости (наследуемости) по сравнению с нормой. Для мутаций это подтверждается всем опытом экспериментальной генетики [463], для уклонений же, индуцированных средой, отсутствие устойчивости самоочевидно. Эти факты и особенно явления параллелизма между мутационными и модификационными изменениями (фенокопиями) позволяют предположить, что в обоих случаях пространство аберраций имеет одни и те же ограничения, т. е. что возможности реализации любого мутантного генома не выходят за пределы, доступные его "нормальным" вариантам. Другими словами, все зиготы одного вида должны иметь одно и то же потенциальное пространство возможностей развития и различаются лишь по относительной вероятности их осуществления (см. рис. 23).
Правомерность этого вывода доказали феногенетические исследования Р. Гольдшмидта [1649, 1650, 1651], основанные на анализе фенокопий у дрозофилы. Выявленная им возможность фенокопирования практически любых мутационных эффектов вместе с их плейотроптными проявлениями позволила заключить, что развитие есть система с ограниченным набором возможных результатов и что все реализуемые в этих рамках аномалии, независимо от их начальных причин, имеют одну и ту же основу - неспецифические нарушения нормальной координации процессов онтогенеза (рассогласование скоростей реакций, изменение количеств, концентраций и времени взаимодействия реагирующих веществ).
Любой из таких количественных сдвигов, вызывающий при определенных пороговых значениях изменение фенотипа, может вызываться как мутациями, так и внешними факторами, т. е. действие мутаций на развитие выражает не их непосредственную специфику, а свойства самой реагирующей системы, которые могут быть выявлены и иными путями. Например, градациями температурных воздействий на личинку дрозофилы можно получить фенотипы всего ряда аллелей для мутаций Ваг или vestigial, причем усиление аномалии в этом ряду связано с отодвиганием момента начального отклонения развития на все более ранние стадии. Ясно, что характер нарушения определяется здесь изменением онтогенеза, а не состоянием хромосомного локуса. Во многих случаях природа количественного изменения, вызывающего подобную специфическую аномалию, выявляется более непосредственно. Например, действие мутации aristopedia у дрозофилы, превращающей перистый придаток антенны в структуру, подобную tarsus, основано на повышении темпов роста и сегментации имагинального диска антенны до уровня, свойственного диску конечности. При задержке роста с помощью колхицина развитие мутанта остается в рамках нормы [1649, 1651]. У мышей действие мутации Dh, вызывающей полидактилию задних конечностей, основано на замедлении гибели клеток апикального эктодермального гребня, вследствие чего пролонгируется его индуктивное действие на почку конечности. Напротив, мутация Os, ускоряющая отмирание гребня и сокращающая срок индукции, ведет к олигодактилии или даже редукции самой почки [505].
Если все нарушения фенотипа суть лишь реакции системы развития, непосредственно не отражающие специфики первичных возмущающих факторов, то одни и те же аномалии должны возникать при самых различных воздействиях на развитие. Это действительно имеет место. Многие типы шоков, задевающие один и тот же чувствительный период онтогенеза, дают одинаковый результат и, наоборот, один и тот же шок может вызывать качественно разные аномалии в зависимости от момента его действия и степени интенсивности [1651]. Кроме того, практически не известно мутаций, уникальных по своему эффекту, и в соответствии с этим исследователями выделяются "гетерогенные группы генов" по принципу общности выражения их мутаций, например группа minute у дрозофилы, охватывающая около 60 локусов в трех хромосомах [1057]. По существу, это лишь иная формулировка вывода Р. Гольдшмидта о неспецифическом воздействии гена на систему развития. Точно так же и при анализе природных фенотипических уклонений одни и те же их типы оказываются связанными с изменениями в разных хромосомах или просто с внешними воздействиями [449, 777]. Эта асимметрия причин и следствий ясно показывает, что при исследовании индивидуального развития мы имеем дело с системным объектом, обладающим устойчивым поведением, т. е. ограниченным набором возможных конечных состояний. Мы убеждаемся в том, что не только нормальный, но и любой аберрантный исход развития не может быть сведен к определенной совокупности элементарных начальных причин, а является выражением целостных свойств данной эпигенетической системы.
Значение выводов Р. Гольдшмидта для эволюционной теории самоочевидно. Они разрушили непреодолимую грань между мутационными и экзогенными изменениями, существовавшую в классической генетике, и свели их к реакциям единой системы развития, определяющей весь спектр осуществимых уклонений. Это открыло путь для совершенно нового понимания формообразовательной роли внешних факторов в эволюционном процессе, воплотившегося в теории И. И. Шмальгаузена. Был создан фундамент для идеи стабилизирующего отбора, рассматривающей наследственные изменения фенотипа не как эффекты определенных мутаций, а как продукты постепенного закрепления отбором целостных онтогенетических реакций, реализуемых гетерогенными носителями. Наследственность из статического и изначально данного свойства генов превратилась в исторически обусловленную характеристику системы развития. Неудивительно, что принцип неспецифического воздействия локусных мутаций на онтогенез занял важнейшее место в обосновании эпигенетической концепции И. И. Шмальгаузена [1196].
Чтобы понять, каким образом естественный отбор создает наследуемые эволюционные новшества, необходимо уяснить, как соотносятся лабильные (аберрантные) и устойчивые (нормальные) онтогенетические пути в пределах одной системы развития. Ответ на этот вопрос облегчает предложенная К. Х. Уоддингтоном [2441, 2442] концепция эпигенетического ландшафта, позволяющая описать главные свойства системы развития. Ограниченность и значительная дискретность пространства аберраций, вскрываемая феногенетикой, столь же очевидна и на эмбриологическом уровне, где она особенно наглядно демонстрируется поведением эксплантатов зародышевых тканей, способных дать в каждом случае лишь определенный набор дифференцировок [920]. Эта видоспецифичная совокупность возможностей развития зачатка (и в конечном итоге - всего организма), включающая нормальный для него путь и потенциальные уклонения, может быть наглядно выражена с помощью трехмерной модели эпигенетического ландшафта - в виде системы наклонных ветвящихся долин, дивергирующих во времени из начальной точки и символизирующих основные области притяжения, к которым стремится траектория развития (рис. 24-26).
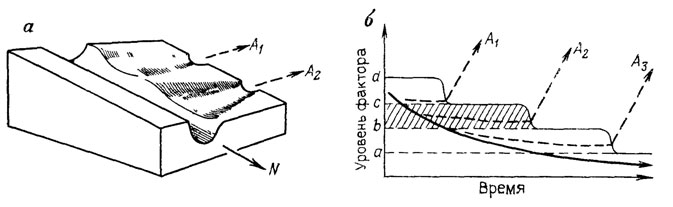
Рис. 24. Дискретность аберраций как выражение ступенчатого снижения помехоустойчивости нормального развития. По М. Л. Шишкину. а - участок эпигенетического ландшафта, показывающий перепады высоты стенок креода в местах ответвления аберративных долин, б - зависимости характера аберрации от соотношения между уровнем повреждающего морфогенетического фактора и устойчивостью креода, в промежутке между двумя пороговыми уровнями устойчивости (заштрихован) колебания меры фактора не изменяют типа развития, a, b, c, d - пороговые уровни устойчивости последовательных отрезков креода, А1-А3 - аберративные пути развития, N - нормальный ход изменений уровня фактора и типичный путь развития
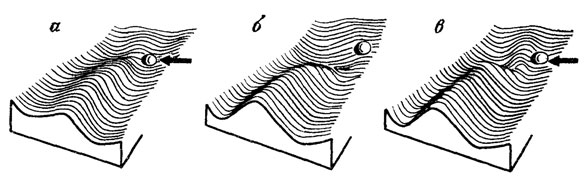
Рис. 25. Соотношения между эпигенетическим ландшафтом и характером повреждающего воздействия. По К. Х. Уоддингтону, интерпретация изменена. а - уклонение развития на боковую долину за счет сильного внешнего воздействия (стрелка), б - такое же уклонение под действием сильной мутации (разрушение стенки креода), в - промежуточное состояние
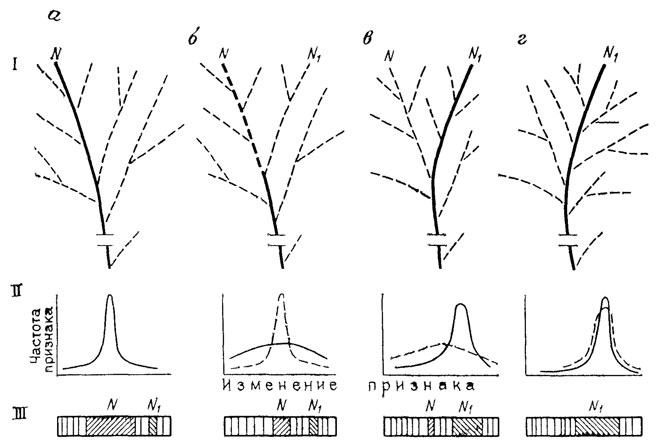
Рис. 26. Перестройка системы развития в ходе становления новой элементарной адаптации. По М. А. Шишкину. I - изменения эпигенетического ландшафта, II - изменение дисперсии преобразуемого адаптивного признака в популяции (сплошная линия - вариация признака на данной стадии, пунктирная - на предыдущей); III - изменения модификационных спектров; а - канализированное развитие адаптивного фенотипа N, соответствующего главной полосе в спектрах и пику кривой вариации, б - дестабилизация развития, ведущая к снижению частоты фенотипа N и уменьшению его роли в спектрах; в, г - две последовательные стадии канализации онтогенетической траектории N1, с развитием вокруг нее новой сети аберративных долин. Фенотип N1, из неустойчивого морфоза превращается в новую адаптивную норму, прежняя норма становится морфозом или исчезает. Жирными линиями выделены креоды; заштрихованы типы реакций, соответствующие последовательным нормам
Помехоустойчивость типичного онтогенетического пути, ведущего к адаптивной норме, выражается в резком заглублении его долины (креода). Все случайные возмущения, не выходящие за пределы последней, оказываются зарегулированными, т. е. отклонившаяся траектория вновь "скатывается" на следующих стадиях в сторону русла. При наличии нескольких адаптивных норм в ландшафте имеется соответствующее число таких креодов, выбор среди которых каждый раз контролируется либо действием среды (при модификационном полиморфизме), либо закономерным рекомбинированием хромосомного аппарата (например, при определении пола). Всякое необратимое уклонение процесса развития в сторону одной из более сглаженных аберративных долин связано с преодолением более или менее высокого защитного порога, отделяющего ее от русла креода (см. рис. 24, а). Области ответвления этих долин соответствуют понижениям стенок креода и, таким образом, отражают фазы относительной неустойчивости, т. е. те чувствительные периоды, с воздействием на которые связано получение экспериментальных аберраций.
Как уже отмечалось, ни один из возможных путей в системе развития не контролируется каким-то особым зародышевым фактором. Весь эпигенетический ландшафт как целое, или фазовый портрет системы [66], представляет собой "свойство высшего порядка", основанное на взаимодействии всех элементов генотипа [2441, с. 34]. Поэтому возникновение такой системы невозможно приписать элементарным мутациям. Элементарные воздействия на эту систему либо забуфериваются ею, либо меняют в ней выбор реализуемой онтогенетической траектории, но они не способны изменить исторически сложившуюся структуру самой системы. Все они, независимо от их природы, могут вызывать лишь два типа количественных сдвигов, нарушающих развитие: 1) снижение помехоустойчивости того или иного участка (временного отрезка) креода ниже критического порога, допускающего регуляцию к норме; 2) повышение интенсивности повреждающего фактора выше порога, допустимого для данного участка. Поскольку возможности регуляции в целом снижаются в ходе онтогенеза, то чем сильнее будут эти нарушения, тем более ранний из чувствительных отрезков креода (т. е. мест ответствления аберративных долин) окажется задетым и тем более глубоко дивергентным и дискретным по отношению к норме окажется реализованный путь развития (см. рис. 24, а, б). Отсюда следует, что дискретность фенотипических изменений, вызываемых мутациями одного и того же хромосомного локуса, выражает не свойства его аллельных состояний как таковых, а специфику реагирования всей системы развития, способной отвечать качественно различным образом на разные степени повреждающего воздействия. К этому выводу фактически привели еще ранние исследования Р. Гольдшмидта по механизму определения пола и С. Райта [2505] по феногенезу мутаций альбинизма, показавшие, что в основе аллельных изменений лежат непрерывные градации одного и того же морфогенетического фактора (например, количества вещества или скорости реакции), действующие с пороговым эффектом.
Исходя из сказанного, можно описать возможные результаты воздействия отдельного повреждающего фактора на индивидуальные циклы развития в нормальной гетерогенной популяции. Все зиготы одного вида принадлежат к одной и той же системе развития, различаясь лишь по относительной вероятности осуществления отдельных траекторий. Для нормальных зигот эти различия минимальны, т. е. путь, ведущий к адаптивной норме, для них наиболее устойчив (канализирован) и обнаруживает лишь частные вариации, выражаемые на эпигенетическом ландшафте как локальные различия в глубине креода (высоте его защитных порогов). Для воздействий, способных вызвать уклонения, возможны три основные ситуации.
Первая - резкое изменение структуры генома, приводящее к столь сильному снижению защитного порога (на определенном участке креода), что независимо от его вариаций развитие неизменно уклоняется на один и тот же боковой путь (см. рис. 25, б). Это - идеальная мутация, наиболее удобная для генетического анализа, т. е. такое локусное изменение, которое при введении в любой вариант генома вызывает с максимальной вероятностью определенную аномалию развития. Однако результат все же должен оказаться не вполне устойчивым, поскольку сглаженность аберративных долин ландшафта сама по себе исключает эффективную регуляцию ими онтогенетических траекторий. И действительно, на практике выражение даже сильных "сырых" мутаций остается изменчивым, т. е. неидеально наследуемых [1079]*.
* (Таковы, в частности, генетические аномалии человека. Обычно они показывают изменчивость выражения и могут иногда не проявляться у гомозигот даже в случае доминирования, например полидактилия [245].)
Вторая (противоположная) ситуация - предельно сильное внешнее воздействие на ход развития, преодолевающее любой порог его устойчивости и приводящее в данном цикле развития к тому же результату, что и сильная мутация (см. рис. 25, а). Это - классическая фенокопия.
Наконец, третий вариант охватывает всю область промежуточных состояний между двумя ранее описанными крайними случаями, когда характер и сама возможность уклонения зависят от конкретного соотношения между особенностями эпигенетического ландшафта и условиями развития (см. рис. 25, в). Действие одной и той же мутации будет либо лежать ниже порога нарушения, либо преодолевать этот порог в различных точках креода, ведя к разным фенотипическим уклонениям - в зависимости от индивидуальных особенностей ландшафта (определяемых исходной конституцией генома) и колебаний факторов среды. И, наоборот, одни и те же уклонения должны возникать при различных комбинациях внешних и внутренних условий развития. В этих случаях говорят о мутациях с неустойчивым выражением и проявлением, т. е. не показывающих при анализе правильного менделевского наследования.
Справедливость этой модели подтверждается реальной картиной неопределенной изменчивости, наблюдаемой в природных популяциях. Общеизвестен факт отсутствия или редкости в них той категории аберраций, которая может быть охарактеризована как доминантные мутации, с хорошим проявлением [242]. При этом даже крупные однотипные аберрации при анализе оказываются связанными с разными хромосомами или индуцированными извне, как например, Abnormal abdomen у дрозофилы [см. 449]. При наличии достаточно больших выборок таких фенотипов выявляются самые различные их градации по устойчивости наследования - от соотношений, близких к менделевским, до полной потери проявления (например, фенотип "пятнистые глаза" у дрозофилы [2]). Поэтому внутри таких групп изореагентов авторы часто вообще не решаются провести границу между наследственными (мутационными) и ненаследственными (модификационными) изменениями [43], или же ищут ее между линиями с минимальным наследованием аберрации и линиями с полным его отсутствием.
Вполне очевидно, что речь здесь идет о границе, которой нет в природе. Все фенотипы одного класса представляют собой варианты реализации одной и той же онтогенетической траектории, различающиеся по степени устойчивости и обусловленные самыми различными сочетаниями индивидуальной генетической конституции и факторов среды. Понятия "мутации" и "модификации" относятся, по существу, к разным объектам: первое - к сравнению особей, второе - к сравнению возможностей развития одной особи. Все одинаковые фенотипы (как и любые другие) всегда генетически неидентичны* и потому могут рассматриваться как скрытые мутанты по отношению друг к другу независимо от результатов гибридного анализа. И наоборот, любой фенотип, оцениваемый на основе такого анализа как мутантный, представляет собой лишь одну из возможностей развития в пределах эпигенетической системы данной зиготы, т. е. одну из альтернатив (модификаций) по отношению к нормальной для вида траектории. Последнее особенно очевидно для тех случаев, когда возвращение на эту траекторию легко осуществимо путем изменения условий развития (например, у дрозофилы - мутации pennant, vestigial, Abnormal abdomen и др. [1192]).
* (Это касается выборок из любых популяций, в том числе и клонов, поскольку ошибки репликации генома неизбежны при любом способе клеточных делений. Представление о полной генетической однородности внутри клонов основаны на том, что их скрытая гетерогенность не улавливается обычными методами в нормальных условиях развития. Однако она обнаруживается при выращивании клонов в экстремальных средах, нарушающих эквифинальное (канализированное) развитие.)
Эпигенетическая система как объект воздействия отбора. Все сказанное позволяет оценить роль элементарной мутации в эволюционном процессе. Мутация не может дать стабильного эффекта, ибо всякое индивидуальное уклонение от нормального пути развития есть нарушение устойчивости. Тем более она не может создать и ничего нового, поскольку ни один путь развития, реализуемый при ее участии, не выходит за пределы пространства возможностей, свойственного данной эпигенетической системе. Поэтому утверждение, что эволюция идет за счет случайных мутаций, по выражению К. Х. Уоддингтона, "представляется пустым" [2441 с. 188]. Как бы ни был широк диапазон случайных изменений на нуклеотидном уровне, возможности их воздействия на онтогенез всегда ограничены исторически сложившимися свойствами системы развития.
Подлинное эволюционное изменение - это изменение структуры (ландшафта) самой системы, т. е. в конечном счете реорганизация контролирующего ее генотипа. Результатом такой перестройки является превращение одной из прежних аберративных областей эпигенетического пространства в область наиболее вероятных событий. Иначе говоря, в ней должен сформироваться новый креод вместе со специфическим для него рисунком главных потенциальных уклонений (см. рис. 26). Фенотипически это должно выражаться в преобразовании адаптивной нормы и спектра ее аберративной изменчивости. Ближайший путь для достижения такого результата - канализация одной из уже существующих в данной системе аберративных траекторий. И если сама система есть продукт отбора, то и любое дальнейшее ее преобразование должно осуществляться тем же путем. Другими словами, эволюционная стабилизация аберрантного фенотипа достигается отбором среди его носителей.
Прямым подтверждением этого являются опыты К. Х. Уоддингтона по генетической ассимиляции (стабилизации) структурных морфозов у дрозофилы [2441], а также многие другие аналогичные эксперименты, связанные с закреплением физиологических аберраций и приводящие к созданию линий, устойчивых к непривычным условиям развития, например [464, 1147, 1148]. Все они, по существу, сводятся к перестройке эпигенетического ландшафта путем отбора на устойчивость отдельных его траекторий.
Так, в опытах К. Х. Уоддингтона исходным материалом послужили крыловые морфозы у дрозофилы (например, фенотипы dumpy и bithorax), полученные с помощью теплового воздействия на нормальных личинок. Сначала отбирались носители одного из морфозов, затем полученные от них личинки вновь подвергались тепловому шоку, а взрослое потомство - отбору в прежнем направлении. После ряда поколений отбора резко повышалась устойчивость такого фенотипа как к внутренним, так и к внешним факторам развития. Первое выражалось в возрастании частоты его наследования (т. е. в снижении чувствительности его развития к эффекту рекомбинации родительских геномов), второе - в уменьшении его зависимости от температурного фактора (аберрация начинает передаваться и при отсутствии шока). В итоге первично индуцированное изменение превращается в наследственное и при контрольном скрещивании с исходной линией выщепляется с той или иной частотой.
Таким образом, возникла новая организация генотипа, обусловившая канализированное развития прежней аберрации. Этапы перестройки эпигенетического ландшафта здесь достаточно очевидны. В исходной линии все его варианты характеризуются одним и тем же креодом, ведущим к норме, или дикому фенотипу N (см. рис. 26, а). В условиях шока устойчивость креода резко падает, т. е. ландшафт размывается, и наряду с нормой реализуются различные индивидуальные аберрации, например N1 (см. рис. 26, б). Отбор в пользу последней еще более стирает различия в вероятности осуществления прежней нормы и ее уклонений - уже безотносительно к температурному фону развития. В то же время постепенная стабилизация отбираемого фенотипа (N1) преобразует шаг за шагом его траекторию в наиболее устойчивый путь развития (см. рис. 26, в, г).
Этот обобщенный механизм элементарного эволюционного изменения может быть описан несколько иначе - по изменению разнообразия индивидуальных систем развития на отдельных этапах становления новой нормы. Поскольку возможности осуществления одних и тех же путей развития для разных зигот неодинаковы, то в любом конкретном диапазоне условий, выходящем за рамки обычных, каждая из них обнаруживает свой собственный спектр реализуемых фенотипических уклонений (морфозов). С этой точки зрения популяция нормальных зигот может быть представлена как серия разнородных модификационных спектров, в которых основную часть всегда составляет адаптивный фенотип (рис. 27).
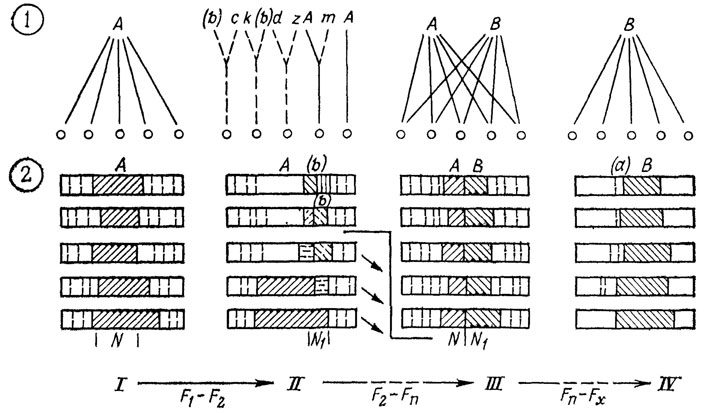
Рис. 27. Преобразование индивидуальных модификационных спектров в ходе элементарного сдвига адаптивной нормы. По М. А. Шишкину. Цифры в кружках: 1 - соотношение между зиготами и реализуемыми фенотипами; 2 - модификационные спектры, характеризующие серию зигот в одном и том же интервале условий; I - стабильное (эквифинальное) осуществление рядом зигот исходной адаптивной нормы А в средних допустимых для нее условиях N; II - дестабилизированное развитие таких же зигот в интервале экстремальных условий N1 среди реализуемых уклонений (b), c, d, k, z морфоз (b) имеет адаптивное преимущество; III - стабилизация морфоза (b) и превращение его в адаптивную модификацию В в рамках новой полиморфной нормы АВ; IV - дальнейшая стабилизация фенотипа В на фоне утраты прежней нормы или сохранения ее в качестве неустойчивого морфоза (a). F1-F2-Fn-Fx - смена поколений; заштрихованы осуществленные типы развития; их элиминируемые варианты показаны нисходящими стрелками F1-F2 - вскрытие изменчивости, F2-Fn, Fn-Fx - этапы стабилизирующего отбора
В обычных условиях все зиготы развиваются эквифинально, реализуя норму (см. рис. 27, I). По мере изменения среды в сторону критического порога канализированное развитие сменяется разнонаправленным, т. е. возникает все большее число морфозов в соответствии со спецификой индивидуальных спектров (см. рис. 27, II). Это - стадия вскрытия "мобилизационного резерва" индивидуальной изменчивости [1186, 1192]. При сохранении таких условий в ряду поколений начинается отбор в пользу наиболее жизнеспособного морфоза, ведущий к постепенному росту его устойчивости и дестабилизации прежней нормы (см. рис. 27, II, III). Элиминация остальных типов аберрантных реакций вначале малоэффективна, поскольку они вновь возникают в потомстве отбираемого варианта ввиду его слабой онтогенетической устойчивости. Но по мере стабилизации этого фенотипа его наследование становится более однозначным и он (в случае ксеногамного размножения) все в большей степени поглощает в скрещиваниях остальные аберрации, остающиеся неустойчивыми.
Это закрепление адаптивно ценной реакции, превращающее ее в новую норму, приводит к тому, что в модификационных спектрах последовательных поколений зигот она занимает все большее место за счет старой нормы. Таким образом, обе нормы сосуществуют на определенном этапе в индивидуальных спектрах как две адаптивные модификации, реализуемые в зависимости от колебаний условий (см. рис. 27, III), пока новая, наконец, полностью не возобладает. Возможность реализации прежнего нормального фенотипа если не исчезает совсем, то низводится до уровня аберрантной атавистической реакции (см. рис. 27, IV).
Наиболее наглядное экспериментальное подтверждение такого хода преобразований дают примеры стабилизации физиологических реакций, поскольку они обычно позволяют судить о возможностях реагирования одних и тех же особей в разных условиях. Кроме того, объектом отбора здесь является не произвольно выбранный признак (как в случае морфологических уклонений), а жизнеспособность особи в целом, что приближает ситуацию к природной. Таковы, например, опыты М. М. Камшилова [464] по воспитанию холодоустойчивости у дрозофилы, И. В. Кожанчикова [498] по выращиванию жуков-листоедов на непривычном корме и в особенности эксперименты Г. X. Шапошникова по смене хозяина у тлей [1147, 1148]. Во всех этих случаях нарушение среды развития приводит к реализации экстремальных неустойчивых физиологических состояний, из которых наиболее жизнеспособное превращается путем отбора в стабильную адаптацию.
Первой реакцией исходной популяции на подобную смену условий обычно является резкое повышение изменчивости, включая дифференциальную плодовитость и выживаемость. Это - описанная выше фаза перехода от эквифинального развития к разнонаправленному, т. е. вскрытие мобилизационного резерва (см. рис. 23, 27,1, II). Далее вновь наблюдается повышение однородности физиологического реагирования, очевидно, за счет стабилизации наиболее жизнеспособного варианта. При этом, как показывают опыты с тлей Dysaphis anthrisci majkopica [1148], ряд поколений не отдает предпочтения старому или новому типу условий. Это - переходный этап сосуществования старой и новой норм в виде двух адаптивных модификаций (рис. 27, III). В дальнейшем стойкая способность к прежней реакции обычно утрачивается, а новая норма стабилизируется как единственная (рис. 27, IV). Обособление ее носителей от исходной линии иногда может достигать уровня репродуктивной изоляции, как это наблюдалось в опытах Г. Х. Шапошникова [1150].
Во всех подобных случаях имеет место создание отбором именно нового приспособления, а не просто сохранение "готовой" адаптивно ценной вариации, как это часто полагают. Например, в опытах с Dysaphis вплоть до 8-11 поколений воспитания на новом хозяине не существовало особей, предпочитающих его старому [1148], точно так же, как в опытах с охлаждением личинок дрозофилы исходная популяция заведомо не содержала особей, устойчиво толерантных к снижению температуры или предпочитающих его [464, 465]. Отбор ведет не к сохранению гена, "детерминирующего" новое свойство, или же определенного варианта генома (набора аллельных состояний локусов), а к преобразованию организации всего генотипа данной линии, т. е. к созданию нового пространства вариаций индивидуальных геномов, допускающего эквифинальное (устойчивое) осуществление изменившейся нормы (см. рис. 23, 27).
Для рассмотренных экспериментов известна масса природных аналогий, связанных чаще всего с "привыканием" различных форм насекомых к пестицидам, где в итоге отбора возникают резистентные расы [353, 1509]. Нет никаких оснований полагать, что процесс их становления отличается от вышеописанного.
Изложенная концепция преобразования нормы созвучна со многими частными обобщениями классической генетики. В них, во всяком случае, признается, что отдельный локус не детерминирует признака [709] и что изменение последнего требует перестройки генотипа [594]. Однако об этих выводах забывают, когда дело касается биохимических мутаций, так как их часто рассматривают именно как доказательство скачкообразного возникновения новых устойчивых приспособлений. В этом, по существу, отражается противоречие между моргановской генетикой, декларирующей множественную обусловленность признака, и принципом молекулярной генетики "один ген - один фермент".
Примеры такого рода достаточно известны. Например, путем пересева штамма кишечной палочки, живущего на галактозе, на среду с лактозой можно выделить жизнеспособную в новых условиях мутацию, причем методом реплик демонстрируется, что она существовала уже в исходной культуре. Налицо как будто бы спонтанное появление новой адаптации. Но нетрудно видеть, что в действительности здесь выделено не стойкое приспособление, а элементарный морфоз, входящий в спектры реагирования ряда клеток наряду с типичной реакцией. Он реализуется у мутантов с той или иной вероятностью только на соответствующем провокационном фоне, тогда как на исходной галактозной среде осуществляется нормальная функция; иначе такие клетки погибли бы до пересева. Выбор между двумя реакциями определяется здесь типом среды. Закрепление же вскрытой формы реагирования может происходить лишь в ходе размножения измененного штамма в новой среде по мере отбора среди множества индивидуальных клеток. При высокой скорости размножения бактерий такой отбор идет крайне быстро, затемняя исходную неустойчивость морфоза.
Этот вывод о путях биохимической эволюции основан не только на рассмотренной эпигенетической концепции отбора. Его подтверждают факты, свидетельствующие о поливариантности и множественной обусловленности всех ступеней матричного синтеза [311, 441]. Вследствие этого одна и та же матрица ДНК соответствует различным модификациям конечного продукта в зависимости от состояния аппарата синтеза. И наоборот, различные нарушения кода (мутации) могут в эпигенезе регулироваться к норме - как на уровне трансляции (в частности изменение свойств тРНК, приобретающей способность "узнавать" чуждые кодоны), так и при посттрансляционных изменениях (например, комплементация гомологичных полипептидных субъединиц, исправляющая либо их первичную структуру, либо конформацию, от которой зависит биохимическая активность белка). Все это наблюдается как при нормальной работе клеток, так и при экспериментальных воздействиях, причем неспецифическими агентами регуляции ("фенотипической супрессии") мутаций могут быть антибиотики, изменения температуры, рН осмотического давления и т. д. [311, 442]. Они воздействуют, несомненно, на самые различные элементы системы синтеза - например, на рибосомные белки, состояние которых влияет на супрессорные свойства тРНК [441].
Все это показывает, что, несмотря на резкое сокращение дистанции между локусом и признаком, отличающее процессы матричного синтеза от целостного онтогенеза, и, конечно, неизмеримо большую роль локуса в определении конечного результата, здесь действуют в принципе те же самые законы, что и в индивидуальном развитии вообще (см. рис. 23). Функциональная устойчивость реализуемого продукта основана не на идеальной стабильности нуклеотидной последовательности матрицы, а на способности целостной системы синтеза к саморегуляции. Синтезируемая макромолекула является итогом взаимодействия всех элементов такой системы и ее адаптивные свойства устойчивы (эквифинальны) по отношению к вариациям этих элементов. Яркое проявление этих соотношений - способность белковой молекулы к сохранению своей функциональной значимой пространственной структуры при множестве вариаций ее первичной структуры (обусловленных ошибками кода, а также транскрипции и трансляции) [1078]. Таким образом, принцип однозначного соответствия между локусным "геном" (цистроном) и его продуктом, лежащий в основе представлений о чисто мутагенном происхождении биохимических адаптации, представляет собой грубое упрощение.
В действительности главная проблема биохимической эволюции - это вопрос о том, каким образом среди всех возможных модификаций того или иного конечного продукта закрепляется лишь один определенный "фенотип", наиболее соответствующий адаптивным требованиям [1078, с. 113]. Нетрудно понять, что отбор, благоприятствующий данной конечной модификации (для белков - посттрансляционной, для РНК - посттранскрипционной), будет стремиться сохранять все относящиеся к ней молекулы, независимо от вариаций исходной для них матрицы, и что среди этих вариаций будут получать предпочтение те, на основе которых данный тип продукта реализуется с наибольшей надежностью. В итоге отбор по результатам синтеза будет неизбежно перестраивать лежащую в его основании матрицу. По существу, это тот же механизм, который управляет любым элементарным эволюционным изменением, т. е. закрепление адаптивно ценной реакции, реализуемой гетерогенными носителями.
Можно с большим основанием полагать, что явления эпигенетической доработки, свойственные всем типам РНК и различным белкам (например, инсулину) и выражающиеся в расчленении, укорочении, агрегации и других закономерных изменениях синтезированных молекул-предшественников, исторически представляют собой стабилизированные "рекапитуляции" тех неустойчивых адаптивно ценных изменений, которым подвергались эти молекулы при нарушениях внутриклеточной среды у предков и которые были выделены отбором из множества других разнонаправленных модификаций. При этом доработка иРНК, предшествующая трансляции полипептидной цепи, может с большой вероятностью рассматриваться как результат ускорения и упрощения преобразований, происходивших когда-то с данным типом белка на посттрансляционном этапе, - подобно тому, как в эволюции "макроонтогенеза" происходит эмбрионализация развития дефинитивных структур.
Таким образом, общность законов, управляющих как целостным онтогенезом, так и внутриклеточными событиями, касается не только осуществления нормальных созидательных процессов, но и способа их эволюции. Система матричного синтеза, как и эпигенетическая система в целом, включает в каждом случае специфический спектр возможных траекторий с разными конечными продуктами, выбор среди которых зависит от внешних и внутренних факторов [441, с. 112]. Наибольшая устойчивость нормальной траектории в обоих случаях основана на регулирующих взаимодействиях внутри системы. Любое стойкое эволюционное изменение конечного продукта должно обеспечиваться такой же согласованной регуляцией, и оно невозможно без постепенной перестройки отбором всей системы синтеза применительно к новым требованиям. Поэтому биохимическая эволюция на основе случайных элементарных мутаций так же немыслима, как и эволюция организмов.
Итак, можно считать, что эпигенетические преобразования, лежащие в основе элементарного сдвига адаптивной нормы и рассмотренные нами на примере ряда экспериментальных моделей, заключают в себе закономерности достаточно общего характера. Каждый такой сдвиг, закрепляющий одну из аберративных траекторий исходной системы развития, распадается на два этапа: разнонаправленное модифицирование (дестабилизацию) прежней нормы и стабилизацию одного из возникших уклонений. Таким образом, в эволюции оба процесса "непрерывно кооперируются" [1192, с. 315]. Первому этапу соответствует индивидуализация онтогенетического развития (см. рис. 27, I, II), второму - его унификация, т. е. создание нового креода (см. рис. 27, III, IV; ср. рис. 26, в, г).
Отсюда следует, что между последовательными историческими состояниями устойчивой нормы (см. рис. 27, I, IV) всегда лежит период неустойчивости, преодолеваемый в ряду поколений посредством отбора (см. рис. 27, II, III). Из этого ясно, что так называемый движущий отбор не представляет собой самостоятельного феномена. Он выражает итог длинного ряда чередующихся фаз стабилизации и дестабилизации нормы. Эволюция предстает здесь как процесс непрерывной репарации онтогенетической устойчивости, нарушаемой последовательными изменениями среды. И. И. Шмальгаузен [1192] справедливо подчеркивал, что правильнее говорить о движущем и стабилизирующем эффектах отбора, чем о двух его типах. Можно уточнить, что речь идет, в сущности, о двух разномасштабных и разнокачественных измерениях одного процесса. Описывая его по совокупному итогу, мы говорим о движущем отборе, но его элементарный механизм всегда составляет стабилизация уклонений. С этих позиций становится понятным истинный смысл дарвиновского определения отбора как "сохранения полезных индивидуальных изменений" [310, с. 139]. Мелкие индивидуальные аберрации сами по себе не могут устойчиво наследоваться; единственный путь их сохранения - отбор на стабильность их онтогенетического осуществления.
О сущности естественного отбора. Все сказанное позволяет уяснить созидательную роль естественного отбора. Она остается непонятной, пока его материалом признаются готовые "наследственные изменения", понимаемые как проявления мутаций генеративных клеток. При таком подходе объяснение наследуемости эволюционных новшеств становится излишним, ибо в ней видят просто, имманентное свойство генов, существующее изначально и не зависящее от отбора. На долю последнего остается лишь роль фильтра таких изменений или в лучшем случае средства, позволяющего их комбинировать. Вся процедура "создания" отбором очередного элементарного новшества приравнивается здесь к одноактному выбору его причинного фактора, за которым должно следовать автоматическое воспроизведение нового признака в поколениях потомков. Ничего нового при этом не возникает. Неудивительно, что в представлениях синтетической теории эволюции, отводящей отбору эту весьма неспецифическую роль, он является лишь одним из многих ее "исполнителей" - наряду с дрейфом генов, изоляцией и т. п.
Напротив, для эпигенетической теории устойчивость воспроизведение (наследуемость) изменений - это и есть то, что требует объяснения на основе принципа отбора. В ее представлениях истинная причина действенности отбора заключается не в изначально высокой наследуемости сохраняемых им вариаций, а в принадлежности их к ограниченному пространству аберраций, свойственных данной системе развития. В результате спектр этих вариаций повторяется в поколениях, обеспечивая стабильность потенциального субстрата отбора без стабильности индивидуального наследования. Отбор в пользу любой из этих вариаций ведет не к сохранению ее "детерминирующих генов", а лишь к увеличению частоты ее гетерогенных носителей и оставляемых ими гамет. Действенность отбора в каждом его шаге оценивается здесь по двум поколениям, и ее мерой является эффективность воспроизведения отобранного фенотипа у его непосредственного потомства. Это воспроизведение вначале бывает крайне неустойчивым, т. е. отобранные изореагенты практически повторяют в своем потомстве весь тот спектр видоспецифичных аберраций, из которого они сами были перед этим отобраны. Отбор на этом этапе почти "черпает решетом воду". Каким же образом растет его эффективность?
Каждый элементарный шаг отбора (т. е. итог двух его поколений) - это преимущественное сохранение особей, сумевших повторить фенотип своих ранее отобранных родителей, несмотря на комбинирование их гамет при скрещивании и различные генетические изменения в процессе самого гаметообразования (мейотическая рекомбинация и ошибки репликации). Поэтому история любого фенотипа, сохраненного длительным отбором, - это цепь последовательных испытаний его носителей на способность воспроизводить самих себя в условиях непрерывного изменения пространства вариаций их геномов. Чем большим числом поколений отбора отделены такие носители от исходной группы изореагентов, давшей им начало, тем больший размах генетических изменений оказалась способна выдержать данная линия, не меняя своего фенотипа. Это означает, что генотип вновь созданной линии (представленный множеством индивидуальных вариантов) перестраивается отбором в направлении все более помехоустойчивого осуществления данного фенотипа. Соответственно последний начинает все эффективнее поглощать в скрещиваниях остальные аберрации, свойственные данной системе развития, т. е. реагирует на гибридизацию с их носителями, как на регулируемые онтогенетические помехи. Сохраняемое уклонение становится, таким образом, все более однозначно наследуемым, превращаясь в новую норму.
Это преобразование нормы посредством отбора, по существу, представляет собой выражение общей способности системных объектов релаксировать возмущения, т. е. изменяться целенаправленно. Восстановление равновесия, или "поиск цели" [1224], осуществляется системой путем последовательной коррекции ее состояния, ведущей к затуханию исходного возмущения. Именно такая ситуация, но только связанная с качественным изменением самой системы, возникает в процессе перестройки адаптивной нормы. При переходе популяции организмов в экстремальные условия их система развития дестабилизируется и реализует неупорядоченные индивидуальные флуктуации. Дальнейшее выживание системы в новых условиях зависит от того, удастся ли ей стабилизироваться в каком-либо из этих изменчивых состояний.
Этот поиск нового равновесия осуществляется посредством преимущественного сохранения (отбора) индивидуальных вариантов развития, реализующих наиболее жизнеспособную флуктуацию. Процесс отбора как раз и составляет цепь затухающих циклов коррекции с обратной связью, ведущих к стабилизации новой нормы (точки равновесия). Каждый акт сохранения носителей адаптивно ценной аберрации есть сдвиг состояния системы в сторону будущего равновесия; "шумы" при воспроизведении этого фенотипа в следующем поколении означают новое отклонение от равновесия; очередной акт просеивания вновь сдвигает облик популяции в направлении будущей нормы и т. д. до тех пор, пока каждое новое поколение не станет фенотипически однородным и подобным родительскому. Прежняя флуктуация становится равновесным состоянием.
Таким образом, созидательная роль отбора, как и каждого творческого процесса, заключается в конечном счете в "запоминании случайного выбора" [478], которое выражается в данном случае в выборе одной из относительно равновероятных флуктуации системы развития и в превращении ее в новую устойчивую норму. Последняя на всем протяжении своего становления играет роль "цели", определяющей направление коррекции свойств системы в ходе ее преобразования отбором.
Эпигенетическая эволюция и геологическая летопись. Изложенная концепция [1777-1182] рассматривает эволюцию как преобразование системы развития, определяющей специфическое для вида пространство потенциально реализуемых фенотипов. Этот взгляд получает ныне все большее признание [1261. 1262, 1741, 1895, 2115, 2375]. Поскольку любой из осуществимых путей развития является целостной реакцией системы и не сводим к действию каких-либо ее элементов, то тем самым эволюция фенотипов не может быть описана в терминах генов и их частот. Для эпигенетической концепции главными рабочими понятиями являются адаптивная норма и ее аберрации, или, иначе говоря, равновесное состояние системы и пространство ее флуктуации. Только нормальному фенотипу свойственно устойчивое осуществление и тем самым - устойчивое наследование, включая и правильные (менделевские) числовые соотношения в потомстве при наличии у вида нескольких норм, т. е. в случае адаптивного полиморфизма. Сырой материал эволюции, или неопределенная изменчивость, согласно теории, не имеет в общем случае этих свойств и рассматривается как пространство флуктуации системы. Утверждается, что любое элементарное новшество стабилизируется на основе одной из таких флуктуации, т. е. возникает вначале как изменчивая по проявлению и выражению вариация прежней нормы.
Из этого представления о механизме эволюции следует, что изучение динамики изменчивости на палеонтологическом материале в принципе может позволить проследить само становление эволюционных новшеств, а не просто их распространение в ископаемых популяциях (понимаемое обычно как распространение соответствующей "мутации", несущей данное полезное изменение).
Такой подход ставит перед палеонтологией нетрадиционные задачи. Обычно основным объектом ее внимания являются преемственные типы нормальной организации, описываемые в таксономических категориях и представляющие по определению продукты стабилизированного развития. Эта практика обусловлена самим характером геологической летописи, отдающей предпочтение формам, максимально устойчивым во времени и пространстве, т. е. реализуемым в онтогенезе с наибольшей вероятностью. Таким образом, как правило, анализируются лишь "готовые" продукты эволюции, но не процессы их преобразования один в другой. Этим объясняется скептическое отношение многих биологов к возможностям палеонтологии судить о событиях микроэволюционного порядка и в конечном счете - о движущих силах эволюции вообще [cp. 952, 1745].
Хотя интерес к изучению динамики изменчивости у ископаемых форм неуклонно растет [2419], это пока оказывает мало влияния на господствующие представления о механизмах эволюции живой организации. В любой области знаний способ истолкования фактов определяется исходными теоретическими представлениями, и соответственно в палеонтологии эволюция вариаций обычно трактуется с позиций господствующей синтетической теории. Хорошо известным примером такого истолкования являются исследования Дж. Симпсона [952, 2287]. В них легко обнаруживаются те же противоречия между теоретическими принципами моргановской генетики и фактическим пониманием эволюционных событий, которые свойственны и другим сторонникам синтетической теории [594, 1509]. С одной стороны, признается, что гены - не детерминанты признаков, а лишь элементы в системе развития, что их действие взаимосвязано и т. д. [2287, с. 60, 64], а с другой, утверждается, что появление новых адаптивных черт есть результат отбора соответствующих мутаций [там же, с. 87, 88]. Появление конкретного нового признака (например, гребешка "кроше" или металофа в эволюции зубов лошадеобразных) прямо отождествляется Дж. Симпсоном с мутацией, а его дальнейшее развитие - с последовательными мутациями в том же или других локусах [2287]. Таким образом, фактически признается однозначная связь генов и признаков. И хотя Дж. Симпсон считает почти невероятным появление новой полно развитой адаптивной структуры или новой систематической группы за счет одной мутации (в противовес мнению О. Шиндевольфа [2223]), он, тем не менее, полагает, что историю каждого приспособления можно проследить назад до первой "ключевой" адаптивной мутации, некогда подхваченной отбором. Все это лишний раз показывает, что в основе "синтетического" мышления, по существу, лежит синтез вейсмановского преформизма (ген определяет признак) и мутационистских идей.
В еще более упрощенной форме мутационистские представления выступают во многих частных палеонтологических работах, пытающихся объяснить появление эволюционных изменений с помощью генетических понятий. Так, Б. Паттерсон [2069] приписывает единичной мутации появление когтей у кайнозойских млекопитающих-тениодонтов из подсемейства Conoryctinae, положившее начало развитию подсемейства Stylinodontinae. Таким же образом истолковано появление экзостозов на костях у вымершей птицы-отшельника с о. Родригес [2059], развитие диморфизма раковины в позднеплейстоценовой истории моллюска Cryptopecten vesiculosus [1713], унисериальной рабдосомы - у монограптидных граптолитов [1783], а также многие другие факты внешне скачкообразного появления новых признаков в геологической летописи. Подобные истолкования применяются и к более постепенным изменениям, например в случае направленной эволюции у фораминиферы Cycloclypeus, объясняемой как результат мутационного давления со стороны последовательности множественных аллелей [1910].
Для объяснения различных изменений у ископаемых форм широко используются и такие понятия синтетической теории, как "поток генов" [2419], "генетический груз" [2021], "генетическая революция" [1719] и т. д. С точки зрения эпигенетической концепции все эти истолкования произвольны уже потому, что любая попытка описать изменения признаков в терминах локусных генов подразумевает наличие между теми и другими упорядоченных соответствий. Поскольку же такое соответствие формально отрицается генетикой [709], то не существует и возможности использовать ее язык для универсального описания фенотипических изменений.
Напротив, понятия эпигенетической теории относятся к фенотипам, т. е. к той реальности, которая непосредственно изучается палеонтологом. Ее основное предсказание, что эволюция нового признака должна выражаться в его стабилизации и соответственно снижении его изменчивости, подтверждается многими палеонтологическими фактами [2287, 2419], нашедшими отражение также и в руководствах по эволюционной генетике [353]. Эту закономерность можно проиллюстрировать рядом примеров. Так, в двух стратиграфически последовательных популяциях раннепермского стегоцефала Diplocaulus magnicornis отмечается значительная стабилизация морфы "crook" (сильно вытянутые и отогнутые назад заднебоковые углы черепа), первоначально редкой и неустойчивой в выражении [2027]. У пермских лабиринтодонтов степень врастания сонных артерий в парасфеноид часто варьирует, тогда как у триасовых форм она стабилизирована уже на ювенильных стадиях [1176]. У миоценового вида пелециподы Glycimeris таким же образом фиксируется морфа "rugose" [200]. Среди граптолитов у лландоверийского представителя Petalograptus отмечены вариации в сторону укорочения тек, приобретающие у более поздних петалограптид закономерный характер [1985]. В эволюции кайнозойских морских ежей-клипеастроидов постоянным отверстиям и вырезкам панциря у поздних форм предшествуют более вариабельные состояния у ранних [2362]. У раннекарбоновых брахиопод рода Eomarginifera отмечаются резкие вариации в степени упорядоченности игл в основании ушек, в расположении сосочков на висцеральном диске и т. д.; но у более позднего представителя маргиниферид правильное расположение этих структур становится нормой [272].
Эпигенетическая концепция позволяет сделать и ряд других предсказаний, в значительной мере подтверждаемых на ископаемом материале, - например, о возрастании вариабельности перед вымиранием филумов или при росте темпов эволюции [ср. 1410, 2419]. Оба случая сводятся к ситуации, при которой изменение условий обитания делает невозможным устойчивое онтогенетическое осуществление прежней организации, и начинается поиск приемлемого для стабилизации варианта среди реализуемых уклонений. В обоих случаях достигается лишь относительная стабилизация, позволяющая персистировать в узком интервале условий, нарушение которых вновь вынуждает систему развития продолжать поиск. Независимо от того, достигает ли она в итоге высоко стабильной организации или прекращает существование, предшествующий отрезок ее эволюции должен характеризоваться большим удельным весом поколений с изменчивым типом развития, а также многообразием попыток стабилизации, запечатленных в виде последовательных или параллельных морфологических вариантов с коротким периодом геологического существования.
В целом изучение фактов с позиций изложенной концепции открывает перед палеонтологией значительные возможности для познания механизмов эволюции формообразовательных систем, которые и составляют истинную основу процесса микроэволюции.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'