
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
7.2. Ранние этапы развития жизни на Земле (Б. С. Соколов, М. А. Федонкин)
Самая уникальная особенность Земли как планеты Солнечной системы - ее стратисфера, т. е. оболочка водно-осадочного происхождения, мозаичная по составу и структуре, но непрерывная в геологическом времени. Она заключает наиболее полную запись событий, касающихся истории как самой Земли, так и Вселенной на протяжении последних четырех миллиардов лет.
До недавнего времени исторический этап развития Земли определялся началом кембрийского периода (570 млн лет). Дифференцированный органический мир кембрия, в том числе скелетные беспозвоночные всех типов, воспринимался как загадочный "популяционный взрыв". В настоящее время следы активной жизнедеятельности организмов и сами их остатки (от прокариотных безъядерных организмов до неминерализованных слоевищ растений и отпечатков бесскелетных многоклеточных животных) установлены во всем осадочном докембрии. Следы былых биот находят в столь древних отложениях, что невольно возникает вопрос - не древнее ли жизнь самой Земли?
Палеонтология докембрия ставит перед исследователями вопросы подобного рода, хотя вряд ли она способна разрешить их. Источником знаний о происхождении жизни являются скорее химия и молекулярная биология, биогеохимия и, возможно, астрономия (см. 2.7). В последние десятилетия экспериментально доказано абиогенное возникновение не только простейших органических веществ, обнаруженных теперь и в космосе, но и таких сложных веществ, какими являются аминокислоты, сахара, короткие пептиды и даже нуклеотиды. Однако в практическом и даже в теоретическом аспекте пока трудно представить себе скачок, в результате которого химическая эволюция перешла в эволюцию биологическую. Вероятно, поэтому, особенно в последние годы, возрождается интерес к биокосмическим идеям В. И. Вернадского.
Сведения о жизни в докембрии получают в результате исследований фитопланктона, акритарх и других органостенных микрофоссилий, сохранившихся в аргиллитах и сланцах, а также ископаемых строматолитов (их распределения, морфологии, микроструктуры и палеоэкологии) и минерализованных остатков, клеток ископаемых прокарит (например, цианобактерий) из строматолитовых окремненных пород. Кроме того, изучают изотопный состав докембрийских пород и их керогеновых компонентов, особенно изотопов углерода и серы. Однако эти свидетельства жизни на современном уровне изученности распределены неравномерно по стратиграфической шкале докембрия. Мы еще очень мало знаем о жизни в архее, много больше - о биотитах протерозоя, особенно его поздней части.
По данным Дж. У. Шопфа [2240], синтез всех данных палеобиологии, палеобиогеохимии и геохимии позволяет предполагать следующий "сценарий" ранней биологической эволюции. Жизнь появилась на Земле более 3,5 млрд лет назад и, вероятно, ранее этого рубежа в органическом мире уже были развиты такие явления, как анаэробная хемогетеротрофия, анаэробная хемоавтотрофия и анаэробная фотоавтотрофия. Аэробная фотоавтотрофия и амфиаэробный (факультативный) метаболизм, возможно, уже существовали 2,9 млрд лет назад. Микроорганизмы, зависимые от аблигатного аэробиоза, появились 1,7-1,5 млрд лет назад.
Наиболее многообещающим подходом к проблеме реконструкции биологической и связанной с ней геологической истории Земли является выяснение сути и последовательности биогеохимических циклов, катализаторами которых были живые организмы, в том числе литотрофные бактерии [384]. Биохимические циклы элементов прокариотной экосистемы обусловили однонаправленное изменение продуктов дегазации планеты. Многообразие реакций, осуществляемых бактериями, допускает предположение о множестве трофических систем для различных начальных условий (что подразумевает полифилию). По мнению Г. А. Заварзина, взаимодействие живых организмов с продуктами дегазации является ключевым процессом в совместной геологической и биологической эволюции, вплоть до стабилизации системы при развитии биологических циклов. Новые эквариотные организмы вкрапливаются в уже существующую систему трофических связей.
Докембрий составляет около семи восьмых времени существования нашей планеты и приблизительно пять шестых времени, представленного палеонтологической летописью. Наиболее древними свидетельствами жизни, возможно, следует считать формации углеродистых отложений глубокого докембрия, хотя основная масса углерода в докембрийских породах находится в рассеянном состоянии. Изотопный состав углерода из архейских и протерозойских углеродистых отложений может свидетельствовать, скорее, о его органическом происхождении [948].
В пользу этого предположения говорят данные по содержанию редких элементов - спутников органогенного углерода - в древнейших углеродистых породах [978]. Однако возможность миграции углерода, связь содержания этого элемента в породах с цикличностью вулканических процессов и ряд других явлений показывают, что проблему органического углерода в древнейших толщах нельзя считать решенной окончательно.
Широко распространенные в докембрии железистые формации, в частности тонкослоистые железистые кварциты (джеспилиты), также нередко рассматриваются как биолиты - продукты жизнедеятельности железобактерий. Однако достаточно строгих свидетельств в пользу этой гипотезы пока нет. Слоистость джеспилитов - не обязательно результат биологической или биохимической деятельности. Разнообразные физические и химические процессы, периодически происходящие в окружающей среде, могут быть причиной подобной ритмичности в слоистости осадка; например, суточные изменения освещенности или сезонные флуктуации температуры могут влиять на скорость осадконакопления, химический, минералогический и гранулометрический состав осадка.
Едва ли не самое видное место среди памятников биологической активности организмов докембрия занимают строматолиты. Одна из существенных особенностей образования строматолитов состоит в том, что фоссилизации в большинстве случаев подвергались не сами трихомы нитчатых и сфероидальные формы цианобактерий-строматолитообразователей, а обволакивающая их скопления слизь, фиксирующая на поверхность наслоения или в толще слизистых пленок хемогенный, а возможно, и терригенный осадок преимущественно карбонатного состава. Нити цианобактерий редко инкрустируются карбонатом кальция. Иногда у современных строматолитов в результате жизнедеятельности синезеленых водорослей выпадают лишь разрозненные зерна кальцита в слизи. Эти и другие данные, полученные при изучении современных строматолитов у побережий Багамских островов [1965], Австралии [1889] и Бермудских островов [1628], во многом позволили понять природу древних строматолитов.
Недавно была сделана попытка измерить скорость роста современных строматолитов [1426]. Использование изотопа 14С в качестве индикатора возраста проб, отобранных вдоль вертикальной оси одиночных строматолитовых построек приливно-отливной зоны, показало, что скорость их вертикального роста составляет от 250 до 400 мм за 700-800 лет. Датирование по изотопу 210Pb свидетельствует, что в последние годы строматолиты растут со скоростью около 100 мм за 50-100 лет. Если учесть, что мощные толщи протерозоя нередко сложены в основном строматолитами или содержащими строматолиты породами, то по аналогии с современными можно определять скорость осадконакопления.
Изучение строматолитов и микрофитолитов в разрезах верхнего докембрия СССР привело к выделению последовательностей характерных таксонов, вполне сравнимых даже для отдаленных районов. Были выделены естественные этапы развития фитолитов и их комплексы, включающие по 10-15 формальных видов. Все это позволило решать задачи расчленения и корреляции верхнедокембрийских (рифейских) отложений в возрастных объемах до 300 млн. лет - для докембрийской стратиграфии это крупное достижение. В результате широких исследований фитолитов было предложено трехчленное [755, 760], а затем и четырехчленное [482] деление верхнего протерозоя СССР. Аналоги строматолитовых комплексов рифея и его подразделений - фитем - устанавливаются сейчас и в других районах мира (Шпицберген, КНДР, МНР, КНР, Индия, Африка, Австралия).
Интересно, что выводы, полученные в результате исследований современных строматолитов, исключают возможность применения докембрийских строматолитов для широких стратиграфических построений, если первые и последние являются полными аналогами [943]. Действительно, в формировании современных строматолитов участвует не один вид водорослей, а ассоциация микроорганизмов, состав которой обычно определяется экологическими факторами, хотя в каждой ассоциации выделяются один-два доминирующих вида. Основные черты морфологии строматолитов обусловлены абиотическими факторами; скоростью и направлением движения воды, интенсивностью осадконакопления, а для литоральных форм, кроме того, высотой прилива и т. д. В то же время текстурно-структурные особенности построек определяются систематическим составом водорослей-строматолитообразователей и непосредственно не зависят от экологических факторов.
Эти другие данные, полученные при изучении современных строматолитов, даже в том случае, если они не применимы в полной мере к их докембрийским аналогам, позволяют наметить некоторые пути исследований последних. Теперь, чтобы продвинуть знание о строматолитах как об индикаторе геологического времени в докембрии (который крайне необходим), нужно поставить задачу не в традиционном плане - строматолиты и стратиграфия, а в плане получения разнообразной информации об их экологии, морфологических и структурных реакциях построек на абиотические факторы, которые можно диагностировать известными физическими и геохимическими методами, и т. п. Не менее важна и интересна попытка выяснить соотношения водорослевых сообществ в самих строматолитах и во вмещающих породах, возможную корреляцию сохранившихся строматолитообразователей с характером микроструктуры строматолитов и пр. Наиболее продуктивны эти исследования при систематическом использовании актуалистического подхода. Трудно ожидать, что сильно зависимые от изменений условий среды строматолитовые постройки, созданные медленно эволюционирующими прокариотами, могли хронологически обновляться в темпах, сравнимых с темпами эволюции фанерозойских организмов. Однако не менее трудно допустить, что разрешающая способность строматолитов и фитолитов в стратиграфическом плане исчерпывается четырьмя фитемами рифея.
Большое значение для понимания природы строматолитов и роли бактерий в постройках последних имеет изучение сообществ организмов в современных газогидротермах. Эти сообщества рассматриваются как персистентные [384, 385], хоть и с существенными ограничениями. Заслуживает внимания сравнительное исследование древнейших ископаемых микрофоссилий и микроорганизмов из водорослево-бактериальных пленок термальных источников, в том числе таких явлений, как посмертные изменения цианобактерий и окремнение современных организмов [238, 542].
Едва ли не самой перспективной группой для изучения древнего органического мира и для биостратиграфии протерозоя в целом являются акритархи. Это собирательное название применяется для разных групп микрофоссилий недостаточно ясной природы. Им охватываются в первую очередь целостные одноклеточные и агрегатные микроорганизмы, обладающие необычайно стойкой неминерализованной наружной оболочкой, и дисперсные остатки спор и грибов, имеющие покровные образования того же типа. Несомненно, в состав акритарх входят и цианобактерий - вероятно, одна из наиболее древних групп.
Одним из труднейших палеоальгологических вопросов в изучении древнейших микрофоссилий является вопрос об их систематической принадлежности. В палеонтологии пока не удается использовать такой признак высшего таксономического значения, как состав пигментов, поэтому приходится опираться на косвенные суждения, вытекающие из некоторых морфологических признаков, которые далеко не всегда надежны. Обычно более определенны заключения о синезеленых водорослях в целом, хотя общеизвестна их поразительная морфологическая изменчивость на протяжении жизненного цикла и в зависимости от условий среды и характера симбиоза. Что же касается пиррофитовых, красных, зеленых и некоторых других отделов водорослей, то хотя их существование в докембрии не вызывает сомнений, конкретные определения, особенно фитопланктона, практически всегда затруднены. Поэтом чаще всего приходится говорить о морфологических группах - различных сфероморфных, филаментарных и пр.
Сложность биологической интерпретации докембрийских микрофоссилий состоит и в том, что палеоальгологи, если они вообще ищут аналогов среди представителей современного фитопланктона, обращаются прежде всего к атласам или к изучению микроорганизмов, в то время как микрофоссилии следовало бы сравнивать не с живыми, а с погибшими организмами. Изменения, которые претерпевают многие микроорганизмы после гибели, в большинстве случаев существенно влияют на их облик, причем настолько, что идентифицировать их часто бывает невозможно. Отсюда вытекают крайне необходимые для палеоальгологов работы актуопалеонтологического характера, в частности изучение (в том числе экспериментальное) постмортальных изменений представителей разных групп микромира в различных: условиях.
Наиболее древней группой акритарх являются сфероморфиды. Они встречаются в осадочных толщах с возрастом 3,3 млн лет, а возможно, и более древних [1049, 1050, 1051, 1052, 1053]. Диаметр их обычно не превышает 30 мкм, хотя для современных прокариот верхний предел не превышает 20-23 мкм. На протяжении нижнего протерозоя наблюдается постепенное увеличение численности и морфологического разнообразия сфероморфид. Расцвет этой группы приходится на средний и поздний рифей. В рифее отмечается массовое развитие одноклеточных колониальных водорослей, скопления которых извлекаются из пород в больших количествах. С верхнего рифея появляются многие морфологические группы акритарх. Однако наибольшего обилия и разнообразия они достигают в нижнем кембрии. Так, к настоящему времени на территории Восточно-Европейской платформы установлено более 100 видов кембрийских акритарх, которые служат надежной основой детального расчленения и корреляции вмещающих отложений [860].
Акритархи извлекаются из осадочных пород путем растворения последних в сильных кислотах и последующей сепарации на центрифугах, а изучаются, как правило, в препаратах. Попытки исследований акритарх под сканирующим электронным микроскопом пока немногочисленны, однако этот метод открывает новые возможности исследования структуры их стенок.
По-видимому, столь же разнородной, но в целом близкой к акритархам группой докембрийских окаменелостей являются микрофоссилии, которые прекрасно сохранились в сингенетичных или раннедиагенетических кремнях. Стенки их сложены органическим веществом, которое в большинстве случаев полностью замещено кремнеземом. Эти микрофоссилии изучаются в тонких петрографических шлифах. Фиксированное состояние делает их в некотором отношении более надежной группой, чем акритархи, так как исключает возможность контаминации, столь обычной в проницаемых для вод осадочных горных породах. С другой стороны, именно кремнезем способствует лучшей сохранности рассматриваемых микрофоссилий, защищая их от различных агентов выветривания. Может быть, поэтому микрофоссилий, сохранившиеся в кремнях, пока являются наиболее древними из прямых доказательств существования жизни на организменном уровне в глубоком докембрии.
Классическим местом изучения данных микрофоссилий стала Барбетонская горная страна в Южной Африке. Здесь из черных кремней ряда формаций группы Онфервахт (возраст 3,3-3,5 млрд. лет) описаны микроскопические структуры в виде темных шариков (отдельных, иногда вытянутых в цепочки) и темных нитей. Несмотря на то что их сохранность намного хуже, чем у микрофоссилий из более молодых протерозойских отложений, в их морфологии можно усмотреть черты сходства с остатками сине-зеленых водорослей [1264, 1978, 1998]. Условия осадконакопления, в которых формировалась группа Онфервахт, интерпретируются как мелководные с незначительным поступлением обломочного материала с суши [1897]. Из формаций Кромберг и Цварткопи описано несколько ассоциаций микрофоссилий, в том числе сферические, нитевидные и в виде "колоний" или скоплений.
Палеобиохимические исследования аморфного органического вещества керогена, широко распространенного в породах группы Онфервахт, показали, что в его состав входят различные биополимеры, в том числе спорополленин, характерный для экзины спор современных многоклеточных растительных организмов [1383]. Изотопные исследования углерода из вмещающих пород свидетельствуют скорее о биогенной природе микрофоссилий, среди которых путем статистического анализа размеров можно выделить несколько популяций мелких сфероидов, нитчатые несегментированные образования и нити, образованные сцепленными сфероидами, напоминающими как водоросли, так и бактерии.
Близкая по возрасту микробиота описана из баритоносных отложений Норт Поул в Западной Австралии [1246]. Здесь обнаружены пять разновидностей микрофоссилий, весьма напоминающих обнаруженные в группе Онфервахт. Возможно, наиболее древними окаменелостями на Земле сейчас являются микрофоссилий из кварцитов серии Исуа юго-западной Гренландии [2084]. Морфологически они напоминают остатки клеток, в том числе одиночные клетки, нити, агрегаты или "колонии" клеток. Структурный и химический анализ этих образований, осуществленный с помощью лазерного микроанализатора, позволил предположить, что они могли относиться к дрожжам и другим эвкариотам. Однако позднее этот вывод был подвергнут сомнению.
Морфология большинства архейских микрофоссилий проста. Преобладают, как видно из приведенных примеров, сферические формы, которые различают статистически на основе доминирующих диапазонов размерности. Нередко многие из сфероидов сидят на матрице измененного аморфного органического вещества, что делает их похожими на современных коккоидных цианофитов, выделяющих большое количество клейкой слизи. Многие современные цианофиты имеют нитевидный габитус в виде цепочки клеток, заключенных в слизистый чехол. В ассоциации микрофоссилий из группы Онфервахт найдены цепочки клеток двух размерностей, но они, по-видимому, не имели чехла, хотя один фрагмент чехла там был обнаружен [1977]. Если микрофоссилий из серии Онфервахт Южной Африки и из серии Исуа юго-западной Гренландии действительно являются остатками сине-зеленых водорослей, то мы вправе считать, что по крайней мере 3,5 млрд. лет назад на Земле начался или уже шел процесс образования фотосинтетического кислорода. Не исключено, что в архее могли существовать организмы, не имеющие сейчас потомков, бактерии или даже такие цианофиты, которые подобно некоторым современным их представителям могут жить в анаэробных условиях. Достаточно уверенно мы можем говорить лишь о том, что эти архейские организмы, еще не имевшие обособленного клеточного ядра, обладали развитой системой обмена веществ и способностью к размножению. Энергетика этих организмов, вероятно, опиралась на процессы брожения, так как кислород должен был связываться прежде всего неорганическими реакциями.
Биологическая интерпретация микрофоссилий остается одной из наиболее острых проблем в палеонтологии докембрия. Возможно, определенную роль в решении этого вопроса сыграют геохимические исследования, связанные с реконструкцией кислородной эволюции атмосферы Земли. Пока на этот счет существуют взаимоисключающие гипотезы. Согласно одной из них [78, 1330], содержание кислорода в атмосфере медленно возрастало, достигнув современного уровня лишь в течение фанерозоя. По мнению авторов этой гипотезы, прокариотная система не могла обеспечить более 1% современного содержания свободного кислорода в атмосфере.
Вместе с гем, как полагают Л. Беркнер и Л. Маршалл (там же), нижний уровень содержания кислорода в атмосфере, составляющий 0,01% современного, мог существовать изначально в результате фотолиза воды. Связывая эволюцию органического мира Земли с изменением состава ее атмосферы, эти исследователи намечают два уровня содержания свободного кислорода в атмосфере (1 и 10% современного его содержания), которые считаются критическими для биологических процессов. Первый из этих уровней в эволюции атмосферы ("точка Пастера") пройден в конце докембрия, что послужило причиной появления в короткий срок разнообразных групп многоклеточных животных. Имеются, однако, сведения о существовании кислородной обстановки в глубоком докембрии. Данные об изотопных соотношениях серы в протерозойских метаосадочных породах свидетельствуют, что по крайней мере 3 млрд лет назад уже существовала атмосфера, близкая по составу к современной [174, 757]. Имеется и ряд геологических аргументов в пользу этой гипотезы [1006, 1505].
Балансовые расчеты М. Шидловски показывают, что 80%, современного объема кислорода появилось очень рано в истории Земли. Биологическим свидетельством окислительной обстановки в раннем докембрии, возможно, являются микроскопические звездчатые образования Eoastrion, описанные наряду с другими микрофоссилиями из широко известной микробиоты серии Ганфлинт в Северной Америке [1305, 1882]. Возраст вмещающих их отложений около 2 млрд лет. Сходные формы отмечались и в отложениях близкого и более древнего возраста Австралии [1976, 2451] и Африки [1366]. Морфологически Eoastrion идентичен ныне живущим марганецосаждающим бактериям Metallogenium, которые впервые были описаны из пресноводных озер Карелии [881]. Если мы допустим, что не только морфологически, но и биохимически Eoastrion идентичны Metallogenium, иначе говоря, это организмы, прошедшие без существенных изменений через большую часть истории биосферы нашей планеты [383, 1434, 1442], то в существовании богатой кислородом обстановки в далеком докембрии можно не сомневаться, так как в природе представители Metallogenium встречаются при разной концентрации кислорода, но всегда в аэробных условиях [263]. Без кислорода окисление железа и марганца невозможно, а сохранение самих "паучков" этих микоплазмоподобных организмов возможно только в окислительной обстановке. Хотелось бы заметить, что реконструкции подобного рода не учитывают одного важного обстоятельства: локально могут накапливаться весьма высокие концентрации многих элементов, в том числе и кислорода, в результате деятельности бактерий и других организмов, например фотосинтезирующей микрофлоры.
Конечно, довольно трудно допустить существование неизменной в течение более чем 2 млрд лет какой-либо группы организмов, в то время как на Земле коренным образом менялись как биос, гак и абиотическая среда. К тому же бактерии вообще имеют крайне ограниченное пространство логических возможностей по морфологическим признакам (мы имеем в виду форму клетки, а не строение клеточной стенки, мембран и т. д.), поэтому вполне возможно неоднократное повторение одной и той же формы клетки в разных группах этого царства на протяжении геологической истории.
Интересно, что в отношении природы самого металлогениума высказывались определенные сомнения, является ли он живым существом или продуктом жизнедеятельности организма. Что же тогда говорить о критериях биогенности древнейших окаменелостей? Живые системы в общем случае определяют на основании того, что они способны двигаться, развиваться, размножаться, мутировать и осуществлять всевозможные акты жизнедеятельности, т. е. на основании таких процессов, которые трудно или невозможно идентифицировать на ископаемом материале, особенно среди столь примитивных окаменелостей, которые известны в глубоком докембрии. Что касается формы, то, как правило, чем сложнее морфология, тем больше вероятность, что мы имеем дело с биологическим объектом. В случае несомненных окаменелостей - чем больше морфологических характеристик, тем больше уверенности в интерпретации. Тем не менее множество разнообразных абиогенных объектов имитируют биологические конфигурации [2379].
Следует отметить, что проблема отличия живого от неживого отнюдь не тривиальна в отношении многих современных объектов, изучаемых биологами. В палеонтологическом аспекте этот вопрос стоит гораздо более остро и, вероятно, требует наряду с теоретическими разработками обобщения накопленного опыта в области сравнительного изучения остатков организмов, псевдоокаменелостей и артефактов. Большую пользу могут принести актуопалеонтологические наблюдения и эксперименты, которым пока почти не уделяется внимания. Несмотря на то что концепция биогенности в палеонтологии почти не разработана, мы считаем полезным привести хотя бы некоторые из критериев органической природы ископаемых остатков [1748].
1. Свидетельства выполнения жизненных функций: а) фоссилизация остатков организмов в стадии выполнения каких-либо актов жизнедеятельности, например, деления клетки, рождения, питания, пищеварения и т. д.; б) морфологические или материальные (вещественные) свидетельства выполнения каких-либо биологических функций, например, разнообразные следы передвижения, питания, жилища, а также копролиты и т. д.
2. Клеточная дифференциация (и отсюда - морфологическая сложность) вместе с морфологическим постоянством большого количества индивидов, включая набор онтогенетических стадий.
3. Сходство с живыми или известными вымершими формами и соответствие определенной эволюционной последовательности.
4. Морфологическое разнообразие в ассоциациях, представляющих сообщества нескольких или многих морфологически четко различимых групп (видов), каждая из которых имеет предположительно собственные экологические требования.
5. Изобилие, географическая протяженность ареала и ограниченный стратиграфический интервал распространения.
6. Характер сохранности.
7. Химические свидетельства, такие, например, как производные биогенных макромолекул (изопреноиды, порфирины и др.), изотопное фракционирование (13С/12С или 34S/32S соотношения).
8. Геологическая вероятность.
Эти критерии, к которым следовало бы добавить еще один - экосистемное положение, - вполне применимы для таксонов относительно продвинутых организмов и для более поздних эпох геологической летописи, если использовать их в комплексе. Однако эффективность их использования резко снижается, когда мы имеем дело с остатками из докембрийских толщ. Здесь окаменелости не только редки и мало изучены, но и морфологически более примитивны. Так, до сих пор мы не имеем четких критериев для определения природы мельчайших сфероидов, которые часто встречаются в шлифах и в препаратах: они могут быть как остатками организмов, так и физико-химическими структурами. С большей уверенностью удается судить о биологическом происхождении подобных объектов, напоминающих клетки, если можно идентифицировать вероятные внутренние структуры и выявлять митотические последовательности. Если же подобные наблюдения не могут быть сделаны, то сфероидальные микроструктуры лучше называть "возможно биогенными". Пожалуй, наиболее плодотворной будет разработка критериев биогенности для относительно узких групп морфологически сходных ископаемых объектов.
Трудности в изучении простых микрофоссилий с органической стенкой связаны еще с тем, что различные группы исследователей применяют разные подходы и методики работы от начальных стадий исследования до стадии интерпретации и классификации результатов. При сопоставлении результатов нередко возникают затруднения, так как диапазон исследований очень велик - от чисто биологических до биостратиграфических.
Биостратиграфическая школа в изучении микрофоссилий (прежде всего акритарх) сформировалась в нашей стране более 30 лет назад. Наиболее яркие ее представители, в частности, С. Н. Наумова, Б. В. Тимофеев, Н. А. Волкова и другие, используя палинологические приемы при выделении микрофоссилий из терригенных пород (преимущественно темноцветных сланцев), добились важных результатов в исследовании биостратиграфии верхнедокембрийских и нижнепалеозойских отложений. Опираясь на чисто морфологический подход, сторонники биостратиграфической школы пытались классифицировать материал и сравнивать его с теми или иными группами органического мира. Однако биологическая интерпретация микрофоссилий достаточно редко обсуждалась в деталях, так же как и влияние фациального контроля на распределение и сохранность микрофоссилий. Определенные сложности были вызваны изъянами стратиграфической основы, особенно в докембрийских толщах. Наиболее сложные проблемы, остро стоящие и сейчас, касаются классификации и диагностики древнейших микрофоссилий.
Существенно иное, палеоботаническое направление исследований микрофоссилий зародилось в США, начиная с работ Ч. Уолкотта [2446], его расцвет связан описанием известной микробиоты из кремнистых толщ Ганфлинт [1305]. Рассматриваемое направление характеризуется особым вниманием к биологической интерпретации и эволюционному значению докембрийских микрофоссилий. Наибольшее число исследований было сосредоточено на раннедиагенетических кремнях, которые образуются в строматолитовых карбонатах. Доминирующими организмами в современных строматолитах являются сине-зеленые водоросли, поэтому по аналогии с ними предполагается, что микрофоссилии в древних строматолитах были цианофитами [2236]. На этой основе большинство строматолитовых микробиот было включено в различные таксономические категории цианофит, а остальные были отнесены к различным таксонам внутри нескольких классов прокариот [2232, 2242]. Однако анализ критериев диагностики докембрийских микрофоссилии показывает, что абсолютно надежных доказательств систематического положения строматолитовых микрофоссилии пока нет [1837, 2243].
Наиболее полное представление о составе ассоциаций микрофоссилии из докембрийских толщ могла бы дать комбинация методики мацерации и петрографический метод (изучение микрофоссилии в тонких шлифах) с отбором проб из возможного широкого спектра фаций. Изучение влияния фаций на распределение и сохранность докембрийских микробиот в терригенных толщах [1507, 2110] показало, что микрофоссилии, которые считались типично строматолитовыми, регулярно встречаются в составе микробиот, характерных для сланцев. Кстати, в этой ситуации возникли сложности с классифицированием микрофоссилии. Например, один и тот же экземпляр следовало бы относить и к роду в системе цианофит или хлорофит (следуя школе палеоботаников), и одновременно к роду в системе акритарх (по биостратиграфической систематике). Это разногласие свидетельствует о глубокой противоречивости и слабой разработке систематики древнейших микрофоссилии.
В целом докембрийская часть палеонтологической летописи характеризуется прогрессивным увеличением размеров и разнообразия микрофоссилии [2237], хотя классифицирование таксонов на основе статистических тестов размерности сфероидов или нитей является достаточно произвольным. Сейчас ни у кого не вызывает сомнений, что самые ранние формы являются прокариотами, а поздние докембрийские микрофоссилии во многих случаях представлены эвкариотами. Наибольшие же дискуссии вызывает вопрос, когда возникли эвкариоты и как их распознавать. У некоторых сфероидов наблюдается темное пятно, которое часто интерпретируют как ядро клетки, однако ряд авторов высказывает предположение, что оно может отвечать сжавшейся мембране или коллапсировавшей цитоплазме, или пигментированным вакуолям. В общем же в литературе преобладает мнение, что эвкариоты появились на Земле (1,5±0,3) · 109 лет назад.
Одним из наиболее заметных рубежей в развитии микромира Земли было начало протерозоя, или афебия, которое датируется примерно в 2,6±0,1 млрд лет. С этого времени огромного масштаба достигает деятельность железобактерий, следы которой известны сейчас в Кривом Роге на Украине, в Канаде, Австралии, Южной Африке и других регионах. К возрастному интервалу 2,1-1,7 млрд лет относятся многочисленные углеродсодержащие формации в пределах всех известных докембрийских щитов и массивов, известных на нашей планете [943]. Время массового развития строматолитов охватывает период от 2,6 до 0,5 млрд лет, хотя максимум приходится на интервал 0,9-0,6 млрд лет.
![Таблица. Характеристика прокариот и эукариот [2240]](pic/000022.jpg)
Таблица. Характеристика прокариот и эукариот [2240]
С рубежом 1,9-1 млрд лет (границей раннего и позднего афебия) в серии Ганфлинт (южная часть шт. Онтарио, США) связаны один из наиболее примечательных находок [1302-1306, 1434-1447, 1747, 4421 и др.]. Помимо строматолитов здесь известны десятки родов, по-видимому, фитопланктонных организмов шарообразной, звездчатой и нитчатой форм, причем последние со следами перегородок (клеток?). В составе этой биоты идентифицируются цианофиты, бактерии и, видимо, грибы. В СССР примерно такое же стратиграфическое положение занимает ятулий Карелии. Возраст ятулийской седиментации около 2 млрд лет или несколько меньше [867]. Здесь встречаются богатейшие проявления жизнедеятельности водорослей - строматолиты [123]. В углистых сланцах Шуньги обнаружен обильный фитопланктон [1050]. О микрофлоре позднего афебия или среднего протерозоя (1,9-0,1 - 1,6-0,1 млрд лет) поступают разнообразные данные и из других районов мира (Канады, Гренландии и т. д.).
Исключительный интерес как биологический мегахрон представляет рифей. Ни одно из стратиграфических подразделений докембрия не характеризуется столь обширным распространением различных породообразующих строматолитов и микрофитолитов. Карбонатные фации этого возраста на всех континентах почти повсеместно содержат строматолитовые постройки. Наиболее значительные результаты стратиграфического и структурного изучения строматолитов были достигнуты в СССР, однако лишь недавно началось исследование самих организмов-строматолитообразователей [385].
Ряд исследователей подтвердили, что строматолитообразователи - это прокариоты, представленные чрезвычайно однообразными филаментами цианофитных, одноклеточными сфероидальными формами того же типа и значительно реже - бактериями. Морфологический консерватизм названных прокариот находится в совершенно очевидном противоречии с макроструктурным разнообразием строматолитовых построек и заставляет думать, что последние в большей мере имеют экологический, чем таксономический смысл. Все это должно резко ограничивать стратиграфическое значение строматолитов определенными фациями и крупными периодами (100-300 млн. лет) экологического выравнивания условий развития древних бассейнов.
В биологическом отношении рифей очень важен тем, что с ним, несомненно, связано развитие эвкариот. Датировать точно этот фундаментальный скачок в эволюционном процессе пока нельзя, по-видимому, он произошел не ранее начала рифея и не позднее рубежа между ранним и средним его этапами. Во всяком случае, в доломитах Бэк Спринг (шт. Калифорния, США) уже были обнаружены зеленые одноклеточные эвкариотные водоросли, возраст которых достигает 1,3 млрд лет [2109]. Одна из богатейших флор конца среднего рифея существовала в Сибири [1050]. В мироедихинской свите Туруханского района и в нижней части лахандинской серии Учуро-Майского района обнаружены разнообразные одноклеточные водоросли прекрасной сохранности, в том числе формы с резко обособленным внутренним телом, сходным с ядром, и с признаками различных стадий амитоза и клеточного деления, многочисленные цепочечные агрегаты "ядерных" клеток и массовые скопления водорослевых филаментов. Здесь же открыты формы, морфологически идентичные или близкие современным Phycomycetes, также содержащие темное внутреннее тело. Подлинной сенсацией стало в свое время открытие позднерифейской микрофлоры в формации Биттер Спрингс Центральной Австралии (возраст около 900 млн лет). Эта микрофлора стала важнейшим объектом интерпретации древнейших эвкариот, среди которых были идентифицированы разнообразные зеленые водоросли (такие, как близкие к Nucellosphaeridium - Caryosphaeroides, Clenobotrydion и другие), возможно, красные и пиррофитовые водоросли (Zosterosphaera), а также очень обильные осциллаторные прокариотные цианофиты [2232, 2233, 2234].
На Сибирской платформе в среднем и верхнем рифее в последние годы встречены окремненные строматолитовые породы, в которых в изобилии находят разнообразные микрофоссилии [690].
По-прежнему остро дискуссионной проблемой остается происхождение многоклеточных растений и многоклеточных животных. Но независимо от того, какую гипотезу происхождения многоклеточных организмов мы примем [их критический обзор см.: 411] и будем ли считать, что обе группы произошли независимо и одновременно или разделению этих царств предшествовал период симбиогенетического развития, три заключения подкреплены сейчас хорошо обоснованными фактами: 1) появление макроскопических органических форм не ранее позднего рифея (900-1000 млн лет); 2) более или менее близкое по времени (в пределах позднего рифея и особенно венда) появление бентосных Metaphyta с необызвествленным слоевищем и следов жизнедеятельности беспозвоночных, а позднее - самих отпечатков и остатков Metazoa; 3) достаточно высокий уровень содержания свободного кислорода в атмосфере того времени, так как только переход к кислородному дыханию мог стать важнейшим энергетическим импульсом в развитии всех жизнеобеспечивающих систем многоклеточных организмов. Этот скачок, который характеризовался переходом от брожения к окислению, известный под названием "эффекта Пастера", произошел не в начале кембрийского периода, как полагали еще совсем недавно [1330], а по меньшей мере на 400 млн лет раньше, т. е. на границе среднего и позднего рифея. Именно в позднем рифее, а возможно, и несколько ранее, произошли события, пока в значительной степени скрытые от нас, результатом которых явилась первая яркая вспышка и дальнейший подлинный расцвет вендской флоры и фауны.
В стратиграфической шкале венд может рассматриваться только в ранге особой геологической системы [921, 994, 2303], а не какого-либо подразделения рифейской или палеозойской группы, или кембрия, как предлагали А. и Ж. Термье [1046], выделив эдиакарский ярус (эдиакарий). Самостоятельность венда определяют: 1) планетарная устойчивость в последовательности основных геологических и палеонтологических особенностей, характеризующих предкембрийский период; 2) геохронологическая продолжительность вендского периода (около 100 млн лет в интервале 680±20-570±20 млн лет; 3) очевидная независимость всей вендо-эдиакарской биоты бесскелетных многоклеточных животных и растительных организмов от биот позднего рифея и кембрия, по отношению к которым первую нельзя считать даже переходной. В палеобиологическом и биостратиграфическом отношениях это принципиально важно для характеристики венда.
Палеонтологическая характеристика венда по своему разнообразию и уровню развития животных и растительных организмов отличается от характеристики рифея, синия и их аналогов, хотя на уровне так называемых "микробиот" и структурных биолитов (строматолитов и микрофитолитов), несомненно, существует преемственность. Самая ранняя кембрийская (томмотская) биота скелетных организмов несет в себе все типичные признаки фанерозойских биот, но по происхождению не имеет прямой связи с наиболее характерной для венда биотой бесскелетных многоклеточных животных.
Вендская биота сейчас представляется столь важной для понимания ранних этапов формирования основных филумов многоклеточных животных, что мы попытаемся более подробно охарактеризовать органический мир этого периода.
Рубеж рифея и венда был эпохой крупных площадных оледенений. Нам чрезвычайно мало известно об этой эпохе, но именно с ней приходится связывать последовавшую вскоре важнейшую эволюционную и экосистемную перестройку жизни в позднем докембрии. Вполне вероятно, что в эпоху лапландского оледенения резко понизился уровень Мирового океана и одновременно увеличилась соленость его вод в связи с тем, что значительная часть объема воды перешла в лед. С одной стороны, это могло вызвать резкую редукцию зоны шельфа, вплоть до полного исчезновения этой наиболее продуктивной зоны моря. Та же участь, вероятно, постигла и обширные материковые моря, столь характерные для позднего докембрия. Часть шельфа, еще затопленного водой, также могла оказаться непригодной для жизни организмов, будучи перекрытой ледником. Если в зоне шельфа до оледенения существовала обильная жизнь, то с сокращением его площади должна была резко повыситься конкуренция за место обитания и ресурсы питания. Вполне допустимо, что для многих групп фауны и флоры, заселявших мелководные зоны материковых морей и шельфа, лапландское оледенение могло стать катастрофическим. В этой ситуации мы скорее всего должны были бы наблюдать угнетение, стирание шельфовых сообществ и расцвет пелагических организмов, прежде всего фито- и зоопланктона. Однако, как известно, повышение солености обычно снижает продуктивность планктона, и мы вправе ожидать этого в течение лапландской гляциальной эпохи. Такой теоретически представляется предвендская эпоха.
Широчайшая эпиконтинентальная ранневендская трансгрессия, последовавшая за таянием ледников (следы деятельности этих ледников в виде тиллитов и тиллоидов залегают в основании венда на всех материках), привела к появлению на выровненных ледниками платформах гигантских мелководных морей. Вследствие беспрепятственного размыва берегов и увеличения привноса терригенного материала с континентов в условиях усилившейся денудации последних огромные площади оказались занятыми зоной литорали. Возникновение новых местообитаний и экологических ниш привело к появлению резко обновленного органического мира, типом которого и является вендо-эдиакарская биота бесскелетных многоклеточных. В этих совершенно новых ландшафтных и трофических условиях произошел, вероятно, самый важный для дальнейшего развития жизни на Земле таксономический и популяционный "взрыв". Роль инкубатора этой жизни, по-видимому, сыграла пелагиаль предвендского Мирового океана, а не "континентальные лужи", как иногда считают. Иначе трудно объяснить принципиальную однотипность вендской биоты на всех современных континентах.
Произошедший примерно через 100 млн лет следующий кембрийский "популяционный взрыв" скелетных беспозвоночных не был, таким образом, первой внезапной вспышкой метазоа, основные филетические стволы которых уже давно разошлись. Трудно установить прямую эволюционную связь известных групп фауны вендско-эдиакарской биоты бесскелетных метазоа со скелетными организмами кембрия, но высокий уровень дифференциации беспозвоночных венда, несомненно, сближает их с фанерозойским миром животных, несмотря на значительную специфичность и как бы завершенность биоты венда.
Если говорить о биоте вендского периода в целом, то в ней сохраняют свое место все названные ранее довендские группы микроорганизмов (а это таксоны царств, типов и классов). Совершенно исключительное значение приобретают разнообразные бесскелетные метазоа, следы жизнедеятельности бентосных беспозвоночных, водоросли, с неминерализованным слоевищем, ясной клеточной структурой и органами спороношения, существенно усложненные группы различного фитопланктона синезеленых, зеленых, красных и других водорослей (вплоть до гигантских сфер Chuariomorphida), нераспознаваемый пока микрозоопланктон, возможные актиномицеты, грибы, первые споровые растения амфибийного или континентального происхождения. В конце венда появились водоросли с обызвествленным слоевищем и первые метазоа с самыми ранними проявлениями склеротизации тканей на основе органических матриц и путем минерализации.
У нас пока нет явных свидетельств экологического характера, объясняющих отсутствие тканевой минерализации в докембрии. Многочисленные гипотезы, высказанные по этому поводу, не выдерживают критики [541, 1162]. К тому же мы не можем указать существенных различий в обстановках венда и кембрия: скорее, они были сходными. Биохимические механизмы формирования минерализованных скелетов, по-видимому, не эволюционировали или не проявлялись во время радиации мегаскопических метазоа в раннем и среднем венде. К тому же (а это надо иметь в виду) многие группы организмов, характерных для венда, не обладали скелетом в силу своей организации, так же как и сейчас не имеют скелета большое количество морских беспозвоночных - сифонофоры, актинии, аплакофоры, плоские черви, сипункулиды, аннелиды, оболочники и др.
Мы располагаем, однако, несколькими фактами, позволяющими утверждать, что в венде на уровне массового распространения вендо-эдиакарской фауны в некоторых группах животного царства биохимические механизмы склеротизации тканей уже проявлялись.
Об этом свидетельствуют прежде всего находки сабеллидитид и сколекодонтоморфных редкиний на редкинском уровне валдайской серии венда на Русской платформе [987, 988].
О наличии спикул (хотя мы и не знаем их вещественного состава) у некоторых вендских колониальных полипов можно судить по тонким узким отпечаткам, расположенным ориентированно на главном стебле, как, например, у известных эдиакарских арборей [1639]. О находках спикул в докембрийских толщах сообщалось много раз. Позже большинство этих находок было подвергнуто ревизии, но не считаться с этим материалом нельзя. Так, в группе Тиндир на Аляске обнаружены мелкие окаменелости, описанные как плоские черви класса турбеллярий [1266]. Повторное изучение этих образований склонило исследователей к мнению, что описанные фоссилии являются спикулами, напоминающими спикулы губок гексактиннелид [1443], а возраст вмещающих пород - кембрийский. Резко рельефные отпечатки некоторых вендских форм, в частности птеридинид, заставляют предполагать наличие у них достаточно жестких, вероятнее всего, органических скелетных элементов.
Вендская фауна СССР (особенно Русской платформы после открытия крупнейших местонахождений в Беломорском и Подольском регионах) сейчас является самой богатой в мире: она включает многие десятки родов и тысячи экземпляров. Эта фауна связана с наиболее полной стратиграфической последовательностью между рифеем и кембрием и сопровождается богатой и разнообразной ассоциацией других групп органического мира. На основании данного материала можно делать наиболее обоснованные общебиологические выводы, касающиеся специфики вендского этапа эволюции.
Для многочисленных животных венда характерны следующие первостепенно важные черты. Фауна венда представлена бесскелетными организмами, т. е. формами, не имеющими минерального скелета в самом широком смысле этих слов. Первые тубулярные формы с минеральным скелетом появились лишь в конце венда. Превосходная сохранность отпечатков бесскелетных может свидетельствовать о том, что в венде происходило прогрессивное уплотнение тканей, связанное с их совершенствованием и дифференциацией. Как продолжение этого процесса можно рассматривать активную склеротизацию тканей и массовое появление в кембрии скелетной фауны. Но не только уплотнение тканей, в том числе и покровных, способствовало фоссилизации отпечатков вендских метазоа. Судя по тому, что до сих пор нигде в мире не известно ни одного поврежденного остатка докембрийского бесскелетного животного, в этот период, по-видимому, не существовало активных хищников, а также организмов, питавшихся крупными пищевыми частицами (в том числе трупоедов). Значение биологических деструкторов было, вероятно, незначительным [299].
Кроме того, есть основания предполагать существование в венде сравнительно коротких пищевых цепей. И наконец, немаловажное обстоятельство - относительно небольшой удельный вес инфауны в бентических сообществах вендских морей, а отсюда и незначительная степень биологической переработки осадка. Все перечисленные обстоятельства способствовали тому, что остатки вендских бесскелетных могли захороняться в больших количествах, а их отпечатки оставались ненарушенными.
Одна из характернейших черт вендской фауны - проявления гигантизма в разных крупных таксонах. В вендских отложениях нередки отпечатки медузоидных диаметром 30-40 см, а длина некоторых перистовидных колоний полипов подчас достигает более 1 м. Такие размеры свойственны наиболее примитивным группам, в частности кишечнополостным и некоторым плоским червям. Гигантизм вендских представителей этих групп выступает особенно ярко и неожиданно на фоне шельфовой биоты скелетных форм начала кембрия (томмотский век), представленной разнообразными, но преимущественно очень мелкими формами, которые не обнаруживают прямого родства с крупными бесскелетными венда. Только позднее, в кембрии же, появляются свои, редко встречающиеся гиганты, но они относятся уже к иным типам и классам. Поэтому нередко можно слышать суждения, что вендо-эдиакарская фауна не имеет потомков. Это представление вряд ли верно, хотя основано на видимой, фактической стороне палеонтологической летописи. Если судить о преемственности животного мира рассматриваемых периодов на уровне типов, то сомнений в ее существовании быть не может: и в венде, и в кембрии известны кишечнополостные, аннелиды, членистоногие, возможные погонофоры, а также моллюски (на их присутствие в венде указывают разнообразные следы ползания и выедания осадков).
Не исключено, что обширная группа археат найдет своих предков среди некоторых "медузоидных" венда, которые имеют плоскоконическую форму, монаксонную гетерополярную симметрию неопределенно большого порядка и демонстрируют все признаки седентарности. Сравнение вендских и кембрийских форм в некоторых случаях может быть осложнено тем, что в вендском материале присутствуют отпечатки мягких тканей, а в кембрийском - только остатки скелетов. В венде, вероятно, одновременно с гигантскими формами (гигантизм некоторых из них, возможно, отражает наиболее поздние фазы эволюции этих групп метазоа) существовали мелкие бесскелетные организмы, давшие в томмотском веке вспышку скелетных беспозвоночных фанерозоя. Микропалеонтологи, исследующие вендские биоты, уже обнаружили в своих преператах некоторые элементы зоопланктона, но в целом эта область пока остается нетронутой, хотя она и чрезвычайно интересна.
Вендская фауна в целом характеризуется высокой морфологической дифференциацией, отражающей уже сложившиеся таксоны очень высокого ранга, до типов включительно. Видовое разнообразие на этом уровне дифференциации незначительно. Надежные данные о дивергенции таксонов высокого ранга в венде пока отсутствуют. Все типы и классы вендской фауны появляются как бы внезапно. Для наиболее вероятного объяснения этого явления приходится допустить, что причина его состоит не в замаскированной длительности путей расхождения филумов, а в необычайно быстром формировании всех главнейших типов беспозвоночных в геологически очень короткое время. На ранних стадиях радиации метазоа при относительно низком уровне их организации каждый новый признак может иметь очень высокий таксономический вес и в то же время не иметь адекватного функционального или физиологического значения для самих организмов. Теоретически более вероятен процесс быстрой реализации всевозможных вариантов архитектоники беспозвоночных в общих чертах после или в процессе возникновения многоклеточности. Это предположение косвенно подтверждается следующим наблюдением: 60 из 90 классов морских многоклеточных животных появились в вендо-кембрийском интервале, а от перми до наших дней - только пять [2239, 2274-2277].
Таким образом, предположение, что метазоа имеют очень длительную историю в докембрии, сейчас кажется менее вероятным. Природа этого "макротаксономического" скачка гипотетична, но скорее всего экологически экзогенная. Для докембрийской эволюции сегодня наиболее популярно представление о биоэнергетических критических рубежах, связанных с общим ходом оксигенизации атмосферы и гидросферы, хотя и этот фактор не автономен в системе абиотических и биологических параметров среды жизни. Само собой разумеется, что необходимо продолжение настойчивых поисков возможных остатков метазоа и в довендском протерозое.
Вендские бесскелетные метазоа характеризуются всеми главными чертами архитектоники позднейших беспозвоночных, что, вероятно, связано с быстро устоявшимися приспособлениями к условиям среды обитания, с формами движения (дно, пелагиаль), с типом питания, роста и размножения. В вендских морях были обильно представлены сидячий и подвижный бентос, нектонные, планктонные и даже плейстонные организмы. В то же время во всех известных местонахождениях на Земле рассматриваемая фауна экологически принципиально однотипна. Она включает сравнительно однообразный набор жизненных форм, представленных десятками (вероятно, до сотни) родов. В этом можно видеть свидетельство однообразия экологических условий вендского шельфа и отсутствия сколько-нибудь существенных экологических барьеров, стертых огромной трансгрессией. Устойчивость в стратиграфической последовательности и однотипность вендской биоты бесскелетных метазоа исключают возможность асинхронности в ее возрастном положении в целом, но порождают другую острейшую проблему, связанную с выявлением характера и скорости эволюции в венде различных групп многоклеточных животных. Пока данные на этот счет чрезвычайно скудны. Более того, иногда можно слышать суждения, что вендская фауна нестратиграфична, так как заметно не эволюционировала на протяжении почти сотни миллионов лет. Быстрое развитие знаний о ней, достигнутое в последние годы прежде всего в нашей стране, позволяет надеяться, что в ближайшее время будут сделаны первые шаги в разработке данной проблемы.
Анализ архитектоники вендских метазоа [1097-1099] показывает, что наиболее ранняя радиация многоклеточных шла по пути реализации симметрийного разнообразия. В процессе ее появились разнообразные планы строения. Некоторые из них были унаследованы в ходе дальнейшей эволюции метазоа, став характерными признаками таксонов высокого ранга в фанерозое. Уже давно замечена определенная количественная асимметрия в распределении представителей подчиненных таксонов в объединяющих их более крупных систематических единицах. В царстве животных венда несомненная доминанта принадлежала кишечнополостным - медузоидам пелагиали и разнообразным одиночным и колониальным полипам. Венд можно было бы охарактеризовать как время бурного расцвета кишечнополостных. Не исключено, однако, что мы несколько преувеличиваем роль этого типа в вендской биоте: кишечнополостные являются в общем наиболее крупными организмами этого периода, среди них имеется большое количество разнообразных полипов, которые в целом имели больше шансов сохраниться в ископаемом состоянии. Все это приводит к тому, что вендские кишечнополостные чаще сохраняются и чаще попадаются на глаза палеонтологу (отсюда и большая численность и большое видовое разнообразие): представители этого типа составляют 67% от 1500 образцов бесскелетных метазоа, собранных в районе Флиндерс Рейндж близ Эдиакары в Южной Австралии [247]. В беломорской же биоте венда кишечнополостные составляют около 70% общего числа описанных видов [1094].
Резкое преобладание Radialia в ископаемых ассоциациях вендских метазоа может рассматриваться как признак наибольшей древности этой группы. Широкое симметрийное разнообразие вендских кишечнополостных отражает более ранний этап начальной радиации данного типа. Самой архаичной является монаксонная гетерополярная симметрия бесконечно большого порядка, обычно сопряженная с концентрическим планом строения, что в целом характерно для многочисленных примитивных форм вендских кишечнополостных (класс Cyclozoa). Более высокой организацией отличаются Radialia с симметрией неопределенного порядка (класс Inordozoa), которая представляется необходимым этапом в эволюции многих линий кишечнополостных на пути к симметрии определенного порядка, типичной для книдарий фанерозоя. Кишечнополостные венда со стабильной симметрией представлены в основном трехлучевыми (класс Trilobozoa) и четырехлучевыми (класс Scyphozoa) формами, которые отвечают высшему уровню организации. Анализ архитектоники вендских кишечнополостных допускает предположение об эволюции Radialia от форм с симметрией бесконечно большого порядка через формы с неопределенно многолучевой симметрией к формам с определенным порядком симметрии и от форм с концентрической организацией к формам с преимущественно радиально-лучистым планом строения [1098, 1099, 1577].
Для Bilateria, составляющих меньшинство среди ископаемых вендских метазоа, характерно преобладание сегментированных форм на фоне большого разнообразия планов строения, которое отражает широкий диапазон типов организации и уровней продвинутости отдельных ветвей этой группы. Резкое преобладание сегментированных форм среди вендских билатерий допускает предположение, что во многих ветвях метазоа процессы становления билатеральной симметрии и метамерии были сопряженными, однако они не всегда вели к целоматам. Симметрия скользящего отражения, характерная для наиболее примитивных билатерий венда (тип Proarticulata) и нетипичная для позднейших билатерально симметричных метазоа, может указывать на вероятность очень раннего происхождения метамерных форм от радиально-симметричных метазоа с осью вращения неопределенного порядка [1097, 1098]. Неупорядоченная метамерия с плоскостью симметрии скользящего отражения могла быть важным этапом в эволюции некоторых групп метазоа на пути к настоящим Articulata.
В конце венда произошло массовое вымирание многих групп метазоа, прежде всего таксонов, составляющих наиболее характерные элементы вендо-эдиакарской фауны: специфические высокоранговые группы, которые нередко именуют как "опыты Природы" [1665].
Завершая характеристику вендского животного мира, следует отметить исключительное значение жизнедеятельности беспозвоночных, изучение которых дает специфическую информацию, которую подчас нельзя получить при изучении иных окаменелостей. В истории прогрессивной колонизации морского дна вендский период был, несомненно, одним из важнейших этапов, хотя в это время бентосные организмы осваивали в основном мелководные зоны бассейнов. В типичных случаях вендские ихноценозы отличаются относительно невысокой степенью биологической переработки осадка (по сравнению с ихноценозами кембрия) и неглубоким зарыванием организмов, субгоризонтальностью следов передвижения и питания, незначительным количеством норок-жилищ, следов покоя и следов перемещения с участием придатков [1096]. Ископаемые следы жизнедеятельности из вендских отложений свидетельствуют о высокой организации вагильного бентоса. В приповерхностной зоне осадка эти организмы передвигались с помощью перистальтических изменений формы всего тела, как, например, современные аннелиды, сипункулиды и другие червеобразные. Среди них были и сортирующие, и несортирующие детритофаги, пропускавшие через пищеварительный тракт осадок, богатый детритом и микроорганизмами. В пользу этого могут свидетельствовать следы Planolites и Neonereites. Другие организмы, судя по таким следам, как Nenoxites, Bilinichnus, Aalfchnites, перемещались по поверхности осадка при помощи волн перистальтики, проходящих по вентральной части тела или по подошве ноги (современные турбеллярии, хитоны и гастроподы). Перистальтический способ перемещения многоклеточных, по-видимому, является одним из наиболее примитивных и древних.
В венде получают распространение следы с повторяющимися элементами симметрии, образованные представителями вагильного бентоса в результате повторения однотипных движений в процессе питания [1091, 1095]. Эти следы указывают на появление определенных наследуемых поведенческих программ, направленных, по-видимому, на рациональное использование ресурсов питания при оптимальных затратах энергии. В дальнейшей истории животного мира с помощью ископаемых следов можно наблюдать поразительную картину эволюции поведения разнообразных групп морского бентоса на фоне прогрессирующей колонизации все более глубоководных зон дна и толщи осадка. Эволюционная сторона этологии, которая совсем недавно начала развиваться на основе изучения ископаемых следов жизнедеятельности, по-видимому, еще долго будет оставаться привилегией палеоихнологии.
К концу венда становится заметным усиление активности вагильного бентоса, но наиболее резкое ускорение процесса-колонизации морского дна, особенно в зонах мелководья, наблюдается в самом начале кембрия, или даже чуть раньше. На Русской платформе в ровенском горизонте балтийской серии появляется богатая и разнообразная ассоциация ископаемых следов - Treptichnus, Teichnus, Phycodes, Cyzolithes и др. [800, 1095, 1096], многие из которых продолжают существовать и в более поздние геологические эпохи. В целом ихноценозы этого уровня характеризуются высокой степенью биологической переработки осадка, значительным разнообразием и морфологической сложностью, а нередко и крупными размерами. Появляются новые формы с повторяющимися элементами симметрии, причем не только горизонтальные, но и вертикально ориентированные. Биота ровенского времени характеризуется обильными сабеллидитидами, значительным количеством одиночных полипов (Bergaueria), которые богаты фитопланктоном, близким к вендскому, а также многоклеточными водорослями (Tyrasotaenia). Вероятно, к этому времени относится появление первых тубулярных метазоа с минерализованной трубкой (немакит-далдыкский горизонт Сибири). Наиболее значительные обновления органического мира происходят начиная с лонтоваского-люкатиского горизонтов на Русской платформе и с сунагинского горизонта и его аналогов в Сибири. Массовое появление скелетной фауны, обусловленное вспышкой минерализации скелетных элементов в самых разных стволах органического мира Земли, послужило началом, точкой отсчета фанерозойского эона и фанерозойской эволюции биосферы (рис. 22), которая до недавних пор была единственным объектом исследований классической палеонтологии.
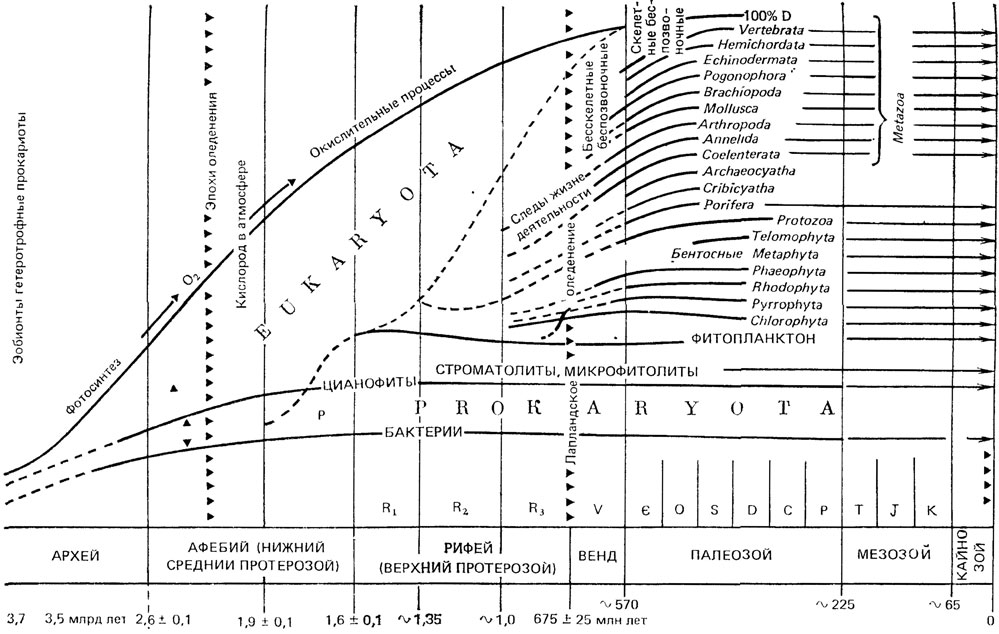
Рис. 22. Схема развития органического мира. По Б. С. Соколову, с незначительными изменениями
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'