
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Морфологическая эволюция онтогенеза
Прогрессивные дефинитивные изменения. Проследим теперь морфологические проявления стабилизационного процесса. Некоторые из них самоочевидны. Если новая элементарная адаптация всегда возникает как ответ на определенный внешний фактор, а позднее (в филогенезе) автономизируется от него в своем развитии, появляясь уже на предфункциональной стадии, то ясно, что речь идет об ускорении (акселерации) развития или, вообще говоря, о гетерохронии.
Другие закономерности удобнее пояснить с помощью схемы (рис. 28). Допустим, что морфогенез органа проходит через стадии А, В к взрослому состоянию С. Отбор на расширение адаптивных возможностей приводит к тому, что в последующих циклах развития взрослая стадия при воздействии фактора среды f реагирует новой приспособительной дифференцировкой D(DI). Тем самым стадия С рекапитулируется здесь в почти ненарушенном виде (С1); изменилась лишь ее реактивная способность в определенных условиях (f). По мере совершенствования дифференцировки D в филогенезе (DII) ее развитие стабилизируется и больше не нуждается во внешней индукции. Роль формативной основы стадии DII целиком берет на себя предыдущая стадия, которая снова должна видоизмениться (С2), чтобы удовлетворить этому новому требованию. Рекапитуляция исходного взрослого состояния С тем самым искажается уже заметнее. Дальнейшая перестройка развития в ходе его интеграции с другими морфогенезами может затронуть уже не только непосредственную базу структуры D (достигшей уровня DIII или DIV), но и более глубокие основы организации, благодаря чему появляются стадии C3 и B1, а затем и C4, B2, A1. Все эти изменения могут иметь на первых порах чисто физиологический характер, но по мере их углубления они получают и морфологическое выражение.
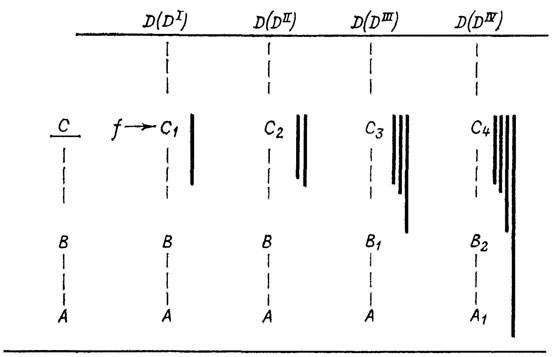
Рис. 28. Перестройка морфогенеза нового признака в процессе его стабилизации. По М. А. Шишкину. A, A1; B, B1; B2; C, C1-C4 - стадии онтогенетического развития органа; D(DI)-D(DIV) - этапы эволюционной стабилизации нового признака; f - внешний формативный фактор; сплошные вертикальные линии - накопление стабилизационных изменений
Из этой картины ясно, что эволюция поздних стадий не является ни их наращиванием (анаболией), ни замещением (субституцией) в чистом виде, а результатом непрерывного превращения первого процесса во второй. Данный пример, по существу, демонстрирует этапы эволюции онтогенеза при создании единственной надставки D. Поскольку новый признак возникает как формообразовательная реакция прежней взрослой стадии, то он вначале является надстройкой над ней. Но параллельно созданию признака стабилизируется и способ его развития, что ведет к нарастающему искажению рекапитуляции прежней взрослой стадии.
Таким образом, эволюция путем изменения поздних стадий в равной степени является причиной как появления палингенетических рекапитуляций, так и их последующего искажения. Или, иначе говоря, нарушение биогенетического закона обусловлено в конечном счете той же самой причиной, что и его выполнение. Процесс стабилизационной перестройки, осуществляющий это нарушение, распространяется с затуханием в последовательных онтогенезах от поздней (взрослой) стадии к ранним (рис. 28). Чем дальше он заходит, тем глубже новый цикл развития отличается от исходного. Так, если онтогенез A-DII уклоняется от предкового А-С, начиная со стадии В, то в следующем филетическом звене A-DIII момент расхождения с A-C отодвигается к стадии A, а в дальнейшем (A1-DIV) - к еще более ранним стадиям.
Эти закономерности открывают нам смысл закона К. М. Бэра, говорящего об углублении онтогенетической дивергенции между родственными формами по мере отдаления их родства. На первый взгляд при таких соотношениях между онтогенезами предка и потомка второй из них никогда не может быть продолжением первого. Поэтому неудивительно, что, несмотря на все попытки согласовать закон Бэра с законом Геккеля [930, 1896], они чаще всего считаются "фундаментально несовместимыми" [1661, 1767]. Но при этом не замечают, что эти два обобщения имеют разные области применения, определяемые тем, насколько длительную эволюцию претерпел данный онтогенез по отношению к сравниваемому исходному. Если бы этой эволюции не было, то мы наблюдали бы в развитии любого сколь угодно отдаленного потомка лишь приращение к анцестральному циклу, а не отклонение от него. Закон Бэра мог выполняться бы только для филетически дивергентных форм, причем общий ранний отрезок их развития состоял бы из одних палингенезов (рекапитуляции взрослых общих предков)*. И лишь стабилизационные процессы, непрерывно совершенствующие и упрощающие в поколениях путь реализации ранее приобретенных новшеств, приводят к тому, что онтогенетическая запись каждого такого приобретения постепенно преобразуется из "продлевающей" в "замещающую". Другими словами, чем более далекие звенья филогенетической цепи мы сравниваем, тем меньше их онтогенетическая разница подчиняется биогенетическому закону и тем больше - закону Бэра.
* (Последние могли бы затемняться только эмбриональными адаптациями (ценогенезами).)
Это дает ответ на одно из главных упоминавшихся возражений против биогенетического закона, указывающее, что его классические примеры в действительности относятся к проявлениям зародышевого сходства, которые имеют отношение к особенностям взрослой стадии предка лишь в той мере, в какой они сохранялись у него до конца онтогенеза [1317, 1623]. Например, присутствие у эмбрионов высших позвоночных жаберных щелей повторяет у них в первую очередь эмбриональное, а не взрослое состояние рыбьих предков, и т. д. Неслучайно такие возражения исходят прежде всего от зоологов и эмбриологов, поскольку в качестве моделей предка и потомка им приходится использовать почти исключительно современные формы, представляющие собой продукты дивергентной эволюции с глубоко разошедшимися онтогенезами. Напротив, палеонтологу часто удается наблюдать очень близкие звенья филогенетической цепи и убеждаться, что в этом случае анцестральная взрослая стадия может отражаться в позднем онтогенезе потомка с максимальной полнотой [887]. Причина этого понятна и заключается в том, что морфогенез исторически недавнего приобретения еще мало затронут стабилизационными преобразованиями и в основном сохраняет свой первоначальный вид надстройки над предыдущим взрослым состоянием. Именно этими возможностями, предоставляемыми ископаемым материалом, и объясняется тот странный для многих факт, что, несмотря на многолетнюю уничтожающую критику биогенетического закона, палеонтология по-прежнему остается его оплотом*.
* (Иногда даже считают, что теория рекапитуляции имела больше воздействия на палеонтологическую мысль, чем любая другая концепция, за исключением самой идеи органической эволюции [1535].)
Рассмотренные нами закономерности стабилизационных изменений по отдельности не составляют чего-то ранее неизвестного. Мысль о преобразовании надставок развития в субституцию высказывалась С. Г. Крыжановским [540], подчеркивавшим, что прибавление новой стадии в морфологическом смысле означает вытеснение ею предыдущей во временном отношении. Обусловленность наиболее характерных проявлений перестройки онтогенеза (акселерации и спрямления пути развития) состоянием взрослой стадии понималась многими исследователями независимо от того, связывали ли они эти изменения с появлением новых надставок, "впрессовывающих" вглубь прежние стадии (Э. Д. Коп, А. Хайетт, А. Вейсман и др.), или же с самим фактом длительного исторического существования данной взрослой организации (Ф. Мюллер, Ф. Бальфур). В последнем случае интуитивное объяснение, по существу, было очень близко к пониманию процесса стабилизации, так как указывалось, что природа стремится достигнуть желательного конца кратчайшим путем, вызывая укорочение первоначальной онтогенетической записи [1299]. В целом, однако, для гетерохронии (изменения темпов развития) и выпадения стадий выдвигались самые различные объяснительные причины - чисто адаптивные [309, 1775, 2506], механические [2470] или же имеющие особую природу [1457].
Но если указанные явления, с одной стороны, обусловлены эволюцией взрослой стадии, а с другой - сами обусловливают нарушение рекапитуляции, то, очевидно, между двумя последними процессами должна существовать прямая связь. Первым это понял эмбриолог Э. Менерт [1945], установивший, что скорость развития отдельных органов в онтогенезе зависит от степени их филетического прогресса. Поскольку эта зависимость при неравномерной эволюции органов ведет к нарушению прежних целостных онтогенетических стадий, то изменения, связанные с гетерохрониями, согласно Э. Менерту, не могут считаться фальсификациями исторического развития, а составляют его закономерный результат. В палеонтологии подобные взгляды были почти одновременно высказаны Дж. Смитом [2295].
Наиболее далеко в вопросе о влиянии изменений взрослой стадии на ход онтогенеза продвинулся А. Н. Северцов [927, 928] в раннем варианте своей теории филэмбриогенезов. Анализируя факторы нарушения рекапитуляции, не связанные с провизорными приспособлениями, он выделил в них две группы - первичные и вторичные филэмбриогенезы. Под первыми подразумевались ранние эмбриональные уклонения, преобразующие ход дальнейшего онтогенеза, под вторыми - искажения онтогенетической записи, обусловленные эволюцией конечных надставок, т. е. те самые изменения, которые интерпретированы нами как результат стабилизационной перестройки.
Основываясь на правиле Э. Менерта, А. Н. Северцов показал, что следствием поздних изменений органов является искажение рекапитуляции даже в пределах отдельных органогенезов. Сдвигаясь на другие стадии, закладки эволюирующих органов меняются и сами по различным параметрам (размеры, гистологическая структура, форма, соотношения с другими частями). Распространение этих изменений в направлении ранних стадий (в филогенезе) было продемонстрировано А. Н. Северцовым как для ускорения, так и для замедления развития. Был сделан также вывод о наибольшей полноте рекапитуляции в самых поздних стадиях, еще мало задетых вторичными изменениями. Наконец, было показано, что ускорение (эмбрионализация) развития взрослых черт связано не с позднейшей надстройкой онтогенеза (как гласит обычная интерпретация биогенетического закона), а с длительностью их собственного исторического существования, как полагал еще Ф. Мюллер. Отсюда следовало (хотя и не было четко сформулировано), что эмбрионализация сама по себе есть признак прогрессивной эволюции органов.
Таким образом, А. Н. Северцову уже были ясны основные морфологические закономерности, рассмотренные нами. Отсутствовало лишь представление о факторе эволюции, приводящем их в действие, - стабилизирующем отборе, ведущем к максимально надежному воспроизведению адаптивного фенотипа в онтогенезе. Вместо этого выдвигались традиционные объяснения, предполагающие компенсацию удлинения онтогенеза и узко адаптивное значение сдвигания стадий для взрослой формы. Поскольку изложенные представления целиком опирались на эмпирическое правило Э. Менерта, понимаемое как принцип "сквозного" изменения темпов развития, то, как только обнаружились случаи, где это правило выполняется не столь жестко (время закладки органа у двух форм может оказаться одинаковым, несмотря на различия в степени его филетического развития), поколебалась и вся основа этих взглядов.
В результате А. Н. Северцов [926, 930] отказался от понятия о вторичных филэмбриогенезах. В его схеме, правда, появились "вторичные архаллаксисы", означающие частный случай того же явления, но они уже не рассматривались как закономерный результат эволюции поздних стадий. В такой же частный модус превратились и гетерохронии. Вместо перестройки, вызываемой надставками, филэмбриогенезами стали называться сами надставки (модус анаболии). Единый процесс распался на изолированные явления. В итоге из двух логически допускавшихся способов перестройки онтогенеза недоказанный путь (ранние изменения) остался в теории А. Н. Северцова, а бесспорный выпал. Теория превратилась в модификацию биогенетического закона, в которой, как и у Э. Геккеля, нарушение рекапитуляции не обусловлено эволюцией взрослой стадии. Разница лишь в том, что в геккелевском варианте все нарушения закона названы ценогенезами, а в северцовском к ним добавлены архаллаксисы и девиации*. Эта эволюция взглядов А. Н. Северцова лишний раз показывает, насколько судьба правильных эмпирических обобщений может зависеть от их теоретического обоснования.
* (Насколько нам известно, никто, за исключением отчасти В. Е. Руженцева [887], не обратил внимания на это радикальное изменение содержания теории филэмбриогенезов.)
Регрессивные дефинитивные изменения. Стабилизационные изменения, сопровождающие формирование новых адаптации, связаны с одновременным распадом старой адаптивной нормы. Оба эти явления составляют две неразрывных стороны эволюционного процесса. В любой морфологической структуре прогрессивное развитие одних черт сопровождается деградацией других (например, превращение плавника кистеперой рыбы в пятипалую конечность - это одновременно утрата кожных плавниковых лучей, развитие копыта лошади - это также редукция боковых пальцев и т. д.). Каждый элементарный шаг в этих изменениях имеет одну и ту же основу - отбор на устойчивость преобразованной нормы, ведущий к перестройке ее генотипа как целого, т. е. к изменению пространства индивидуальных геномных вариаций, допускающих ее надежное осуществление ([1180]; см. 7.3).
По отношению к любому признаку, потерявшему свое значение и не поддерживаемому более отбором, это изменение пространства вариаций выглядит как накопление беспорядочных генетических изменений, все более расшатывающих его развитие. Оно выражается в постепенном сдвигании различных морфогенетических параметров к тем порогам, за которыми уже невозможна регуляция нормального развития [1196, рис. 11], в результате чего последнее становится все более изменчивым и неполным. Вначале такие сдвиги задевают лишь позднее развитие; но по мере их накопления у потомков они начинают нарушать и более эффективно регулируемые ранние стадии.
Таким образом, при разрушении формообразовательного процесса обнаруживаются те же характерные черты стабилизационной перестройки, что и при его созидании. В обоих случаях изменения распространяются с затуханием в сторону все более ранних стадий и постепенно автономизируются от влияния факторов среды. Если сначала прежний ход развития преимущественно лишь теряет устойчивость, осуществляясь во все более узком интервале внешних условий, то в последующих поколениях он нарушается уже независимо от их колебаний. И хотя высокая изменчивость рудиментарных органов всегда сохраняется [310, 2141], она приобретает все более неупорядоченный характер. Среди примеров незавершенной рудиментации можно назвать развитие глаз и пигментации кожи у пещерной амфибии Proteus, где утраченная норма все еще в большей мере восстанавливается при выращивании личинок на свету. Напротив, стойкая утрата способности к метаморфозу, характерная в той или иной степени для многих хвостатых амфибий, есть проявление более глубоко стабилизированного недоразвития.
Путь углубления редукционного процесса хорошо виден из сопоставления типов рудиментации конечностей у современных рептилий, изученных А. Н. Северцовым [931]. Сравнение развития геккона (Ascalobotes) и змеевидных ящериц (Seps. Ophisaurus) позволяет считать, что в филогенезе редукция начинается с конечных стадий нормального морфогенеза, а затем захватывает все более ранние стадии, причем поздние к этому времени могут уже совсем выпадать. Так, при относительно меньшей степени редукции (Seps) не возникают фаланги и метаподий 1-го пальца, а также концевые фаланги остальных пальцев (нормально возникающие последними), тогда как проксимальные (рано развивающиеся) элементы закладываются уменьшенными. Дальнейшая рудиментация (Seps. Ophisaurus) ведет к исчезновению почти всех элементов кисти и уже сильному уменьшению зачатка zeugopodium. При этом у обеих форм процесс редукции уже настолько сокращен и ускорен, что целиком осуществляется эмбрионально*.
* (А. Н. Северцов [931] называет рудиментацию отрицательным архаллаксисом, поскольку весь процесс здесь выглядит эмбриональным, и в онтогенезе зачатки дегенерирующих органов сразу возникают уменьшенными. Однако вторичность обоих явлений ясно вытекает из закономерностей, вскрытых в этом случае самим же Северцовым. Им показано, что зачаток, уменьшенный как будто бы "сразу", на самом деле становится таковым в филогенезе постепенно ("делается все меньше и меньше" [931, с. 514]) и что рудиментация в ходе эволюции распространяется с поздних стадий на ранние. Если на основании этих двух положений экстраполировать к начальному этапу рудиментации, то неизбежен вывод, что ранние зачатки вообще ею не затрагивались, и изменения проявлялись лишь на самых поздних (постэмбриональных)) стадиях. Таким образом, не орган уменьшился вследствие уменьшения его зачатка, а наоборот, зачаток стал маленьким вследствие филогенетического углубления редукции органа.)
Наиболее характерные проявления недоразвития - выпадение анцестральных конечных стадий и замедление предшествующих преобразований, ведущие к переходу прежних эмбриональных или ювенильных черт во взрослое состояние - описаны под множеством названий; ретардация (Э. Д. Коп, А. Хайетт), брадигенез (А. Грабау), неотения (Ю. Колльман), эпистаз (О. Иекель), фетализация (Л. Больк), педоморфоз (У. Гарстанг), аббревиация (В. Франц), отрицательная анаболия (А. Н. Северцов), протерогенез (О. Шиндевольф) и т. д. На первый взгляд это - процесс, противоположный прогрессивному развитию, но, как мы видели, его эволюция подчиняется тем же самым закономерностям. Он точно так же начинается всегда с изменения (в данном случае - распада) дефинитивного состояния и по мере его усиления распространяется у потомков на все более ранние фазы развития. Стирание рекапитуляции полно развитого состояния органа происходит при его регрессе точно таким же путем, что и в случае его дальнейшего усложнения или специализации. Другими словами, нет принципиальной разницы, затемняется ли в онтогенезе потомков стадия пятипалой когтистой лапы благодаря тотальному недоразвитию конечности (угревидные хвостатые амфибии и ящерицы) или же вследствие ее преобразования в крыло птицы.
Непосредственные адаптивные изменения промежуточных стадий. Все сказанное об эволюции взрослой стадии в равной степени приложимо и к любым другим стадиям. Каждая фаза онтогенеза может изменяться путем отбора ее регулируемых вариаций сколь угодно далеко при условии, что это не нарушает нормального течения последующего развития. Эти регулируемые, т. е. сходящие затем на нет провизорные приспособления, начинаются, подобно дефинитивным, с конечных (по времени) изменений той стадии, на которую направлен отбор; затем их механизм стабилизируется, захватывая и предыдущий отрезок развития. Сравнительно-эмбриологические доказательства поздних изменений провизорных стадий приведены, например, для яиц рыб и личинок амфибий [540, 932]. Стабилизационное ускорение таких изменений прослежено в развитии рисунка окраски у гусениц бражников, в котором более простые предковые типы рекапитулируются и сдвигаются на ранние стадии [2470].
Другой пример - развитие головастиков бесхвостых амфибий. Среди них существуют четыре типа строения жаберных камер, причем только в одном из них имеется парное spiraculum. (рис. 29, А1-Г1). Этот тип почти наверняка является исходным для остальных, которые, очевидно, когда-то возникли у вполне "взрослых" головастиков путем слияния обоих отверстий или зарастания одного из них. Парные отверстия действительно рекапитулируются во всех типах и далее преобразуются обоими указанными путями. Однако у современных форм это происходит на такой ранней стадии, когда опекулярные складки (стенки камер) только начинают развиваться, а самих внутренних жабр, характерных для "взрослого" головастика, еще нет (рис. 29, А-Г ср. [2319]). Очевидно, что ход первично позднего преобразования здесь давно изменен и ускорен в процессе его стабилизации. В основе последней лежит перестройка морфогенетических механизмов, следы которой иногда вполне явственны. Так, у наиболее специализированных головастиков передняя лапка развивается в жаберной камере и выходит наружу, прорывая operculum. Вероятно, вначале этот прорыв осуществлялся за счет механического давления лапки; но у современных форм перфорация operculum - вполне автономный процесс, вызываемый продуктами распада внутренних жабр [2014].
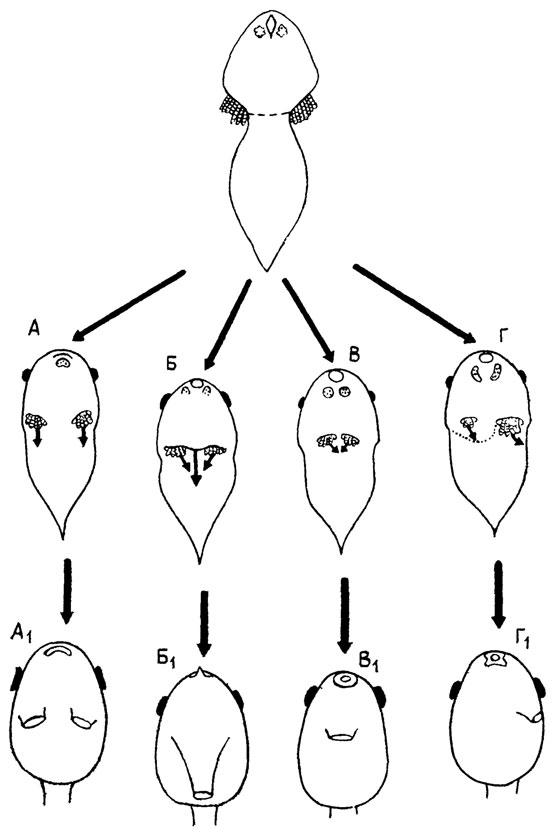
Рис. 29. Типы онтогенетического развития spiraculum у личинок бесхвостых амфибий. По П. Х. Старетт. А, Б, В, Г - ранние стадии, показывающие рекапитуляцию парного spiraculum; А1, Б1, В1, Г1 - стадии 'взрослых' головастиков (А-А1 - Pipidae и Rhinophrynidae; Б-Б1 - Microhylidae; В-В1 - Ascaphidae и Discoglossidae; Г-Г1, - остальные бесхвостые)
Таким образом, непосредственная адаптивная эволюция любой стадии онтогенеза протекает по одним и тем же законам, т. е. начинается с ее конечной фазы и затем, по мере стабилизации достигнутого изменения, распространяется вглубь в последующих поколениях. Тем самым исторические преобразования любой отдельной стадии сами по себе могут влиять лишь на предыдущий, но не на последующий ход онтогенеза. Эта значительная самостоятельность эволюции стадий приводит к тому, что чем больше нарастают изменения любой из них, тем более глубокой должна быть перестройка при переходе к следующему отрезку развития, имеющему собственную адаптивную эволюцию. Именно в этом состоит причина резких превращений, свойственных развитию с выраженным метаморфозом, отражающему перестройку между существенно различными возрастными адаптивными типами (у ракообразных, насекомых с полным превращением, бесхвостых амфибий и т. д.). Благодаря такому механизму преобразований онтогенеза его течение характеризуется не просто уменьшением изменчивости к концу, как подчеркивал К. М. Бэр, а чередованием стабильных, адаптивно значимых фаз, или "установок" [415], и более вариабельных отрезков, подготавливающих эти фазы. К числу первых относятся, например, различные специфические личиночные типы. В развитии черепа хвостатых амфибий такими адаптивно важными "установками" являются моменты прорыва рта и перехода к активному питанию, когда основные морфологические изменения, в отличие от предшествующих стадий, оказываются строго скоординированными [566]. Общим итогом развития всегда остается стабильное осуществление взрослой адаптивной нормы. Иными словами, главный принцип эволюции онтогенеза состоит в том, чтобы обеспечить это осуществление вне зависимости от самостоятельных адаптивных изменений его промежуточных стадий.
Провизорные адаптивные изменения вместе с их стабилизационными последствиями составляют то, что может быть названо ценогенезами. Это понятие имеет разные толкования вследствие его двойственности у Э. Геккеля [1688], который отождествил в нем: а) любые нарушения палингенезов и б) приспособления ранних стадий. Для авторов, отрицающих палингенезы, ценогенез как сопряженное понятие вообще не имеет содержания [540, 1623]. При использовании только второго из названных критериев [414, 928] оно охватывает и типичные палингенезы (например, жабры у личинок амфибий); если же исходить из первого [1945], то ценогенезом окажется любое стабилизационное изменение типа гетерохронии, несводимость которых к собственно провизорным адаптациям давно отмечена [926, 565, 928, 1317]. Неопределенность употребления термина иногда заходит так далеко, что им обозначают просто любой новый признак [2078]. Введение понятия о стабилизационной перестройке впервые позволяет внести ясность в этот вопрос. Дело в том, что ценогенез в представлениях Геккеля был прежде всего равнозначен изменениям, не связанным с эволюцией взрослого организма. Но на самом деле часть нарушений рекапитуляции обусловлена именно последствиями этой эволюции (стабилизацией дефинитивных изменений). Тогда все, что остается за их вычетом, и должно составлять ценогенез в идеальном понимании. Сюда должны включаться собственно адаптивные изменения не только палингенетических признаков [358], но и любых других. Таким образом, ценогенезы - это любые преобразования, не связанные с эволюцией взрослой стадии и совершенствованием способа ее осуществления.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'