
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
"Эмбриональные модусы" и онтогенетическая запись
Рассмотрим теперь вопрос о реальности эволюции взрослой стадии путем одновременных изменений в раннем онтогенезе (архаллаксисов и девиаций). Как уже было сказано выше, это представление несовместимо с фактами, говорящими о целенаправленном (канализированном) характере онтогенетического процесса, основанном на регуляции промежуточных вариаций. Среди доводов, выдвигаемых в пользу возможности эволюции путем онтогенетических скачков, особое место занимают попытки найти ее следы непосредственно в самих онтогенезах. В этой связи нам придется сначала затронуть проблему исторического истолкования стадий развития в свете представлений о стабилизационных изменениях онтогенеза. Для этого проследим закономерности стабилизации на конкретном схематизированном примере - срастании костей голени в эволюции бесхвостых амфибий (рис. 30).
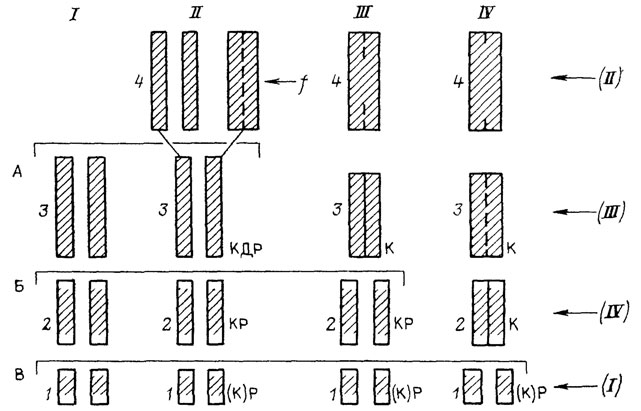
Рис. 30. Эволюция слияния элементов голени в истории бесхвостых амфибий. Составил М. А. Шишкин, оригинал. I - исходное состояние (пермские рахитомы), II - предки бесхвостых, близкие к раннетриасовому Triadobatrachus; III - промежуточный этап, IV - современные бесхвостые, 1-4 - стадии морфогенезов, А, Б, В - уровни зародышевого сходства между исходным типом развития и морфогенезами II-IV, (I)-(IV) - исторический порядок возникновения от дельных стадий морфогенеза IV, К (полное окостенение), Д (полное удлинение), Р (раздельность элементов голени) - рекапитулируемые признаки взрослого предка, f - внешний фактор, штриховкой показана степень окостенения
Это срастание, как и многие другие скелетные изменения, было связано у бесхвостых с передвижением прыжками, возникшим, по-видимому, как приспособление к мгновенному укрытию в воде от прибрежных хищников [1620]. У древнейших раннетриасовых Salientia (Triadobatrachus), как и у их рахитомных предков, соединения костей голени еще не было или же оно в какой-то мере проявлялось в позднем возрасте* (см. рис. 30, I, II). Облегчая прыжки, оно должно было в то же время затруднять нормальное четвероногое передвижение, уменьшая подвижность голени. Первоначально оба способа локомоции, очевидно, сосуществовали у древних Salientia, причем степень фиксации взаимоположения костей голени у взрослых животных могла в большой степени определяться интенсивностью функции прыгания, игравшей роль индуктора (см. рис. 30, II). По мере того как новая форма поведения становилась основной, отбор вел к ускоренному соединению, а затем и слиянию берцовых костей, так что оно начиналось уже у самых юных сформированных особей и притом все более автономно (см. рис. 30, III). Следующая эволюционная стадия (см. рис. 30, IV) типична для современных лягушек (Rana), у которых начало слияния приурочено к раннему метаморфозу [1562, 1826].
* (Последнее возможно в том случае, если единственный имеющийся образец Triadobatrachus в действительности представляет собой личинку в стадии позднего метаморфоза [1674]. У современных бесхвостых кости голени на этой стадии уже частично слиты.)
Этот схематизированный эволюционный ряд, представляющий собой пример гетерохронии, позволяет проиллюстрировать рассмотренные ранее закономерности. Мы видим, что новый признак может почти не меняться в филогенезе (см. рис. 30, II-4-IV-4), но его формообразовательный механизм преобразуется. Эти изменения захватывают все более ранние стадии: если сначала объединение голени, по-видимому, индуцировалось поведением взрослого животного, то у поздних форм оно детерминируется уже на стадии головастиков. Рекапитуляция исходной "двухкостной" взрослой голени, будучи практически полной вначале, у потомков все более обедняется. Если у формы II на стадии 3, предшествующей слиянию, рекапитуляция охватывает три учитываемых признака - окостенение (К), взрослое удлинение (Д) и раздельность составляющих элементов (Р), то в морфогенезе потомков эта стадия выпадает, а ее признаки все более теряют взаимосвязь, распределяясь по другим стадиям. Стабилизация ведет, таким образом, к стиранию палингенеза. Общей для всех форм остается лишь ранняя стадия с ясно выраженным признаком Р, выступающая как элемент зародышевого сходства. В целом видно, что рекапитуляции на поздних стадиях наиболее полны, но исторически недолговечны; напротив, на ранних они максимально обеднены, но устойчивы.
Этот ряд позволяет еще раз показать обусловленность закона К. М. Бэра стабилизационной перестройкой, распространяющейся у потомков "в глубь" онтогенеза. В нашем примере действительно видно (см. рис. 30, А-В), что при сравнении онтогенеза I со все более преобразованными (акселерированными) типами II-IV уровень дивергенции между ними смещается на все более раннюю стадию. И если у формы II конец онтогенеза еще представляет собой надставку над взрослой стадией (3) предка I, то уже у формы III поздний онтогенез (стадии 3, 4) выглядит как замещение этой предковой стадии. Как уже отмечалось нами, это постепенное преобразование онтогенетической записи из продлевающей в дивергентную и есть причина того, что биогенетический закон по отношению к каждому анцестральному состоянию выполняется достаточно полно лишь у ближайших потомков.
Данный пример демонстрирует еще одно важное следствие стабилизационного процесса: несоответствие между порядком появления онтогенетических стадий в филогенезе и онтогенезе [1176, 1178]. В наиболее измененном типе развития IV порядок стадий 2-4 прямо противоположен историческому. Взрослая стадия 4 в основных чертах унаследована здесь от далекого предка (II), тогда как стадия 3 появилась в филогенезе позднее (III), а наиболее ранняя - стадия 2 - только что возникла (IV). И если онтогенетически стадия 4 является следствием стадии 2, то исторически, наоборот, стадия 2 является следствием стабилизации развития стадии 4. И лишь в раннем развитии (тип IV, стадии 1, 2) обе последовательности в нашем примере совпадают.
Подобное несоответствие, в общем, не было новостью уже для Э. Геккеля, который фактически признал его, введя понятие ценогенеза. Об этом же говорит его апелляция к методу тройного параллелизма как средству проверки филогенетических реконструкций, а также его признание, что эти реконструкции всегда гипотетичны и во многом "опираются на интуицию" [цит. по 2079]. На невозможность прямого прочтения онтогенеза как эволюционной последовательности указывалось потом множество раз, главным образом в контексте критики биогенетического закона, причем нередко признают, что единственный способ достоверного истолкования онтогенеза - это знание предшествующего филогенеза [540, 1661, 2079, 2141, 2284]. Причину этого, однако, видят не в стабилизационном процессе, а в том, что эволюция онтогенеза может идти разными путями, но каким именно в каждом конкретном случае - мы не знаем. В дальнейшем будет показано, что это представление в основном есть просто следствие произвольной расшифровки результатов стабилизационного процесса. Именно воздействие последнего (вместе с ценогенезами) и приводит к тому, что в итоге "развитие организма не несет в самом себе метода для определения исторической значимости стадий развития" [540].
Этот вывод имеет решающее значение для оценки существующих представлений об эволюции онтогенеза. Мы можем убедиться, что мнение о возможности эволюции взрослой стадии посредством единовременных эмбриональных изменений, бытующее уже полтора века, в действительности почти целиком основано на примерах прямого исторического прочтения онтогенезов, т. е. на том же самом принципе, что и биогенетический закон, но доведенном до абсолюта. А именно: если два онтогенеза дивергируют от некоторой общей ранней стадии, то заключают, что одна из сравниваемых форм возникла из другой путем девиации на этой общей стадии [926, 2141]. В нашем примере (см. рис. 30) это означает, что из прямого сопоставления морфогенезов I и IV, дивергирующих на стадии начального окостенения разделенных элементов голени (1), мы должны были бы заключить, что слияние голени в эволюции лягушек произошло скачкообразно путем объединения ее зачатков у головастиков! Возможность того, что взрослый признак может быть древнее, чем его эмбриональная стадия, обычно не принимается во внимание, если нет прямых доказательств этого; уклонение онтогенеза потомка отождествляется с историей возникновения этого потомка.
Возникает парадоксальное противоречие, которого не замечают. С одной стороны, история первого появления нового взрослого признака есть свершившийся факт, который меняться не может; с другой стороны, разница между первичной и преобразованной онтогенетической записью этого события нарастает в эволюции непрерывно, по мере стабилизации (эмбрионализации) признака у все более отдаленных потомков. Например, на рис. 30 истинная история слияния голени у лягушек иллюстрируется филогенетическими этапами I и II, но она будет выглядеть каждый раз по-новому при сравнении состояния I с этапами II, III и IV, если считать, что разница исходного и итогового онтогенеза показывает нам в каждом случае "модус" возникновения признака. Указанное противоречие не возникает лишь в том случае, если морфогенез раз возникшего признака никогда более не менялся бы.
Самое удивительное, однако, то, что реконструирование онтофилогенетических событий указанным способом не мешает исследователям признавать гетерохронии, выпадения стадий и другие вторичные изменения морфогенезов, искажающие подлинную картину возникновения новых взрослых структур. Другими словами, вполне однотипные явления онтогенетической дивергенции между предком и потомком могут истолковываться в одних случаях как доказательство происхождения потомка путем девиации, а в других, если имеются переходные онтогенезы, - как результат эмбрионализации изменений, первично возникших в позднем развитии.
В нашем примере (см. рис. 30) это означает, что один и тот же онтогенез IV может быть расценен двояко: либо как результат ускорения развития взрослой стадии 4, возникшей в онтогенезе II, - если мы знаем всю последовательность I-IV; либо как изменение онтогенеза I путем девиации на стадии 1, если известны только I и IV. При этом выбор интерпретации не имеет никакой иной основы кроме наличия или отсутствия фактического материала по промежуточным историческим этапам. Этот дуализм методологии типичен для исследований, классифицирующих эволюцию онтогенеза по модусам. Например, у А. Н. Северцова в одной и той же работе можно найти аргументы для обоих взаимоисключающих способов интерпретации онтогенеза. С одной стороны, говорится, что древний взрослый признак со временем передвигается на ранние стадии онтогенеза [928, с. 141], с другой же, утверждается, что "если признак появляется сразу в зачатке данного органа ... и сохраняется ... до взрослого состояния, то мы можем с большой вероятностью сказать, что этот признак филогенетически возник путем эмбрионального изменения" [928, с. 129].
Возникает вопрос - на чем же тогда следует основывать конкретный выбор между этими двумя правилами? Другими словами, как гарантировать, что между сравниваемыми типами развития не существовала в каждом случае цепь онтогенезов, показывающих первично взрослое происхождение нашего "эмбрионально возникшего" признака? Этот вопрос, однако, не ставится, и в большинстве приводимых примеров, в частности в случае смещения вперед брюшных плавников у костистых рыб [930], мы находим один и тот же аргумент: если бы строение потомка возникло путем надставки (анаболии), то мы бы видели у него рекапитуляции предшествующих взрослых состояний; а раз этого нет, то имело место эмбриональное изменение. Этот довод означает на самом деле молчаливое допущение, что никакой перестройки онтогенеза при его длительном конечном наращивании не происходит - ибо только в этом случае можно было бы ожидать полного отражения всего ряда предков в онтогенезе потомков. И, однако, наряду с этим признаются "вторичные архаллаксисы" [926], или изменения, основанные на вторичных искажениях записи надставок, т. е. как раз на том явлении, которое неявно отрицается при доказательстве "истинных" эмбриональных модусов. Эта произвольность интерпретации еще больше бросается в глаза, когда она оказывается разной для вполне однотипных явлений. Так, утрата в развитии конечности лошади пятипалой стадии расценивается как вторичное искажение анаболии, а аналогичная утрата закладок боковых пальцев у змеевидных ящериц - как истинный архаллаксис [ср. 928, с. 158; 930, с. 428].
Обоснование "эмбриональных модусов" у других авторов строится на той же самой предпосылке - представлении о неизменности предшествующей онтогенетической записи при эволюции путем надставок. Например, А. Ремане [2141], также предлагающий систему модусов, на первый взгляд признает, что разница между двумя онтогенезами (или морфогенезами гомологичных органов) не может служить прямым указанием на ход филогении. Однако в противоречии с этим он тут же пытается доказать существование девиаций именно из сопоставления двух онтогенезов. Рассуждения А. Ремане в схематической форме повторяют ход рассуждений А. Н. Северцова [926] при доказательстве им девиации в развитии ноздрей у рыб семейства Scombresocidae и сводятся к следующему. Если онтогенезы двух форм дивергируют от стадии А, то в принципе это может быть результатом их дивергентной эволюции по способу анаболии от общего предка, у которого взрослый облик соответствовал стадии А, а онтогенез - предшествующим общим стадиям двух форм. Однако подобное расхождение двух онтогенезов часто происходит на столь недифференцированной стадии, что она заведомо не может быть рекапитуляцией взрослого предка. В этом случае следует признать происхождение одной формы из другой путем эмбриональной девиации на стадии А.
Из этих соображений видно, что в представлении А. Ремане анаболия означает полное сохранение стадии взрослого предка (и его онтогенеза) в онтогенезе сколь угодно отдаленного потомка, и нарушение этой картины для него представляется не иначе, чем результатом девиации. Нетрудно видеть, к каким выводам мы можем отсюда придти. Если, например, в развитии крокодила и птицы нет стадии, представляющей их общего архозаврового предка, то мы должны признать, что либо обе формы произошли путем девиации от эмбрионов каких-то третьих форм, либо что птицы произошли от эмбрионов крокодилов. На этом же основании можно вывести какой угодно организм из какого угодно, поскольку здесь требуется лишь чисто негативный критерий - отсутствие общей стадии онтогенеза, соответствующей общему взрослому предку!
Приведенные соображения ясно показывают неприемлемость сравнительно-онтогенетического обоснования "эмбриональных модусов". Видеть в эмбриональном появлении взрослого признака прямое указание на способ его появления в филогенезе - значит судить об эволюции онтогенеза, исходя из отсутствия такой эволюции. Этот способ реконструкции исторических событий предполагает столь буквальное и ненарушимое их отражение в онтогенезе, на какое никогда не претендовал даже биогенетический закон.
Все сказанное относится также к "архаллаксисам" и "девиациям", описанным у ископаемых животных. Методология здесь та же самая - прямое отождествление онтогенетического различия с историческим событием. Если новый взрослый признак проявляется в онтогенезе рано, то нередко заключают, что именно так он возник в филогенезе - без скидок на возможность его вторичной акселерации или, наоборот, ретардации прежнего ювенильного состояния. И это несмотря на то, что проявления гетерохронии наиболее обычны именно на палеонтологическом материале, где они встречаются, по словам В. Е. Руженцева [887], на каждом шагу*. Даже признавая, что предложенные А. Н. Северцовым критерии для распознания первичных и вторичных эмбриональных модусов по сути ничем не различаются, исследователи все же иногда продолжают выделять "истинные" архаллаксисы и девиации [708, с. 127]. Более того, в тех же самых работах, где описывается "скачкообразное появление" новых взрослых признаков в раннем онтогенезе, нередко можно найти примеры, когда эти же признаки у близких групп формируются постепенно, так что вывод об их вторичном сдвигании на ранние стадии как бы напрашивается сам собой. Например, Д. М. Раузер-Черноусова [861] описывает как девиацию раннее (в онтогенезе) появление субцилиндрических камер взрослого типа у фузулиниды Wedekindelia (Foraminifera), но тут же указывает, что типичным для фузулинид является позднее развитие таких камер. Точно так же выглядит описанная этим автором "девиация" (появление крупных камер) у Nodosaria nechaevi на фоне онтогенеза у других видов этого рода.
* (Палеонтологам еще со времен А. Хайэтта известно о выпадении ранних стадий при акселерации ("вторичные архаллаксисы" Северцова); это явление позднее называли липогенезом, или липопалингенезом [1386, 1863].)
В. Е. Руженцев [887] и Б. И. Богословский [91] считают девиацией дорсальное смещение сифона у климений (Ammonoidea), несмотря на то что среди ранних аммоноидей (гониатитов) подобное же, хотя и менее устойчивое, смещение проявляется постепенно - либо на поздних оборотах раковины (в подотряде Tornoceratina), либо только на средних, т. е. ценогенетически (в подотряде Goniatitina) [343]. Логично предположить, что климениевый способ развития возник либо путем акселерации первого варианта, либо фетализации второго. Эти допущения невозможно устранить на том лишь основании, что между гониатитами и климениями, вероятно, нет прямой связи [91]. Речь здесь идет о типичном способе эволюции данного морфогенеза, который мог осуществляться параллельно в разных филетических рядах.
И все же палеонтологи, начиная с А. Хайэтта и Л. Вюртенбергера, в большинстве своем признают, что основным способом эволюции является изменение (надставка или выпадение) поздних стадий развития [291]. Это мнение преобладает и в современных работах, например [306, 343, 627, 703, 861, 887, 965]. Из них видно, что все преобразования онтогенезов, реально наблюдаемые у ископаемых форм (помимо смены конечных стадий), - это в действительности либо провизорные ценогенетические изменения, либо различные гетерохронии и их последствия. "Девиации" палеонтологов - это конечные результаты гетерохронии, скрытых от нас неполнотой ископаемой документации.
Из сказанного понятно, что, говоря о роли раннеонтогенетических изменений в эволюции взрослого организма, следует различать две стороны проблемы: 1) вероятность их прямого воздействия на взрослую стадию в пределах одного онтогенетического цикла (т. е. вопрос о реальности девиаций развития) и 2) возможность постепенного перехода таких изменений во взрослое состояние в филогенезе по мере замедления развития и выпадения его конечных стадий у потомков (фетализация). Первая из этих возможностей, как мы видели, не согласуется с фактами, говорящими о канализированности (забуференности) развития, и принципиально не доказуема путем сопоставления дивергентных онтогенезов. Напротив, фетализация составляет хорошо известное явление, неразрывно связанное с прогрессивной эволюцией и подчиняющееся тем же самым закономерностям, т. е. распространяющееся в филогенезе от поздних стадий развития к ранним.
Принципиальная разница между "всплыванием" ранних стадий при фетализации и гипотетическими "эмбриональными модусами" часто не осознается исследователями, и первый процесс либо объединяют под общим названием с девиацией (педоморфоз в понимании Г. де Бира) [1317], либо считают разновидностью последней - "девиация с запаздыванием" у фораминифер - [261, 667, 629], либо отождествляют с архаллаксисом [527, 887, 1984]. Главная причина этого, по-видимому, в том, что при недоразвитии материалом для изменений взрослого состояния служат уже существующие провизорные морфологические особенности и их "всплывание" нередко понимают как скачкообразную замену одного фенотипа другим [1185, 2420]. В действительности это не так. Реализация ювенильного признака в онтогенезе и его дефинитивное проявление в ходе эволюции - это совершенно разные события, из которых первое совершается во всех индивидуальных циклах по заданной программе, а второе - достигается в поколениях шаг за шагом. Всякое адаптивное изменение взрослого фенотипа связано с последовательным отбором и стабилизацией мелких онтогенетических вариаций, направленных в сторону создаваемого изменения. Это касается и замедления развития, что в итоге и обусловливает постепенность утраты прежней взрослой стадии. Реальность такой изменчивости, составляющей потенциальный материал для регрессивных преобразований, показана, например, для келловейского аммонита Kepplerites, у которого "субкосмоцерасовая" стадия онтогенеза (соответствующая взрослому состоянию его потомка Kosmoceras) обнаруживает индивидуальные уклонения в сторону увеличения ее длительности за счет взрослой кепплеритной стадии [412].
Что же касается явлений, как будто бы показывающих "скачкообразную остановку" развития (на деле всегда осложненную гетерохрониями в развитии репродуктивной системы и другими перестройками), то они представляют собой в действительности следствие эволюции, а не ее причину. Эти явления существуют лишь в виде факультативной неотении, т. е. разновидности созданного отбором модификационного полиморфизма, когда развитие в зависимости от внешних условий либо останавливается на определенной стадии, либо идет до конца. Общеизвестный пример - мексиканская саламандра Ambystoma tigrinum среди хвостатых амфибий. Как и все адаптивные модификации, "скачки" такого рода легко обратимы в поколениях, и соответственно в плейстоценовой истории мексиканской амбистомы, прослеженной на ископаемом материале, отмечается чередование двух ее адаптивных состояний в зависимости от смены похолоданий и потеплений [2383].
Следует упомянуть о том, что, признавая раннеонтогенетические сальтации в качестве модуса эволюции, исследователи не обязательно имеют в виду одновременное распространение их действия на поздние стадии, т. е., по существу, могут подразумевать просто одноактное возникновение ценогенетических изменений. Иногда эти гипотетические явления достаточно четко отделяются от "классических" архаллаксисов и девиаций. Например, у О. Шиндевольфа [2223] такой скачок, или "типогенез", предшествует постепенному переходу онтогенетического новшества во взрослое состояние. А. С. Северцов [932] выделяет такие события как особые модусы филэмбриогенеза, действующие как архаллаксисы и девиации, но только в отношении промежуточных стадий, а не взрослой. Иногда, однако, эти различия не акцентируются. Так, у колониальных кораллов в качестве архаллаксиса [101] был описан переход личинки-основательницы от бокового прикрепления к осевому и превращение протокораллита-капсулы в базальный диск - безотносительно к ходу позднего развития, которое может при этом не затрагиваться изменениями.
Во всех подобных случаях вывод о способе эволюции ранних стадий развития основан на том же самом подходе, что и при обосновании скачкообразных изменений взрослой стадии, а именно, на отождествлении различия двух онтогенезов с историческим событием. В действительности, как уже было показано выше, каждый отрезок онтогенеза, завершающийся более или менее устойчивым морфологическим типом, эволюирует в общем случае, как и взрослая стадия, путем терминальных изменений и их последующего стабилизационного углубления.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'