
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
О некоторых примерах "скачкообразных изменений"
Рассмотрим подробнее некоторые из перечисленных ранее явлений, используемых для доказательства эмбриональных скачков и относящихся к преобразованиям в рамках уже существующих структур.
Изменения в числе. Сюда относятся классические архаллаксисы А. Н. Северцова (увеличение числа метамеров тела у змей, изменение иннервации брюшных плавников у костистых рыб) и иные примеры - изменение числа щупалец у Cnidaria и т. д. [2141]. Признание в этих явлениях эмбриональных скачков основывается на убеждении, что подобные структуры по своей природе могут возникать лишь дискретно и эмбрионально, как, например, новые сегменты тела у змей по мере его филетического удлинения [928].
В действительности мы снова видим здесь неоправданную параллелизацию хода нормального развития с историческим событием. При этом не учитывается, что на популяционном уровне объектом вариаций является не только число метамерных структур, но и время их закладки, а также длительность проявления. Отсюда очевидны две возможности возникновения таких структур, не отражаемые непосредственно в итоговом онтогенезе: 1) новый метамер может развиваться эмбрионально как регулируемая (временная) вариация, которая в последующих поколениях сохраняется все дольше, пока не переходит, наконец, во взрослое состояние; 2) допустимо думать, что у рептилий время завершения сегментации в хвостовом отделе (где она наступает позднее всего) может варьировать, вплоть до индивидуального появления лишнего метамера в постнатальный период (в филогенезе эта вариация стабилизируется и эмбрионализируется); в пользу этого говорят соотношения у амфибий, у которых число хвостовых метамеров может в норме резко возрастать уже после метаморфоза [2014].
Перенос половых признаков на противоположный пол. Это явление встречается в норме у многих животных, и обычно полагают, что оно происходит в эволюции скачкообразно [1174]. По мнению И. И. Шмальгаузена, более общим правилом является внезапное появление таких "заимствованных" признаков в эмбриональном периоде и лишь затем переход их у потомков во взрослое состояние. В то же время для многих случаев им фактически признается одномоментная перестройка всего развития - появление самцовой окраски у самок насекомых, ящериц, птиц и т. д. [см. 1192]. Механизм переноса объяснялся на основе данных по морфогенезу вторичных самцовых признаков у птиц, показывающих зависимость их детерминации от чувствительности тканей к уровню концентрации мужского (стимулирующего) и женского (тормозящего) гормонов. И. И. Шмальгаузен полагал, что эволюционная стабилизация исходно самцовых признаков, связанная с повышением их чувствительности к действию активирующих факторов и одновременной автономизацией от влияния тормозящих факторов, в конце концов приводит к тому, что концентрации этих веществ у самок делают для них возможной формообразовательную реакцию по самцовому типу, что и вызывает у них скачкообразное изменение.
Между тем вызывает возражения само представление о скачкообразности такого переноса. И. И. Шмальгаузен видит ее доказательство в том, что в популяциях многих видов среди самок типичного облика встречаются с той или иной частотой особи с признаками другого пола - например, самцовая окраска у ряда насекомых, рептилий и птиц; рогатые особи среди безрогих самок копытных и т. д. Но, как уже было сказано, подобные "мутанты" могут представлять собой относительно стабилизированные адаптивные типы внутри полиморфных систем. Спорадичность их проявления нельзя отождествлять со скачкообразностью эволюционного возникновения. Кроме того, множество наблюдений показывает, что переносимые половые признаки могут обнаруживать все градации в степени выражения - от едва заметных зачатков до полного развития, как, например, рога самок у разных видов антилоп или матка у самцов дельфина Phocaena communis. Потенциальная способность любой зиготы к реализации обоих полов, а также различных морфологических переходов между ними ("интерсексов") показана многими экспериментальными исследованиями, в особенности по межрасовым скрещиваниям - у шелковичного червя, бесхвостых амфибий и т. д.; при это чем менее значительно уклонение в сторону противоположного пола, тем более поздние стадии развития оно задевает [29, 257].
Все это показывает, что механизмы детерминации половых признаков обеспечивают возможность постепенного перехода между крайними альтернативами и что она действительно реализуется в эволюции путем последовательного накопления и закрепления промежуточных вариаций. Данный процесс может быть сопряжен с поздними онтогенетическими изменениями, которые затем углубляются в поколениях по мере усиления уклонения взрослого признака в сторону противоположного состояния (рис. 32). Или же, наоборот, переносимый признак обнаруживается сначала как временное уклонение раннего развития, а затем постепенно "всплывает" во взрослую стадию.
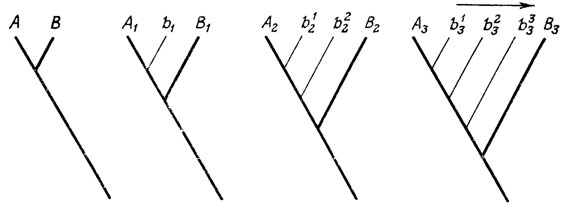
Рис. 32. Эволюционное расширение спектра промежуточных уклонений b1, b2, b3 по мере эмбрионализации развития альтернативных половых признаков А и B и углубления онтогенетической дивергенции между ними - до состояний A3 и B3. Составил М. А. Шишкин, оригинал. Непосредственный скачок от A2 к B3 возможен лишь как аномалия развития; эволюционно значимое изменение в направлении В3 осуществляется лишь через последовательную стабилизацию промежуточных уклонений (показана стрелкой)
Не подлежит сомнению, что явление переноса половых признаков существует так же давно, как и сам половой диморфизм. А поскольку половые признаки по своему происхождению связаны со взрослой стадией, то и древнейшие их нарушения должны были ограничиваться поздним онтогенезом. По мере филетической эмбрионализации, углубляющей дивергенцию между двумя альтернативными вариантами осуществления полового признака, все более расширяется спектр возможных нарушений выбора между ними. Чем раньше в онтогенезе наступает такое аномальное уклонение, тем больше его результат фенотипически приближается к противоположному нормальному состоянию (см. рис. 32). Нет никаких оснований думать, что эволюция идет путем непосредственного закрепления наиболее крупных из этих аномалий; факты по изменчивости выражения переносимых признаков говорят об обратном. Вместе с тем эмбрионализация развития полового признака создает условия для формирования в его раннем морфогенезе временных аберраций в сторону противоположного пола (таких, как, например, провизорная закладка яйцеводов у самцов млекопитающих).
Таким образом, наиболее вероятно, что эволюционный перенос половых признаков осуществляется путем последовательного закрепления и суммирования отбором небольших уклонений, начинающихся либо непосредственно с исходного взрослого состояния признака, с последующим распространением изменения на предыдущие стадии, либо с одной из ранних стадий, изменение которой может в дальнейшем постепенно перейти в дефинитивное состояние путем фетализации.
Возникновение зеркально-симметричных вариантов. Подобные инверсии, в особенности появление отдельных левозакрученных особей среди гастропод с типичной правозакрученной раковиной (или, реже, наоборот) - один из популярных примеров "архаллаксиса" [359, 2141; 2287]. Различие между двумя вариантами прослеживается здесь с первых же дроблений и может детерминироваться еще в материнском яичнике; скрещивание таких форм дает простое менделевское расщепление [2343]. Морфологические переходы между этими двумя типами невозможны, и обычно принимается, что один возникает из другого путем мутации. Так объясняют появление у гастропод левозавернутых форм.
На самом деле, однако, здесь нет ничего, что говорило бы о скачкообразных эволюционных изменениях. Наличие правильного расщепления ясно указывает, что речь идет о сложившейся полиморфной системе, и вопрос состоит в том, как исторически она возникла. Само по себе наличие право- и левозакрученности (и соответствующих типов асимметрии внутренних органов) - есть следствие конического (турбоспирального) навивания раковины, которое возникло у древнейших гастропод из симметричного планоспирального навивания [61, 1997]. Естественно считать, что исходным материалом для такого изменения послужили незначительные отклонения устья раковины от плоскости спирали и что лишь в последующих поколениях этот процесс стал захватывать все более ранние обороты, вплоть до макушки, соответствующей личиночному протоконху.
Уже первые неустойчивые отклонения такого рода неизбежно должны были осуществляться в двух вариантах - "правом" и "левом", причем их адаптивная ценность едва ли могла заметно различаться (рис. 33). Каждый шаг стабилизации этих отклонений в ходе отбора открывал путь к их дальнейшему усилению благодаря постепенному увеличению амплитуды возможных для них новых вариаций. Таким образом, могло идти параллельное преобразование двух зеркально-симметричных морф и углубление онтогенетической дивергенции между ними (см. рис. 33, I-VI). Реально в эволюции отдельных филумов (или на отдельных ее отрезках) частота одной из таких морф обычно остается очень низкой, что означает ее меньшую адаптивную ценность и недостаточную стабилизацию ее онтогенеза. Но, очевидно, всегда сохраняется потенциальная возможность для усиления отбора в пользу этой морфы (при соответствующем изменении условий) и перехода ее тем самым в статус основной нормы. Вероятный пример таких преобразований показывает для кайнозоя Англии морская гастропода Trophon antiquum, у которой в плиоцене в условиях относительно теплого климата преобладала левозавернутая форма, тогда как ныне доминирует правозавернутая [2419]. Пространственные изменения частот двух соответствующих морф известны у ряда современных видов (например, Fructicicola lantzi [218]).
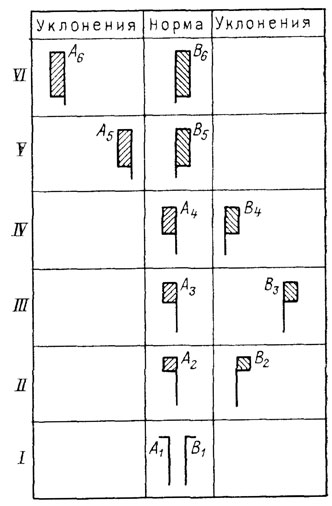
Рис. 33. Коэволюция двух возможных симметричных состояний фенотипической нормы (на примере право- и левозавернутости у гастропод). Составил М. А. Шишкин, оригинал. I-VI - последовательные исторические этапы; А1-A6, В1-В6 - последовательные стадии эволюции и эмбрионализации развития альтернативных морф (A и В). Заштрихованы отрезки онтогенеза, следующие за моментом дивергенции двух типов развития. Новый адаптивный признак (A-В) возникает в виде двух слабых и неустойчивых противоположных уклонений взрослой стадии на этапе I, усиливающихся и ускоряющихся в процессе дальнейшей эволюции этапы II-VI. Сужение нормы в пользу стабилизируемого варианта А ведет к снижению частоты симметричной ему морфы В, сохраняющей пониженную устойчивость осуществления (наследования, II-IV). Последующий сдвиг нормы в сторону морфы В стабилизирует ее и дестабилизирует фенотип А (V-VI)
Все это позволяет предполагать, что перераспределение адаптивных ролей между двумя типами навивания раковины является не только обратимым процессом, но и происходит на фоне их непрерывной параллельной эволюции. По мере углубления и спрямления онтогенетических преобразований, связанных с диссиметризацией внутренних органов в этих двух альтернативных вариантах, их дивергенция должна была начинаться в развитии все раньше, пройдя в итоге путь от дефинитивной стадии у билатерально-симметричного предка до первых фаз дробления у потомков.
Таким образом, наличие у гастропод двух глубоко дискретных типов развития с наибольшей вероятностью может рассматриваться как итог их постепенного исторического становления. Их предопределенность в яйце говорит об эволюционном скачке не больше, чем хромосомная детерминация пола в зиготе говорит о скачкообразном происхождении полового диморфизма.
Изложенная картина возникновения диморфизма раковины на основе планоспиральной организации взрослого предка требует некоторых дополнительных обоснований. В развитии асимметрии тела гастропод выделяют два основных момента: 1) торсионный процесс, т. е. поворот раковины с внутренностным мешком относительно головы и ноги на 180° и 2) приобретение раковиной турбоспиральной формы и отведение оси ее навивания назад, что вызывает одностороннюю редукцию парных органов. Независимо от того, развивались ли эти изменения синхронно или последовательно, есть все основания связывать их начало с переходом взрослого предка от пелагического обитания к жизни на дне [61, 430, 1997]. Весь ход развития гастропод несет на себе признаки филетического ускорения - "адьюльтации" [1785], приведшего к появлению зачатков ряда взрослых органов (нога, радула, раковина) на личиночных или эмбриональных стадиях. Поэтому и приуроченность торсионного процесса к концу личиночной жизни логично связывать с таким же сдвигом. В пользу справедливости этого мнения говорит сравнение разных типов личинок и в особенности соотношения у примитивного переднежаберного Haliotis, где вторая фаза скручивания наступает лишь после перехода к обитанию на дне и продолжается весьма долго [430]. Мнение У. Гарстанга [1624] о том, что скручивание возникло еще у личинок как результат мутации, вызвавшей одностороннюю редукцию мышц, представляет собой типичный пример отождествления истории возникновения признака с его специализированным морфогенезом*.
* (Другое возражение против взрослого происхождения скручивания [695], указывающее на невозможность втягивания тела в раковину при наличии перекреста первично парных мускулов, основано на представлении, что торсионный процесс осуществлялся у предка сразу же во всей полноте. Между тем речь должна идти о его постепенном филетическом усилении, параллельно с редукцией одного из мускулов.)
Все это позволяет считать, что сопутствовавший скручиванию переход к турбоспиральному типу навивания (а с ним и появление двух его зеркальных вариантов) начал впервые формироваться на поздних стадиях развития планоспирального предка.
Приведенные данные показывают, что реальная возможность эволюционных изменений на основе онтогенетических скачков крайне мала и при этом, вопреки распространенным представлениям, не имеет достоверных фактических доказательств. Однако из этого вовсе не следует, что эволюция имеет монотонный (градуальный) характер. Дело обстоит как раз наоборот. Каждый элементарный шаг в изменении адаптивной организации является результатом двух событий: 1) дестабилизации прежнего фенотипа, ведущей к уклонениям позднего онтогенеза (повышению изменчивости) и 2) стабилизации одного из таких уклонений в качестве новой нормы. Следовательно, на этом уровне рассмотрения эволюция предстает как постоянное чередование фаз устойчивости и неустойчивости (снижения адаптивности) или соответственно фаз персистирования и направленного изменения ([1180, 1181]; см. 7.3). Отбор направлен на поддержание первых, пока они сохраняют свое адаптивное значение, и скорейшее преодоление вторых. В этом смысле эволюция всегда прерывиста, ибо переход между двумя стабильными состояниями невозможен без снижения стабильности в промежутке. На макроэволюционном уровне этот механизм сдвига нормы служит основой как для постепенных ("филетический градуализм"), так и более неравномерных по своим темпам изменений. Однако ни на макро-, ни на микроуровне эта качественная неоднородность эволюционного процесса не имеет отношения к онтогенетическим скачкам (девиациям) и не подтверждает их существования.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'