
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
"Эмбриональные модусы" и мутационизм
Представления об эволюционно значимых изменениях раннего онтогенеза, одновременно меняющих взрослую стадию, основываются отнюдь не только на сравнительно-эмбриологических доводах. По существу, речь должна идти об обоснованиях всей совокупности мутационистских взглядов на эволюцию, ибо предполагаемое ими скачкообразное изменение организмов невозможно представить себе иначе, как результат ранних индивидуальных нарушений нормального хода онтогенеза. Необходимость допущения таких усклонений или даже перестройки всего развития подчеркивается многими авторами, признающими эволюционные сальтации, - например, когда говорится о "системных мутациях" [1650] или "онтомутациях" [1482], или же когда глубокие мутационные аномалии прямо отождествляются с арахаллаксисами А. Н. Северцова [2142]. Однако чаще этот аспект проблемы остается без внимания.
Независимо от способов аргументации в основе мутационизма всегда лежит явное или неосознанное убеждение, что между нормальным и аномальным итогом онтогенетического развития нет принципиальной разницы в отношении эволюционной устойчивости. Неприемлемость этого взгляда (см. 7.3) делает концепцию скачков несостоятельной для объяснения магистрального эволюционного процесса; однако следует помнить, что мир природных явлений бесконечно разнообразен, и нельзя исключить априори возможность осуществления в нем относительно маловероятных событий. К тому же граница между "ранним" и "поздним" онтогенезом, отделяющая область регулируемых или летальных уклонений от области эволюционно значимых вариаций, меняющих взрослый фенотип, всегда достаточно условна, и, строго говоря, мы можем лишь утверждать, что вероятность сохранения отбором онтогенетических изменений снижается тем резче, чем раньше они наступают. Поэтому важно определить, в каких случаях теоретически возникает наибольшая вероятность уклонения от типичных общих закономерностей онтогенетической эволюции, рассмотренных выше. Рассмотрим основные принципиальные аргументы в пользу мутационизма, относящиеся к собственно морфологической эволюции.
На первый взгляд, первостепенная роль здесь принадлежит палеонтологии, так как, начиная со времен Ж. Кювье и А. д'Орбиньи, она многократно порождала сальтационистские концепции (неокатастрофистские идеи О. Иекеля, Д. Н. Соболева, О. Шиндевольфа, Р. Ведекинда, К. Бойрлена, Э. Даккэ и др. [291, 318]) и продолжает выдвигать их и сегодня, например [527, 1878]. Но на деле используемые ею факты не могут играть в нашем вопросе решающей роли в силу самой специфики палеонтологической летописи, не позволяющей с уверенностью отличать истинную смену поколений от случайной преемственности, созданной седиментационными проблемами и локальными миграциями. Между тем для доказательства эволюционно значимых скачков необходимым (хотя и не достаточным) условием является именно регистрация их у непосредственных потомков. С этой точки зрения более существенны свидетельства, относящиеся к рецентным организмам.
Один из самых традиционных доводов в пользу скачкообразной эволюции, ведущий начало от Э. Жоффруа Сент-Илера, заключается в указаниях на появление в природе резких аномалий и, вообще, дискретных изменений как на путь возникновения новых форм (А. Келликер, У. Бэтсон и др.). Особое развитие он получил в контексте критики дарвиновской теории, когда ее главному принципу отбора мелких вариаций стали противопоставляться бесспорные случаи спонтанного появления у животных и особенно у растений (преимущественно домашних или культивируемых) крупных наследственных уклонений, давших начало новым устойчивым формам (С. И. Коржинский, Г. де Фриз и др.). Но еще Ч. Дарвин [310] указывал, что такие аномалии, используемые в практике селекции, имеют мало шансов закрепиться в природных условиях в связи с их редкостью и нарушением в них нормального развития, ведущим к снижению приспособленности.
Это возражение, в основном сохраняющее свою силу и сегодня для мира животных, часто считается менее весомым в отношении растений в связи с накоплением данных, свидетельствующих о большой жизнеспособности многих растительных мутантов в природе. Последнее заставляет многих авторов, даже мало склонных к мутационистским представлениям, допускать скачкообразное возникновение отдельных растительных групп через посредство таких аномалий [1194, 2249, 2520].
В последнее время оспаривают также и другое возражение Ч. Дарвина - о единичности и малой вероятности сохранения крупных аномалий в природе. При этом приводят случаи, когда подобные аномалии (большей частью экзогенного характера) дают широкие вспышки в небольших популяциях и могут удерживаться достаточно долго. Сюда относятся случаи массовой полимелии (появления лишних конечностей) и полидактилии (многопалости) у современных амфибий. На этой основе Л. Ван Валеном выдвинута новая модель эволюции, в которой подобные массовые аберрации служат объектом отбора (генетической ассимиляции) и, таким образом, становятся не зависимыми от первично вызвавшего их внешнего фактора [2420].
Весьма существенным доводом в пользу спонтанного появления новых таксонов у животных и растений считают факты близкого параллелизма между аномалиями одних родов и видов и нормальной организацией у других [1174, 1650, 2323]. Например, бескрылые виды мух по типу рудиментации крыльев могут очень напоминать соответствующих мутантов у дрозофилы. Особенно удивительными эти параллелизмы кажутся исследователям в тех случаях, когда речь идет об изменении числа однотипных органов или о переносе полового признака на противоположный пол. Так, по мнению В. М. Шимкевича [1174], наличие лишних ротовых отверстий у одного из реснитчатых червей или добавочной пары рогов у антилопы Tetraceros указывают на их происхождение из мутаций, аналогичных таким же уродствам у близких таксонов.
Но, пожалуй, самым распространенным источником представлений о скачкообразных эволюционных изменениях является господствующая сегодня синтетическая теория микроэволюции. Согласно ей, новые формы или расы во многих случаях возникают из старых путем единичной мутации, которая затем распространяется в популяции, приобретая ранг нормы [269, 1509]. С этих позиций обычно трактуются и популяционные изменения у ископаемых форм, когда распространение какой-либо особенности - например, новой складки зубов в эволюции лошадеобразных или новой формы раковины у двустворок, - расценивается как результат отбора в пользу мутантного гена [1713, 2287]. Сторонники этих взглядов обычно даже не осознают своей приверженности к мутационизму, полагая, что последний непременно связан с допущением крупных скачков типа "системных мутаций" Р. Гольдшмидта [594, 1509]. Но в действительности сущность мутационизма выражается не в масштабах допускаемого эволюционного уклонения, а в том, что оно мыслится возникающим в готовом виде независимо от естественного отбора, который затем лишь увеличивает его частоту.
Наиболее очевидными случаями спонтанных изменений, расцениваемыми нередко как единственная бесспорная возможность мгновенного видообразования [594], большинство эволюционистов считает межвидовую гибридизацию и полиплоидию (т. е. кратное увеличение числа хромосомных наборов в клетках). Различают прежде всего авто- и аллополиплоидию, т. е. соответственно изменения на основе одного видового генома или же двух (и более), объединенных у гибридов. Говоря о таких скачках, обычно имеют в виду не столько обязательное появление морфологических отличий, сколько внезапность репродуктивной изоляции от родителей.
Здесь снова обнаруживается поразительное различие между животным и растительным царствами. Если у растений полиплоиды обоих типов охватывают, по некоторым оценкам, до половины всех покрытосеменных и до 95% папоротникообразных [269], то у животных они намного более редки и охватывают лишь группы небольшого ранга (у наземных позвоночных - не выше вида). Обычно это автополиплоиды, связанные в огромном большинстве случаев с партеногенезом, но иногда включающие и группы бисексуальных видов, например у бесхвостых амфибий [ср. 594]. Аллополиплоиды (обычно триплоиды) нередко встречаются у животных среди межвидовых гибридов [111], которые в целом также характеризуются партеногенезом или другими формами клонирования (гибридогенез, гиногенез). Переход этих гибридов к однополости объясняется неизбежными нарушениями у них гематообразования при конъюгации в мейозе нетождественных родительских хромосом, что в свою очередь нарушает оплодотворение.
Попробуем теперь оценить степень доказательности указанных фактов. Прежде всего очевидна неубедительность многочисленных примеров "включения полезной мутации в норму", используемых синтетической теорией эволюции, поскольку под мутациями здесь в действительности подразумеваются уже сложившиеся адаптивные морфы или экотипы, стабилизированные в данное время (или в данной популяции) на низком частотном уровне, - например "мутации" меланизма у грызунов или жилкования крыльев у земляных ос [244, 2519]. Их дальнейшая экспансия означает просто перераспределение частот фенотипов в полиморфных системах. О предыстории таких "мутантных" фенотипов мы ничего не знаем; там же, где она известна хотя бы отчасти (как в случае меланистической морфы у бабочки Bistort betularia), факты говорят об их постепенном (а не скачкообразном) формировании [1181].
Наибольший интерес для решения нашей проблемы представляют явления параллелизма между аномалиями и нормальными признаками у разных таксонов. Исследователи, усматривающие в этом доказательство скачкообразного возникновения новых форм, не учитывают, однако, что подобие или тождество двух фенотипов не означает здесь ни их одинаковой устойчивости в поколениях, ни идентичности способов их онтогенетической реализации. С точки зрения представлений о пространстве (ландшафте) системы развития ([1180, 2441], рис. 25, 26, 31, а-в, ср. 7.3) эти способы глубоко различны. Аномалия соответствует индивидуальному уклонению развития от канализированного пути (см. рис. 31, б), означающему снижение его устойчивости, а значит, и воспроизводимости конечного результата. Напротив, эволюционный сдвиг в пользу того же фенотипа есть результат постепенной перестройки самого ландшафта системы, ведущего к стабилизации прежней аберративной траектории. Этот сдвиг суммируется из ряда более мелких шагов, каждый из которых связан с небольшим уклонением развития и его последующей стабилизацией, создающей основу для нового подобного шага (см. рис. 31, а).
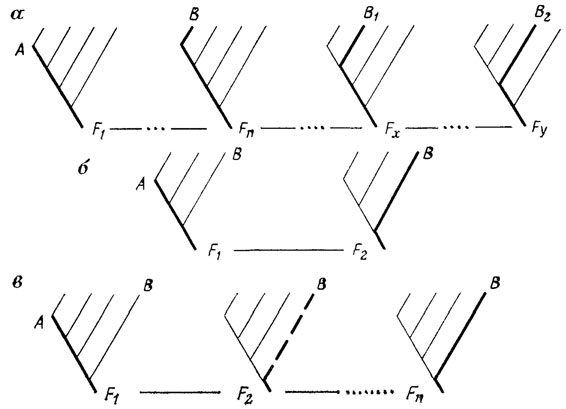
Рис. 31. Обычный (а) и гипотетические сальтационные (б, в) пути эволюции онтогенеза. Составил М. А. Шишкин, оригинал. а - постепенное преобразование фенотипа А в В2 путем смещения развития на промежуточные траектории В, В1 и их последовательной стабилизации (соответствующие типы развития F1, Fn, Fx, Fy разделены рядом преобразуемых отбором поколений); б - скачкообразное уклонение от исходного пути развития А на глубокодивергентную траекторию В в ходе одной смены поколений F1-F2, без участия отбора (классическое мутационистское представление о механизме эволюции); в - такое же, но неустойчивое скачкообразное уклонение с последующей стабилизацией его траектории путем отбора в ряду поколений F2-Fn (модель скачкообразной эволюции Ван Валена)
В соответствии с этой моделью в онтогенетических системах между нормой и выраженной аномалией (прообразом возможного эволюционного изменения) обычно в самом деле существует ряд непрерывных или дискретных промежуточных состояний, например последовательные градации выражения крыловой мутации vestigial у дрозофилы, связанные со все более ранними нарушениями онтогенеза (1650]. Поэтому приписывать, например, появление бескрылых видов насекомых скачкообразной утрате крыльев (и вообще отождествлять эволюционное изменение хода развития со скачком), значит, по меньшей мере не замечать реальных морфогенетических возможностей для постепенного приобретения нового признака. И действительно, во многих случаях природной редукции крыльев (среди жесткокрылых) могут наблюдаться самые различные степени ее выражения - как в пределах изменчивости одного вида, так и среди видов или родов одного семейства [1650]. К тому же аналогия между эволюционной и аномальной (мутантной) редукцией нередко оказывается достаточно поверхностной и на деле осложненной многими адаптивными изменениями в нормальном типе, как показывает, например, строение рудиментов крыльев у мухи Amalopteryx maritima, параллелизуемой Р. Гольдшмидтом с мутацией vestigial у дрозофилы [806].
Все сказанное относится и к специальным случаям подобных параллелизмов, связанных, например, с изменением числа нормальных органов. Мутантные и экзогенные аномалии такого типа обычно также показывают самые разные градации выражения, причем они могут выражаться асимметрично на разных сторонах тела - например, удвоение крыльев у дрозофилы, полимастия и полидактилия у млекопитающих и т. д. [30]. Поэтому трудно усмотреть здесь какие-либо доказательства скачкообразных изменений.
Как уже отмечалось, становление всякого эволюционного новшества неизбежно связано с отбором на канализированность (помехоустойчивость) его развития. Однако остается нерассмотренным вопрос - всегда ли оно идет путем накопления и фиксирования мелких изменений, задевающих конец онтогенеза, или же наряду с этим возможна непосредственная стабилизация (в ряду поколений) резкого одноактного раннеонтогенетического уклонения? Такая постановка вопроса как будто бы примиряет два противоположных подхода, совмещая крупномасштабность изменения с постепенностью его закрепления в ходе отбора, как это, в частности, предполагается упомянутой гипотезой Л. Ван Валена ([2420]; ср. рис. 31, в).
В том, что прямая стабилизация крупных аномалий принципиально возможна, нет, конечно, никаких сомнений. К ней, например, сводится процедура выведения в генетике "чистых линий" мутантов, а также механизм генетической ассимиляции индуцированных аномалий. К этой же категории явлений относятся и многочисленные примеры выведения новых сортов и пород на основе внезапных уклонений, традиционно используемые как возражение против дарвинизма.
Однако все эти примеры, в сущности, подчеркивают одно и то же: поразительный контраст между возможностью стабилизации крупных аномалий при искусственном отборе и отсутствием или редкостью достоверных случаев такого рода в естественных условиях. Причины этого достаточно понятны. Нарушение адаптивного фенотипа, порождаемое глубокой девиацией развития, в общем случае имеет шансы сохраниться и стабилизироваться лишь при одновременном резком и необратимом специфическом изменении внешних условий, дающем такой аномалии адаптивное преимущество. Вероятность подобного события очень мала. Такие ситуации пытались моделировать экспериментально, например, культивируя смешанные популяции дрозофилы в условиях постоянного ветра, когда крыловые мутанты, не способные летать, получали преимущество и вытесняли сдуваемых нормальных особей [1731]; подобные же явления отмечались и в узко локальных сезонных очагах в природе [2]. Однако эти наблюдения имеют не большую доказательную силу, чем любые случаи искусственной или вообще кратковременной в исторических масштабах селекции. Отбор в пользу любого фенотипа стабилизирует последний лишь применительно к условиям отбора; их исчезновение неминуемо ведет к расшатыванию и утрате этого фенотипа в последующих поколениях. Чтобы понять, насколько природная норма, сбалансированная по отношению к сложному комплексу условий, устойчивее временно стабилизированной аномалии, достаточно вспомнить многократно отмечавшиеся генетиками факты возвращения или приближения "чистых линий" мутантов к нормальному фенотипу при разведении их в свободной культуре или перенесении в природную среду [243, 707]. Отбор в исторически типичных условиях, таким образом, легко сводит на нет эпизодическое закрепление онтогенетических девиаций, лишая его эволюционной перспективы.
Следующий важный вопрос касается возможности скачкообразных изменений через посредство полиплоидии и гибридизации. Наименее значимой в этом плане следует считать автополиплоидию. Если в норме она связана с изменением кариотипа гамет, то это еще не доказывает, что именно так она и возникает в эволюции. Как у животных, так и у растений часто встречается соматическая автополиплоидия (эндополиплоидия), обусловленная изменениями митоза и закономерно охватывающая определенные ткани и органы, преимущественно в конце их развития [979]. Она может проявляться в соматических клетках и как обычная вариация, например, у потомства самок шелковичного червя, искусственно стимулированных к партеногенезу [31].
Поэтому случаи спонтанного появления полиплоидных гамет в природе не могут приниматься априори за достоверную модель эволюционного события. С не меньшей вероятностью мы можем считать, что кратное увеличение гаметного генома, наблюдаемое при типичной автополиплоидии, является в действительности не причиной ее, а, наоборот, конечным этапом ее эволюционного становления. В этом случае она должна сначала возникать митотически на поздних стадиях развития диплоидной формы и лишь затем сдвигаться в поколениях в глубь онтогенеза вплоть до зародышевых клеток.
Единственной очевидной возможностью преобразований, основанных на внезапном изменении типичной структуры зародышевой клетки (зиготы), является межвидовая гибридизация. Однако с точки зрения результата развития это событие в действительности не доказывает ни онтогенетического скачка, ни тем более создания какого-то новшества. Всякое нарушение нормального развития - вызвано ли оно абиотическим фактором, или влиянием чуждой гаметы - означает дестабилизацию, т. е. снижение жизнеспособности. Поэтому теоретически наибольшие шансы закончить развитие и дать нормальное потомство имеют те из гибридных особей, у которых ход онтогенеза остается предельно близким к одному из родительских вариантов, тогда как влияние противоположной гаметы низведено до уровня регулируемой помехи. Иначе говоря, отбор должен вначале благоприятствовать особям, фенотипически максимально близким к одному из родителей.
Косвенным подтверждением этих представлений служат многочисленные экспериментальные факты, показывающие, что ранний эмбриогенез у межвидовых гибридов (иглокожие, рыбы, амфибии) протекает обычно целиком по материнскому типу [357]. Очевидно, что чем позднее это доминирование нарушается, тем менее дестабилизированным оказывается итог развития. Следует добавить, что и при нормальном оплодотворении у огромного большинства животных раннее развитие (иногда вплоть до гаструлы) контролируется исключительно организацией материнской клетки, причем начальный этап функционирования диплоидного генома, по-видимому, сводится лишь к синтезу продуктов, уже существующих в этой клетке [333]. Все это заставляет думать, что у природных гибридных видов сохранение стенотипической близости к одному из родителей (описанное для ряда случаев) может служить существенным указанием в пользу их недавнего возникновения. Вероятное отсутствие скачка при формировании гибридных видов у животных касается не только их морфологии, но и пути перехода к типичному для них однополому размножению. В природе сохраняются зафиксированными различные стадии этого перехода, а именно, гибридогенез (скрещивание гибридов с самцами родительского вида, с последующим уничтожением мужского генома в оогенезе дочерних особей [111]) и гиногенез, где роль отцовской гаметы сведена лишь к стимуляции развития. Мы видим, что механизм включения развития лишь постепенно освобождается здесь от необходимости акта оплодотворения, делая существование гибридной формы все более независимым от контакта с родительскими видами.
Огромное различие в частоте случаев гибридного видообразования у животных и растений, несомненно, заключается в большей пластичности способов размножения у последних [31, 378]. Растительные гибриды через развитие соматической полиплоидии могут дать вегетативным путем обоеполое аллополиплоидное потомство, способное к нормальному гаметообразованию и продуктивному скрещиванию [269]. Напротив, для гибридов животных подобная возможность крайне затруднена, поскольку их существование поддерживается либо возвратным скрещиванием с родителями, либо однополым размножением. Ни в том, ни в другом случае не возникает эквивалентного по кариотипу полового партнера. Поэтому неудивительно, что двуполая аллополиплоидия у животных до сих пор достоверно неизвестна за пределами эксперимента [31, 111].
Проведенный обзор показывает отсутствие сколько-нибудь убедительных доказательств в пользу скачкообразной эволюции онтогенеза. Все приводившиеся с этой целью доводы основаны так или иначе на смещении в сознании исследователей продуктов эволюции с ее сырым материалом (истолкование адаптивных морф в качестве "мутаций", отождествление нормального развития с аберративным, отождествление дискретности нормальных фенотипов с онтогенетическим скачком между ними и т. д.). Верно лишь то, что многие изменения в механизме онтогенеза или в структуре зародышевой клетки создают потенциальную возможность для ускоренного преобразования взрослой организации у потомков, но само оно осуществляется лишь путем последовательной стабилизации в ходе отбора. Среди "подготавливающих" изменений такого рода наиболее специфический случай составляет, несомненно, межвидовая гибридизация, получившая массовое распространение в мире растений. Но поскольку, как мы видели, сама по себе она не доказывает одновременного уклонения взрослой стадии за рамки одного из родительских фенотипов, то возникает вопрос - действительно ли специфика онтогенеза у растений облегчает для них возможность скачкообразной эволюции, как это принято считать?
Этот взгляд кажется сильным упрощением. Систематика растений была бы невозможна, если бы их виды не обладали в общем случае определенной устойчивостью, которая и является выражением саморегуляции их онтогенеза. Последняя же как раз и представляет собой механизм, направленный на эффективное погашение ранних аберраций. Слабая морфогенетическая интегрированность растений, обычно подчеркиваемая как фактор, способствующий их сальтационной эволюции, в действительности играет здесь несколько иную роль. Чем менее интегрирована данная система, тем менее контрастны в ней различия между равновесным состоянием и отклонениями от него и тем более длителен для нее период восстановления нарушенного равновесия. В применении к живым системам это означает, что у растений отклонения от равновесного фенотипа (нормы) оказываются относительно "терпимыми" (жизнеспособными) в более широких пределах, чем у животных, и могут дольше удерживаться в поколениях.
Поэтому масштабы уклонений, для которых потенциально возможно закрепление отбором, у растений значительно шире. Иначе говоря, здесь возрастает вероятность случаев эволюции по схеме Л. Ван Валена - путем стабилизации относительно ранних аберраций. Эти случаи, однако, не означают девиации в общепринятом смысле, поскольку возникающие уклонения составляют здесь лишь сырой материал для создания устойчивой нормы в последующих поколениях. Тем более это не меняет общих закономерностей, рассмотренных нами ранее.
Сравнительный анализ приводит многих ботаников к выводу о преобладании в эволюции онтогенеза растений конечных изменений и о вторичном распространении их на более ранние стадии [2322, 2518], как это имеет место и у животных. К этому надо добавить, что сама по себе возможность утилизации отбором относительно ранних уклонений у растений еще не означает, что она доказана фактически. Наблюдая аномалии развития, мы не можем судить об их эволюционной судьбе; в то же время для уже возникших видов исходный тип развития никогда не может быть установлен вполне достоверно.
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'