
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Общая характеристика и морфология
Строение протоплазматического тела современных фораминифер
Протоплазма и ложноножки. Протоплазма фораминифер дифференцирована на внутреннюю эндоплазму, выполняющую полости раковины, и наружную эктоплазму, соприкасающуюся с внешней средой. По своему микроскопическому строению эндоплазма отличается относительной неоднородностью и содержит различные включения, ядра, а в некоторых случаях, возможно, распыленное ядерное вещество; эктоплазма по своей структуре более однородна.
В известные моменты жизни фораминифер эктоплазма может частично, а в некоторых случаях полностью превращаться в эндоплазму или объем эктоплазмы может иногда значительно увеличиваться за счет эндоплазмы. У большинства современных фораминифер в периоды, предшествующие размножению, как половому, так и бесполому, вся протоплазма втягивается внутрь; практически эктоплазма как таковая в это время отсутствует и тело фораминифер состоит в основном из эндоплазмы. У Elphidium crispum (Linne), наоборот, протоплазма перед началом бесполого размножения выходит из раковины и относительный объем эктоплазмы при этом значительно увеличивается. Через эктоплазму у фораминифер осуществляется обмен веществ с внешней средой; эктоплазма является местом образования ложноножек; она же выделяет раковину.
Эктоплазма у фораминифер с непрободенной раковиной нередко концентрируется перед отверстием раковины - устьем, образуя подобие пробки или затычки. При наличии сложного устья,состоящего из нескольких или многочисленных отверстий, эктоплазма растекается в пространстве между этими отверстиями и не представляет собою такого заметного обособленного образования, как в первом случае. У форм с прободенной, пористой раковиной обычно отмечается сплошная эктоплазматическая оболочка, облекающая всю раковину. При изучении под микроскопом фораминифер с хитиноидной (псевдохитиновой, ложнохитиновой) прозрачной раковиной иногда удается различить два слоя эктоплазмы: один - лишенный характерной для эндоплазмы зернистости, выстилающий стенку раковины с внутренней стороны, а другой, тоже более или менее гомогенный, облекающий ее снаружи (рис. 1).
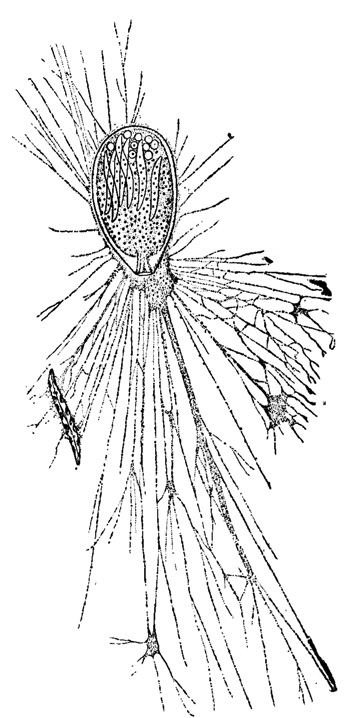
Рис. 1. Allogromia ovoldea Rhumbler. Раковина псевдохитиновая однокамерная в форме фляги. Вне раковины - эктоплазма, облекающая раковину сплошным чехлом и образующая разветвленные и анастомозирующие псевдоподии; эктоплазма выстилает раковину тонким слоем изнутри. В одном из сплетений ложноножек захваченная пища - диатомовая водоросль; кремневые оболочки переваренных водорослей видны и внутри раковины - в эндоплазме. Раковина обращена устьем вниз (Schultze, 1854)
Ложноножки, или псевдоподии, фораминифер являются в основном органоидами захвата пищи и передвижения; они играют у некоторых форм большую роль при построении раковины. Ложноножки представляют собою подвижные, сократимые, тонкие выросты эктоплазмы, длина которых может в сотню с лишним раз превышать их собственную толщину и быть в несколько раз больше поперечника раковины: у Elphidium crispum - в 2-3 раза, у Iridia lucida Le Calvez - примерно в 20 раз.
В ложноножках удается отличить более плотную осевую часть, состоящую из так называемой стереоплазмы, и внешнюю оболочку, образованную относительно жидкой реоплазмой, несущей центробежные и центростремительные токи зернышек. Стереоплазматическая ось при рассматривании ее в обычном проходящем свете представляется однородной, но в поляризованном свете удается установить ее способность к двупреломлению (Saedeleer, 1932).
Подобная особенность стереоплазмы отчетливо видима у ряда более высоко организованных фораминифер, обладающих известковой раковиной (например у Peneroplis, Elphidium, возможно, у милиолид); у большинства форм с агглютинированной раковиной эта особенность выражена слабо.
Передвижение фораминифер осуществляется путем повторного растягивания и сокращения ложноножек; при этом обычно животное распускает сеть ложноножек, приклеивается концами их к субстрату, а затем, сокращая ложноножки, при помощи них подтягивается. Скорость передвижения фораминифер изменяется от нескольких миллиметров до немногих сантиметров в сутки.
Основная функция ложноножек как органоидов пищеварения - захват пищи, которой могут быть различные микроорганизмы - низшие водоросли, простейшие, иногда даже личинки рачков и других микроскопических живых существ, а также органический детрит. При этом отмечается известная избирательная способность ложноножек, выражающаяся в том, что определенные пищевые частицы приклеиваются к их поверхности и немедленно облекаются тонким слоем протоплазмы, а другие, наоборот, не воспринимаются протоплазмой, не приклеиваются к ней или, в крайнем случае, если и облекаются, то остаются без изменений и вскоре выбрасываются. Кроме того, в ложноножках происходит, во всяком случае, хотя бы частичное, переваривание захваченных пищевых частиц, которые в дальнейшем транспортируются токами реоплазмы в центробежном направлении в эндоплазму, где окончательно ассимилируются. Наконец, ложноножки являются местом извержения (дефекации) непереваренных остатков пищи. Подобным же образом выбрасываются так называемые ксантозомы - желтоватые глыбки - отбросы обмена веществ, образующиеся в эндоплазме не только у форм, заглатывающих пищу, но и у видов, переваривающих ее вне эндоплазмы, в сети ложноножек. Ксантозомы нередко переполняют эктоплазму и, в частности, набиваются в систему каналов, если таковая имеется. Иногда в порядке удаления отбросов, фораминиферы выбрасывают прочь переполненную продуктами обмена веществ затычку в области устья [Planorbulina mediterranensis Orbigny и Elphidium crispurn (Linne), Le Calvez, 1953].
Совершенно несомненно, что ложноножки, широко распространяющиеся в толще воды, несут и дыхательную функцию: через их поверхность осуществляется газообмен организма с внешней средой.
Как показывают новейшие исследования (Le Calvez, 1938, 1953), ложноножки играют большую роль при выделении раковин. Процесс образования новой камеры раковины у Discorbis bertheloti (Orbigny) (рис. 2) протекает следующим образом. Сперва в области устья у основания последней камеры появляется веер ложноножек более значительный, чем обычно. Затем ложноножки укорачиваются и окружаются слоем различных посторонних частиц - детрита, из которого образуется вздутие - циста, до известной степени определяющее форму будущей камеры. Внутри цисты в течение нескольких часов длится интенсивная работа протоплазмы, заметная по энергичной циркуляции зернышек. Далее масса ложноножек сокращается еще больше, образуя плотное плазматическое тело, уже вполне отвечающее по очертаниям будущей камере; при этом между внешней поверхностью тела и внутренней поверхностью оболочки - цисты - сохраняется рыхлое сплетение ложноножек; вместе с тем поверхность вздутия покрывается тонкой хитиноидной уплотненной оболочкой. В поляризованном свете в наружных частях плазматического тела на этой стадии заметны небольшие блестящие зернышки, в результате слияния которых и образуется, очевидно, окончательная известковая раковина. Наконец, ложноножки выпячиваются через устье вновь сформировавшейся камеры; на первых порах они образованы одной протоплазмой, но вскоре в них появляются частицы захваченной пищи.
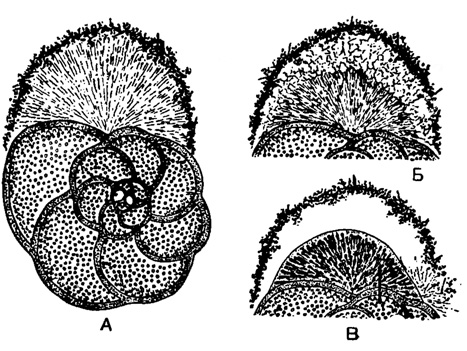
Рис. 2. Discorbis bertheloti (Orbigny). Образование новой камеры раковины: А - первое сокращение пучка ложноножек и образование оболочки - цисты из детрита; Б - второе сокращение ложноножек; уплотненный комок эктоплазмы определяет своей формой очертания будущей камеры; В - появление прободенной оболочки. Рисунок с живого объекта, × 40 (Le Calvez, 1938)
Весь процесс образования новой камеры длится у Discorbis около 8 часов. Образование стенки новой камеры происходит исключительно за счет деятельности эктоплазмы, в частности ложноножек; эндоплазма в этом процессе не участвует и внедряется изнутри раковины в уже окончательно сформировавшуюся камеру. Сходным путем образуются новые камеры и у Elphidium crispum, по данным М. Джеппс (Jepps, 1942); здесь при помощи особых длинных псевдоподий, в дальнейшем исчезающих, образуется временная циста - "маска" - из детрита, под которой происходит образование стенки новой камеры с поровыми канальцами и характерными для Elphidium септальными мостиками. С момента излияния формирующей новую камеру массы протоплазмы и до возобновления нормальной жизнедеятельности у Elphidium проходит около 24 часов.
Исключительно интересно поведение ложноножек у Planorbulina mediterranensis в первый период развития молоди (Le Calvez, 1945). Зародыши образуются в теле матери. Каждый из них окружен покровом ложноножек, выделяющим первичную раковину. В то же время по ложноножкам зародышей передаются питательные вещества в их эндоплазму из эндоплазмы материнского организма. Таким образом, ложноножки играют роль своего рода детского места - плаценты (рис. 3).
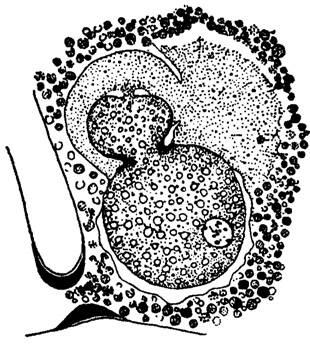
Рис. 3. Planorbulina mediterrnanensis Orbigny. Зародыш перед образованием третьей камеры; ложноножки играют роль плаценты, × 450 (Le Calvez, 1945)
Эндоплазма является наиболее объемистой частью протоплазмы; она отличается обычно более темной окраской, иногда желтой, красной, зеленой или коричневой по причине обилия темноокрашенных включений. Включения могут быть различного рода: во-первых, поступившие извне, как-то: пища, минеральные частицы, симбиотические водоросли - полезные сожители фораминифер, паразитические микроорганизмы; и, во-вторых, включения, являющиеся результатом обмена веществ, жизнедеятельности самого организма, а именно - продукты выделения - экскреторной деятельности организма - ксантозомы, жировые капельки, растворимые или не растворимые в жирах окрашенные вещества - пигменты, зернышки, воспринимающие основные микроскопические красители, так называемые базофильные зерна, мельчайшие зернышки - микрозомы и т. п. Точная микрохимическая характеристика различного рода включений протоплазмы фораминифер до настоящего времени отсутствует, и поэтому и функционально значение включений во многом остается неясным (Le Calvez, 1938, 1953).
Так, например, у Planorbulina захваченная пища, а также отбросы пищеварения находятся в протоплазме, занимающей последнюю или последние камеры и образующей "вегетативный сегмент"; физиологические функции этого "сегмента" в основном пищеварительные. Внутренний "хромативный сегмент" не содержит пищевых включений, но зато в нем, помимо ядерного вещества (обычно в виде одного или нескольких ядер), находятся микрозомы, базофильные зерна, пигменты и жировые капельки; все эти включения постепенно накапливаются в течение вегетативного периода жизни, а в процессе размножения проявляют значительную активность (Le Calvez, 1938).
У однокамерных фораминифер эндоплазма относительно однородна: включения располагаются в ней более или менее равномерно. У наиболее примитивных форм, таких, как Iridia, Webbinella, Rhynchosaccus, Technitella и у других агглютинированных форм, в эндоплазме содержится большое количество детрита Bathysiphon filiformis G. О. Sars, протоплазма сохраняется только лишь в незначительных просветах между бесформенными буроватыми иловыми посторонними частицами.
Перед наступлением процессов размножения эндоплазма претерпевает глубокие изменения, которые внешне выражаются в том, что она становится прозрачной или белой. Это связано с массовым выталкиванием во внешнюю среду отбросов и пигмента. Наиболее полное очищение протоплазмы происходит перед наступлением полового размножения (гамогонии), менее полное - перед образованием зародышей бесполым путем (шизогонии).
Цисты. Многие современные фораминиферы способны образовывать, помимо раковины, временные оболочки, или так называемые цисты. Цисты фораминифер состоят в основном из посторонних частиц: песчинок, игл губок, обрывков нитчатых водорослей, различных фрагментов животного и растительного происхождения, мелко раздробленного органического материала - детрита. Кроме того, в некоторых случаях в состав стенки цист, особенно образующихся в связи с процессами размножения, могут входить продукты, вытолкнутые протоплазмой: пустые оболочки переваренных микроорганизмов и различные протоплазматические включения, в том числе ксантозомы. Цементируется весь этот материал выделяемым ложноножками псевдохитином.
Цисты фораминифер образуются при различных условиях: при образовании камер, как это упоминалось выше, а также у ряда видов перед началом процесса размножения: в последнем случае циста окружает всю раковину, либо образует полушаровидный колпак, прикрывающий раковину, лежащую на субстрате. У некоторых прикрепленных фораминифер значительная часть раковины постоянно прикрыта цистой [Cibicides lobatulus (Walker et Jacob), Planorbulina mediterranensis Orbigny, некоторые виды рода Discorbis]. Песчанистые оболочки таких форм, как Iridia diaphana Heron-Alien et Earland и др., могут рассматриваться скорее не как агглютинированные раковины, а как цисты, лишь постоянно окружающие раковину в собственном смысле слова, в данном случае хитиноидную. Возможно, что первичные агглютинированные раковины возникли в процессе филогенеза из цист, способностью образовывать которые обладают, как известно, очень многие простейшие.
Ядерный аппарат. В нормальном состоянии все фораминиферы обладают, по меньшей мере, одним ядром. Однако имеются указания на существование у фораминифер и безъядерных стадий. Наличием одного ядра в течение всего жизненного цикла, за исключением периодов, непосредственно связанных с размножением, отличаются наиболее примитивные фораминиферы, а также планктонные формы (сем. Globigerinidae). У большинства фораминифер одноядерными являются так называемые гамонты, т. е. особи, образующие по достижении зрелости в процессе гамогонии половые элементы - гаметы: деление ядра у гамонта непосредственно предшествует образованию гамет. Шизонты, т. е. особи, размножающиеся бесполым путем с образованием в процессе шизогонии зародышей - мерозоитов (бродяжек, зооспор, "эмбрионов"), обладают у высших форм (в сем. Miliolidae, а также у родов Peneroplis, Elphidium, Planorbulina), как правило, значительным числом ядер уже на ранних стадиях своего вегетативного существования. У низших форм (Iridia, Myxotheca и др.) многоядерная стадия, наоборот, всегда кратковременна и наступает лишь к концу жизни шизонта.
В состав покоящихся ядер фораминифер входят ядерный сок, эндозомы, или ядрышки, и хроматиновое вещество, распыленное в ядер ном соке. Ядро имеет в той или иной степени отчетливо выраженную оболочку, представляющую собою перепонку, отделяющую содержимое ядра от вмещающей его протоплазмы. Ядерный сок - это студенистое вещество, в котором во взвешенном состоянии находятся структурные элементы ядра. Эндозомы (Hofker, 1930), или ядрышки (нуклеолы) - многочисленные включения, встречающиеся в ядрах большинства фораминифер и наблюдаемые у живых особей, благодаря сильной светопреломляемости. Окрашиваются эндозомы как кислыми, так и основными красками, но характерной для хроматина, точнее для тимонуклеиновой кислоты, реакции Фёльгена они не дают (Le Calvez, 1938, 1953). Хроматин лишь у пластогамных форм с амёбоидными гаметами, таких, как Spirillina vivipara Ehrenberg и Раtellina corrugata Williamson, достаточно восприимчив к специфическим для него ядерным краскам; у этих форм в покоящемся ядре могут наблюдаться хромозомы в виде спутанных нитей, содержащих в себе базофильные (т. е. окрашивающиеся основными красителями) зерна - хромомеры. У прочих фораминифер хроматин в покоящемся состоянии ядра почти не различим; обнаружить его удается лишь при обычно наблюдаемых у фораминифер кариокинетических (митотических) делениях ядра.
В зависимости от характера ядрышек, или эндозом, Л. Румблер (Rhumbler, 1911-1913) выделил три типа ядер фораминифер - суспензоидный, эмульсоидный и альвеолидный (Le Calvez, 1938). Первый тип - суспензоидный, свойственный большинству песчанистых однокамерных форм (Bathysiphon, Rhabdamtnina, Technitella, Pelosina, Saccammina, Hippocepinella, Rhynchosaccus, India, Myxotheca, Webbinella), а также представителям сем. Lagenidae (Dentalina, Nodosaria и др.). Эндозомы ядер суспензоидного типа сферические, маленькие, неодинаковой величины: расположенные ближе к центру относительно мелкие, по мере приближения к периферии становящиеся крупнее. По мере роста ядра количество эндозом постепенно увеличивается за счет их почкования или деления с образованием бисквитообразных фигур.
Второй тип ядер - эмульсоидный. Он свойственен ядрам многокамерных фораминифер (отр. Miliolida, Discorbis, Ероaides, Cibicides, Planorbulina, Elphidium). Эти ядра по форме очень изменчивы: они имеют то тонкогубчатую, то пузырчатую структуру; с поверхности иногда изрыты углублениями.
Третий и последний тип ядер - альвеолидные - пузырчатые, характеризуются смешением эндозомного вещества и кариоплазмы. Подобного типа ядра отмечались для пелагических фораминифер [Orbulina universa Orbigny, Globigerina, Hastigerina pelagica (Orbigny)]. У двух первых вещество эндозом собрано в одной половине ядра, тогда как другая занята относительно прозрачным веществом.
Характер деления ядра у фораминифер зависит в основном от двух причин: от степени насыщенности ядра хроматином и от фазы жизненного цикла. У видов, отличающихся значительным содержанием хроматина, митозы шизогонии не отличаются заметно от митозов, происходящих от гамогонии.
Типичные кариокинезы наблюдаются при размножении ядер (митоз в собственном смысле слова) и при делениях созревания, происходящих у фораминифер в конце шизогонии, перед образованием мерозоитов (так называемый мейоз) у Spirillina vivipara Ehrenberg (Myers, 1936) и Patellina corrugata Williamson (Le Calvez, 1938), также при гамогонии у Iridia lucida (Le Calvez, 1938), (рис. 4) и др., а при шизогонии у Discorbis vilardeboanus (F∅yn, 1937).
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'