
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Размножение и жизненный цикл
Уже во второй половине прошлого столетия некоторые исследователи обратили внимание на то, что у фораминифер наблюдаются виды, группирующиеся попарно. Так, Ф. Гарп (Harpe, 1879) отметил, что у нуммулитов из одного и того же местонахождения встречаются такие "парные" виды, которые сходны между собой во всех морфологических признаках, за исключением лишь того, что у одного из них начальная камера очень мала, неразличима невооруженным глазом, а общие размеры и число оборотов относительно велики; у другого вида - партнера, наоборот, начальная камера крупная, хорошо заметна, общие размеры при этом меньше, и раковина образована сравнительно небольшим числом оборотов. Е. Мюнье-Шальма (Munier-Chalmas, 1880) указал на то, что мы в данном случае имеем дело не с самостоятельными видами, а с двумя различными формами одного вида - с явлением диморфизма. Природу этого последнего Мюнье-Шальма объяснял возрастными отличиями.
В 1886 г. Мюнье-Шальма и Шлюмберже (Schlumberger) предложили термины "микросфера" для маленькой начальной камеры одной из упомянутых форм и "мегалосфера" для крупной начальной камеры другой формы. В дальнейшем эти формы получили, соответственно, название микросферической, или формы В, и мегасферической (мегалосферической или макросферической) или формы А (рис. 26, 27).
Истинная природа явления была выяснена работами Дж. Листера (Lister, 1894, 1895, 1903) и Ф. Шаудинна (Schaudinn, 1894,1895,1,2, 1903). Эти исследователи установили, что у представителей рода Elphidium и некоторых других имеет место чередование поколений - полового и бесполого. Дальнейшие исследования Ф. Винтера (Winter, 1907), Е. Майерса (Myers, 1933, 1934, 19351,2, 1936, 1940 и 1943), Ле Кальве (Le Calvez, 1937, 1938, 1950, 1953), Джеппс (Jepps, 1942) и др. подтвердили выводы Листера и Шаудинна и значительно приблизили нас к познанию истинной природы процессов размножения у фораминифер.
Жизненный цикл фораминифер распадается на два основных этапа: шизогонию, или агамогонию - бесполое воспроизведение со стадией образования мерозоитов (эмбрионов) и заканчивающееся образованием гаплоидного поколения - мегасферических гамонтов; и гамогонию - половое воспроизведение, в конечном итоге которого восстанавливается диплоидное состояние - образуются микросферические шизонты; этот второй этап сопровождается образованием многочисленных половых элементов - гамет и попарным слиянием этих последних.
Гаметы фораминифер, образованные как одной материнской особью - гамонтом, так и разными, не отличаются ни по своим размерам, ни по строению, чем определяется наличие изогамии. У большинства фораминифер гаметы жгутиковые - так называемые флагеллоспоры, но у некоторых (Spirillina и Patellina) гаметы амёбоидные. В. А. Догель (1951) указывает, что строение гамет является важным указанием на характер предковых форм фораминифер: по его мнению, гаметы представляют исходную стадию онтогенеза фораминифер, которая отражает, как правило, в своем строении особенности жгутиковых предков фораминифер. Догель считает, что амёбоидные гаметы некоторых фораминифер являются ценогенетическим новообразованием, что утрата жгутиков произошла в процессе приспособительного изменения хода онтогенеза в течение филогенетического развития группы. Если отказаться от интерпретации Догеля и рассматривать особенности строения гамет как унаследованные от предков - палингенетические, то придется допустить полифилетическое происхождение фораминифер от двух корней - жгутикового и амёбного.
Жгутиковая гамета фораминифер - округлоовальное образование размерами 1,5-2 μ, реже до 5 μ, снабженное двумя жгутами, неравными, вопреки мнению Шаудинна, из которых один, более короткий, направлен назад. В протоплазме гаметы можно различить ядро и сильно преломляющее свет жировое включение. У некоторых фораминифер гаметы снабжены тремя жгутами.
Копулируют у фораминифер гаметы, образованные разными гамонтами; автогамии, т. е. копуляции гамет, происшедших из одного и того же гамонта, не отмечалось. Слияние гамет происходит обычно в свободной воде, вне материнской раковины.
У многокамерных форм в результате слияния гамет образуется шаровидная зигота, которая несколько разрастается и выделяет на своей поверхности оболочку первой камеры микросферической формы. Развивающаяся из зиготы особь - шизонт у фораминифер - сохраняет одноядерное строение только на ранних стадиях своего развития; очень рано, еще задолго до начала характерного для него процесса бесполого размножения - шизогонии, он становится многоядерным путем последовательных делений ядра.
В дальнейшем, с достижением шизонтом более или менее определенных специфических размеров, его протоплазма разбивается на ряд участков соответственно числу ядер; каждый такой участок обособляется и превращается в одноядерного мерозоита. Образование мерозоитов может происходить внутри раковины материнского шизонта (India, Nubecularia, Peneroplis, Planorbulina) или вне ее (Elphidium); в последнем случае протоплазматическое тело заблаговременно покидает раковину.
То обстоятельство, что у фораминифер наблюдается чередование поколений диплоидного и гаплоидного, т. е. то, что они являются диплогаплонтами, представляет большой интерес; подобной особенностью отличаются многие низшие растения, но у животных диплогаплоидия до сих пор не отмечалась.
В результате копуляции гамет, у Discorbis образуется амёбоидная зигота; ее ядро неоднократно делится, благодаря чему молодой шизонт уже при выходе из цисты размножения содержит 8-16 ядер. В этом отличие от образующихся при шизогонии мерозоитов, которые остаются длительное время одноядерными. У Discorbis дифференцировка на микро- и мегасферические раковины сказываются не в величине их начальной камеры, а в общих размерах и направлении навивания спирали.
Гаметы Spirillina vivipara представляют собою амёбоидные образования размером около 10 μ. В результате попарного слияния гамет образуются амёбоидные зиготы; при этом происходит слияние пронуклеусов - ядер гамет.
У Patellina corrugata Williamson и различных, видов рода Discorbis образование зародышей также происходит вне раковины, так как от этой последней остается только крышечка, прикрывающая комочек протоплазмы, в котором происходит образование мерозоитов; перегородки между камерами раковины - септы, а также ее брюшная стенка растворяются незадолго до начала шизогонии. Иногда перед этим процессом, фораминифера окружается цистой, внутри которой и происходит размножение.
Вышедший из материнской раковины или обособившийся от материнской протоплазмы зародыш - мерозоит является, по сути дела, молодым организмом, обладающим всеми основными особенностями взрослого; он состоит из протоплазмы, дифференцированной на эндо- и эктоплазму, и снабжен ядрами, имеет раковину, может самостоятельно передвигаться и питаться при помощи псевдоподий. Поскольку процессы созревания ядра (Le Calvez, 1953) протекают в конце шизогонии, мерозоит гаплоиден. Подобное состояние сохраняется и во времени дальнейшего роста зародыша, когда он постепенно превращается в гамонта.
Le Calvez (1953) различает гамогонию двух типов: моногамную и пластогамную. В первом случае (рис. 4) образование гамет происходит в изолированных гамонтах, во втором (рис. 5, 6) - до образования гамет - два гамонта или более объединяются в общий так называемый сизигий. При моногамной гамогонии не все ядерное вещество используется при образовании гамет, а лишь небольшая его часть - "микронуклеусы", тогда как остальная погибает. Наоборот, у пластогамных форм при образовании гамет используется все ядерное вещество.
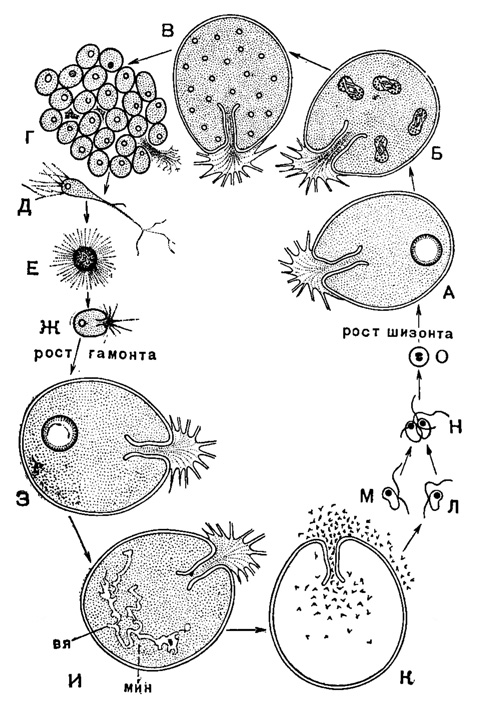
Рис. 4. Жизненный цикл India lucida Le Calvez - моногамный вид с двужгутиковыми гаметами: А-Ж - шизогония: А - почти взрослый шизонт; Б - митозы в конце вегетативного периода жизни шизоита; В - зрелый шизонт; прекращение питания и очистка протоплазмы; Г - образование эмбрионов; Д - самостоятельный гаплоидный зародыш - мерозоит с пучком псевдоподий; Е - пелагическая стадия существования зародыша; Ж - зародыш, возвратившийся к бентонному образу жизни, находящийся на пути превращения в гамоита; налицо видовые особенности: хитиноидная раковина и устьевая затычка; З-О - гамогония: З - начало самостоятельного питания гамонта; И - зрелый гамонт, переставший питаться; начало гамогонии - обособление вегетативного ядра (вя) и репродуктивного микронуклеуса (мин); К - окончание гамогонин - в результате последовательных митозов образуются многочисленные гаметоциты, дающие путем деления двужгутиковые гаметы, покидающие раковину; Л, М - гаметы, происходящие от двух различных гамонтов; Н - копуляция гамет; О - зигота (Le Calvez, 1953)
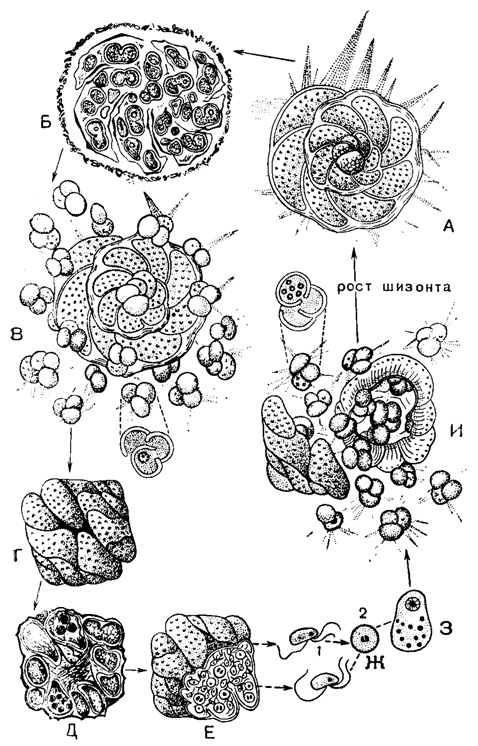
Рис. 5. Жизненный цикл Discorbis patelliformis Brady - пластогамный вид с трехжгутиковыми гаметами: А-В - шизогония; А - зрелый диплоидный и многоядерный шизонт с дискондальной раковиной и навиванием спирали вправо, с крупной начальной камерой (18 μ в диаметре); Б - образование зародышей - мерозоитов с двукамерной и трехкамерной раковиной; В - рассеяние зародышей благодаря растворению брюшной стенки материнской раковины; одноядерные зародыши с крупной начальной камерой (16 μ) начинают самостоятельно питаться. Г-И - гамогония: Г - слияние двух зрелых гамонтов - образование сигизия; каждый гамонт гаплоидный, одноядерный, с конической навитой влево раковиной, относительно малых размеров; Д - митозы гамогонин ядра каждого из гамонтов; растворение промежуточных стенок слившихся особей; Е, Ж - образование трехжгутиковых гамет (I), и их копуляция - образование зиготы (2); 3 - рост зиготы за счет поглощения не копулировавших гамет; видно ядро зиготы и погибающие ядра ассимилированных гамет; И - формирование молодых шизонтов в сизигии, их освобождение, деление ядра, образовавшегося при копуляции (Le Calvez, 1953)
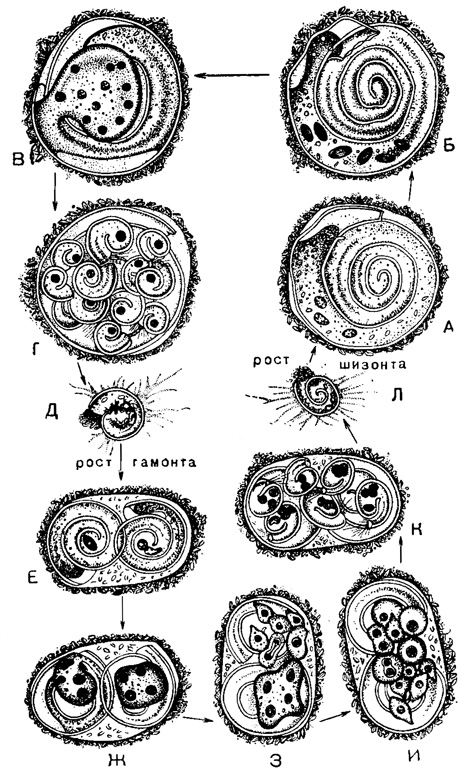
Рис. 6. Жизненный цикл Spirillina vivipara Ehrenberg - пластогамный вид с амебоидными гаметами: А-Г - шизогония: А - зрелый диплоидный шизонт крупных размеров; прекращение питания и формирование цисты; Б - деления созревания, происходящие к концу времени существования шизоита; В - накопление многоядерной протоплазмы в пупочной области; Г - формирование зародышей - мерозоитов и их выхождение, благодаря растворению части брюшной стенки материнской цисты; Д-И - гамогония: Д - молодой свободноживущий гаплоидный одноядерный гамонт; раковина из начальной камеры и короткой следующей за ней трубки; начало питания и роста; Е - слияние двух зрелых гамоитов - образование сизигия; гамоиты небольших размеров, одноядерные, начало митоза; сизигий окружен общей цистой копуляции; Ж - четырехъядерные гамоиты - плазмодии покидают свои раковины; З - двукратное деление каждого плазмодия на четыре гамегоцита; каждый из последних снова немедленно делится, в результате образуется восемь амебоидных гамет; И - гаметы и зиготы в общей оболочке сизигия; К - образование раковин будущих шизонтов; послекопуляционные деления дают по четыре ядра в каждом молодом шизонте; одно из ядер обычно дегенерирует; Л - молодой, вышедший из оболочки сигизия диплоидный шизоит с 3-4 ядрами; начало питания и роста (Myers, 1936 из Le Calvez, 1953)
К моногамным видам Le Calvez (1953) относит Elphidium crispum (Linne) (рис. 14), Peneroplis pertusus (Forscal,) Myxotheca arenilega (Schaudinn), Iridia lucida Le Calvez (рис. 4) и Planorbulina mediterranensis Orbigny.
В результате многократных делений "микронуклеусов" гамонт делается многоядерным. Отдельные ядра становятся центрами образования так называемых жгутиковых гаметоцитов, в результате деления которых надвое по типу обычного продольного деления жгутиконосцев получаются половые элементы - гаметы. Образование гамет происходит всегда внутри материнской раковины, которую в известный момент гаметы покидают в виде роя, через устье; этот процесс происходит обычно ночью. Гаметы известны для целого ряда моногамных видов фораминифер.
К числу пластогамных видов относятся Spirillina vivipara Ehrenberg, Patellina corrugata Williamson, а также Discorbis patelliformis Brady, D. pulvinata Brady и др., относимые обычно к Discorbis s. str., но отличающиеся некоторыми особенностями строения раковины и заслуживающие, возможно, выделения в особый род, для которого Гофкер (1951) предложил особое название Conorbella.
Шаудинн (1895), открывший пластогамию, не связывал ее с половым размножением. Эту связь установил Майерс (1933, 19351,2, 1936 и 1940); в дальнейшем она была подтверждена исследованиями Ле Кальве (1938) и др.
Для явлений пластогамии у Discorbis (рис. 5) характерно слияние плазмы двух особей в единый - общий сизигий; при этом происходит прочное объединение раковин партнеров. Половые процессы начинаются с взаимной химической активизации, затем осуществляется переход всего ядерного вещества в образующиеся гаметы, отличающиеся относительно крупными размерами и наличием трех жгутов. При образовании сизигия эктоплазма партнеров растворяет брюшную стенку раковины, а также септы. В результате ряда последовательных делений ядра и обособления участков протоплазмы, вокруг конечных продуктов этого деления образуются гаметоциты. Эти последние делятся, по крайней мере, еще раз, образуя упомянутые трехжгутиковые гаметы,остающиеся внутри полости,образованной двумя слившимися партнерами, и здесь копулируют попарно.
У Spirillina vivipara Ehrenberg (рис. 6) явления, происходящие при гамогонии, отличаются от того, что наблюдается у Discorbis, во-первых, меньшим количеством гамогонических делений ядра, а во-вторых, амёбоидным характером гамет. Гамонт у Spirillina vivipara существует самостоятельно недолго; вскоре две особи сближаются, сливаются псевдоподиями, после чего окружаются общей цистой. Ядро каждого гамонта делится два, реже три раза подряд; четырехядерные плазмодии партнеров вытекают под свои раковины в полость общей цисты, где каждый плазмодий делится на четыре одноядерных образования, представляющих собою гаметоциты. В результате однократного деления гаметоцитов образуются амёбоидные гаметы, числом восемь из каждого гамонта.
Сходно, по данным Майерса (1933,1934), протекает жизненный цикл Patellina corrugata Williamson.
В некоторых случаях чередование поколений у фораминифер осложняется за счет того, что между двумя поколениями гамонтов вклинивается не одно, а два поколения шизонтов. Цикл развития протекает не по типу: А - В - А - В - А... (где А - гамонты, а В - шизонты), а по типу: A1 - А2 - В - А1 - А2 - В - A1 - А2... (где А2 представляют собою шизонтов же, но второго поколения). Деления созревания падают в данном случае на конец существования поколения А2. Такие случаи приводят к триморфизму раковин, описанному Гофкером (Hofker, 1930).
Ле Кальве (1953) предполагает, что образование поколения А2 носит скорее случайный характер и происходит благодаря тому, что при шизогонии некоторый процент мерозоитов образуется без предшествующих делений созревания, т. е. остается диплоидным; такие мерозоиты не могут превратиться в гамонтов (последние, по мнению Ле Кальве, должны быть обязательно гаплоидными) и "повторно" становятся шизонтами.
Наличие закономерной смены трех поколений у некоторых, по крайней мере, форм подтверждается наблюдениями Гофкера (1930) над Streblus beccarii (Linne) var. flevensis Hofker.
Гофкеру удалось показать, что различные генерации Streblus beccarii var. flevensis приурочены к различным временам года. Жизненный цикл этой разновидности является годичным, а отдельные его стадии - сезонными. Зимой и весной встречаются исключительно представители поколения В, т. е. шизонты; зимой они молодые - их раковины образованы двумя-тремя оборотами, а весной, в мае, попадаются уже взрослые особи. Начиная с мая появляются мегасферические особи поколения А1, с июля - А2. В ноябре удается обнаружить впервые молодь поколения В. В связи с размножением путем шизогонии, в мае и июне отмечается инцистирование форм В. В июле, в связи со второй шизогонией, сопровождающейся делениями созревания, инцистируются формы A1. Формы А2 (гамонты) инцистируются в конце августа - начале сентября, когда имеет место гамогония.
У Spiroplectammina sagittula (Defranсe) форма В отличается двухрядным спирально-винтовым текстуляроидным строением как раннего отдела раковины, так и последующих; она однотипна по своему строению - мономорфна. Формы A1 и А2 биморфны - сочетают в себе два типа строения, отличаются от микросферической наличием начального отдела, имеющего спирально-плоскостное строение.
Чаще наиболее резко гетероморфное (би- или триморфное) строение раковины бывает выражено у микросферических форм, что особенно хорошо можно видеть на примере различных милиолид (рис. 7). У Bigenerina nodosaria Orbigny биморфное строение выражено в обеих генерациях. Форма А отличается более крупной начальной камерой, меньшими размерами, меньшим общим числом камер и значительно меньшим развитием начального двухрядного, текстуляроидного отдела (рис. 8).
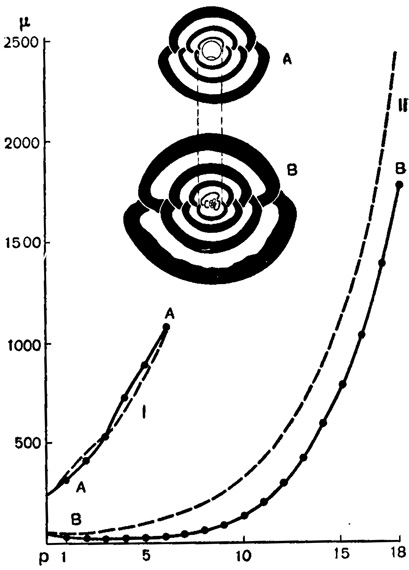
Рис. 7. Pyrgo bradyi (Schlumberger). Мегасферическая (А) и микросферическая (В) особи (аксиальные - главные сечения раковины): АА - кривая возрастания ширины последовательных камер у мегасферической особи; ВВ - то же у микросферической; по оси абсцисс - порядковые номера камер, по оси ординат - ширина камер. Пунктиром (I и II) показаны кривые соответствующих геометрических прогрессий со знаменателем q = 1,28 (из Rhumbler, 1923 по Schlumberger, 1883 и Rhumbler, 1911)
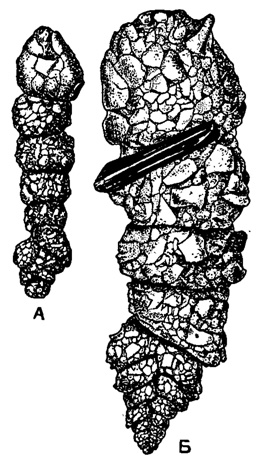
Рис. 8. Bigeneritia nodosaria Orbigny. А - мегасферическая особь; В - микросферическая особь. Обе раковины построены в основном из посторонних частиц различных размеров; обе гетероморфны (биморфны): начальный отдел каждой из них спирально-винтовой, двурядный, а поздний одноосный и однорядный (Rnumbler, 1911)
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'