
ГЕОХРОНОЛОГИЯ
ЭВОЛЮЦИЯ
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ПАЛЕОКЛИМАТОЛОГИЯ
ПАЛЕОЭКОЛОГИЯ
Отряд Endothyrida (Н. А. Волошинова, Е. А. Рейтлингер)
Раковина свободная, многокамерная, спирально-плоскостная или плектогироидная, иногда конечный отдел развернутый. Устье простое, ситовидное или ситовидное с добавочными устьями по септальным швам; стенка секреционная, известковая, зернистая, нередко с агглютинированными частицами (обычно известковыми), непористая или пористая, с простыми или сложными порами; у многих форм своеобразные дополнительные скелетные отложения. Девон - н. мел. Семейства: Endothyridae, Bradyinidae, Mezoendothyridae и условно Spirocyclinidae.
Первая эндотира была описана в 1843 г. из нижнего карбона Англии, там же позднее Брэди (Brady, 1876) был установлен ряд характерных видов. Однако, только изучение эндотирид в прозрачных шлифах Мёллером (1878, 1880) позволило выявить их основные морфологические признаки и показало их стратиграфическое значение для нижнего карбона. С тридцатых годов этого века в Советском Союзе начинается планомерное изучение эндотирид, подтвердившее их большую практическую ценность в разработке дробных стратиграфических схем Европейской части СССР. Только в последние годы (1950-1955) в Америке, при изучении эндотирид по шлифам, также было доказано их большое стратиграфическое значение для дробных зональных схем [работы Е. Целлер (Е. Zeller, 1950), Д. Целлер (D. Zeller, 1953), Скотта и др. (Scott and al., 1947)] в последнее время весьма интересны исследования Каммингса (Cummings, 1953) по пересмотру типов фораминифер коллекции Брэди и топотипов ряда родов.
Сем. Mezoendothyridae выделяется впервые. Два юрских рода из этого семейства - Choffatella Schlumberger, 1904 и Pseudocyclammina Jabe et Hanzawa, 1926 - были включены Кешмэном (1928) в сем. Lituolidae подсем. Lituolinae на основании их якобы песчаной стенки и "лабиринтовых" камер. Бонт (Bonte, 1942) отнес род Choffatella к сем. Spirocyclinidae Munier-Chalmas, 1887, выводя этот род как дегенерирующий из более древнего и более специализированного рода Spirocyclina. Два нижнемеловых рода - Stomatostoecha и Phenacophragma - были включены авторами этих родов (Applin, Loeblich et Tappan, 1950) в сем. Lituolidae подсем. Endothyrinae. Все эти мезозойские роды составляют самостоятельное семейство, которое включает вероятных потомков эндотирид.
Сем. Spirocyclinidae было выделено Мюнье-Шальма (Munier-Chalmas) в 1887 г., но незаслуженно забыто; так, относящиеся к нему роды были включены Кешмэном (Cushman, 1928) и Гэллоуэем (Galloway, 1933) в сем. Lituolidae, подсем. Lituolinae. Бонт (Bonte, 1942) в сем. Spirocyclinidae включил преимущественно высокоспециализированные роды с циклическими камерами на последней стадии развития.
Спираль представителей отр. Endothyrida плоская, или плектогироидная, или представляет собой Сочетание этих двух типов, иногда в конце роста развернутая. Плектогироидной называется спираль, расположенная в трехмерном пространстве и образованная путем последовательного отклонения оси навивания каждого последующего оборота или полуоборота на какой-то угол от положения оси предыдущего (рис. 150).
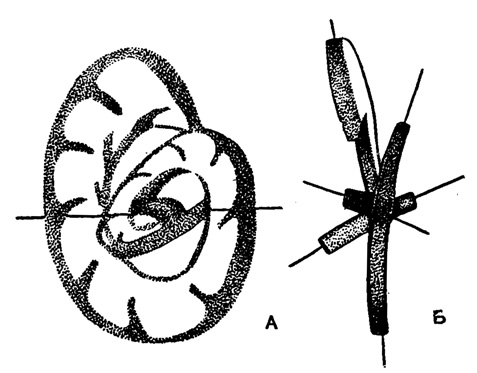
Рис. 150. А-Б. Схематическое изображение плектогироидной спирали (Scott H. W., E. Zeller and D. N. Zeller, 1947)
Септы в месте сочленения со стенкой образуют чаще тупой угол (наклонные септы) (рис. 151 А), реже угол около 90° (рис. 151 Б). В некоторых случаях (подсем. Chernyshinellinae) подразделение на камеры происходит за счет резкой выпуклости задней части камер, не сопровождающейся загибом стенки в месте причленения последующей камеры (рис. 151 В). Септы бывают плоские (рис. 151 В), выпуклые и крючковатые (рис. 151 Г).
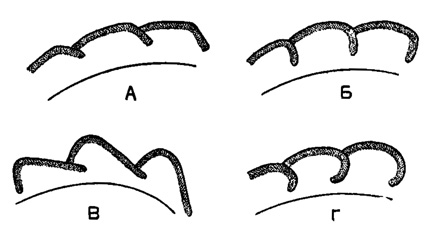
Рис. 151. А-Г. Типы септ Endothyrida, схемы: А - септы косые, отходящие под тупым углом от стенки; Б - септы прямые; В - чернышинелловый тип; Г - септы выпуклые и крючковатые
В стенке раковины часто дифференцируются слои, различной зернистости и окраски, иногда имеется дополнительный внутренний стекловатолучистый слой. Известны следующие типы стенки. 1) Стенка однослойная, неоднородно мелко- или тонкозернистая, с отдельными более крупными зернами (большинство турнейских плектогирин и чернышеллин). 2) Стенка однородно тонкозернистая, иногда с внутренним стекловато-лучистым слоем (большинство визейских плектогирин, эндотирин и Quasiendothyra). В некоторых случаях в стенке обособлен тонкий темный покровный слой (тектум) и имеются простые поры (табл. IV, фиг. 13 А, Б, группа Plectogyra omphalota). 3) Стенка многослойная, до четырех слоев: а) тонкий, темный покровный слой (тектум), б) светлосерый, неоднородно зернистый, в) темный, тонкозернистый, г) дополнительный стекловато-лучистый слой (табл. IV, фиг. 14, Globoendothyra). 4) Стенка грубозернистая, с агглютинированным материалом, преимущественно зернышки кальцита, редко мелкие раковины фораминифер, кварц и др., обычно дополнительный стекловато-лучистый слой (табл. V, фиг. 16, Endothyranopsis). 5) Стенка зернистая, с агглютинированным материалом (зерна кальцита, мелкие фораминиферы и мелкие обломки скелетов других организмов), с крупными порами, часто в виде разветвленных к поверхности пор-альвеол (табл. IV, фиг. 15-16, Bradyina). 6) Зернистая, двухслойная, с наружным непрободенным и внутренним ячеистым слоем (Pseudocyclammina, Cftoffatella).
Дополнительные скелетные образования нередко хорошо развиты у эндотирид, причем различные их типы свойственны определенным подсемействам и родам. Для подсем. Endothyrinae характерны отложения с двух сторон от устья характера хомат (рис. 152. А-В) и псевдохомат, а также утолщение септ над устьем. У подсем. Plectogyrinae дополнительные отложения располагаются параллельно септам в виде гребней и валиков по основанию камер, несколько отступя от устья. Подобный тип дополнительных отложений можно назвать экранным, так как он усложняет сообщение с внешней средой устья, отгораживая от основной камеры как бы вестибюль. Экраны бывают различных типов: в виде тонких гребней слабо наклоненных вперед или вогнутых, дающих в поперечном сечении крючки, а в продольном-бугорки (рис. 153. А-В; рис. 154. А-Б), в виде массивных валиков (рис. 154. В) и валиков, сливающихся в основании (рис. 154. Г). Довольно распространены у эндотирид дополнительные отложения, заполняющие боковые части камер и достигающие приустьевой области (Endothyranopsinae, рис. 155, и некоторые другие).

Рис. 152. А-В. Отр. Endothyrida. Дополнительные образования типа хомат: А - а продольном сечении оборота; Б - в поперечном качении оборота по линии е-е; В - в поперечном сечении оборота по линии f-f
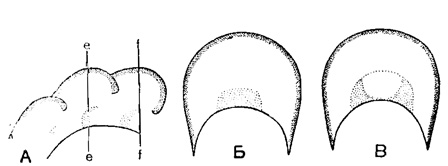
Рис. 153. А-В. отр. Endothyrida. Дополнительные отложения экранного типа - вогнутые гребни: А - поперечное сечение оборота; Б - продольное сечение оборота по линии е-е; В - продольное сечение оборота по линии f-f (Липина, 1955)
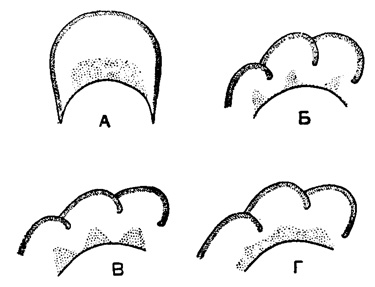
Рис. 154. А-Г. отр. Endothyrida. Экранный тип - валики: А - продольное сечение оборота; Б - поперечное сечение оборота; В - поперечное сечение оборота с массивными валиками; Г - поперечное сечение оборота со сливающимися валиками
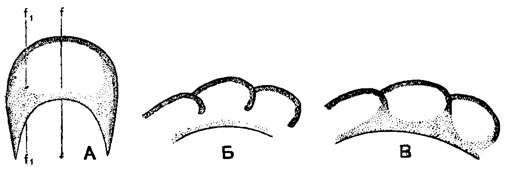
Рис. 155. А-В. Сем. Endothyridae. Дополнительные отложения в боковых частях камер, схемы: А - продольное сечение оборота; Б - поперечное сечение оборота по линии f-f; В - поперечное сечение оборота по линии f1-f1
Устье четырех типов: 1) простое - полулунное или щелевидное отверстие в основании камер (у большинства эндотирид); 2) простое овальное в центре устьевой поверхности (Paraendothyra); 3) ситовидное (Cribrospira, Michailovella и др.); 4) ситовидное устье и добавочные устья, расположенные вдоль септальных швов. Эти добавочные отверстия открываются в интерсептальные пространства, образованные септой и одной или двумя септальными пластинками и сообщаются с особой пупочной полостью (рис. 156-158).
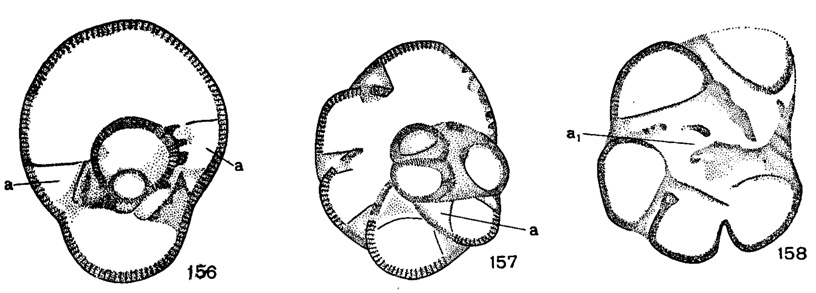
Рис. 156-158. Сем. Bradyinidae. Строение интерсептальных пространств, схемы: 156. Продольное сечение Bradyina: а - боковые камерки. 157. Поперечное сечение Bradyina: а - боковые камерки. 158. Поперечное сечение Janischewskina, а1 - боковая полость
Самые ранние находки сем. Endothyridae известны из ср. девона. Начиная с верхней части фаменского и турнейского ярусов, Endothyrida быстро развиваются (подсем. Endothyrinae, Chernyshinellinae, Plectogyrinae). Максимальный расцвет отряда приходится на визейский век, когда продолжают развиваться Plectogyrinae, Endothyrinae и появляются Endothyranopsinae и Bradyinidae. В последующее время (ср. и в. карбон, пермь) эндотириды эволюируют медленно, причем основное развитие получает сем. Bradyinidae. В триасе встречены фораминиферы, напоминающие эндотирид. С юры появляются сем. Mesoendothyridae и сем. Spirocyclinidae.
Эндотиридеи, по-видимому, в девонскую эпоху обособились от аммодисцид, вероятно от общего корня с турнейеллидами путем развития сначала пережимов, а потом все более четкой септации. В конце фаменского века и особенно в этреньское время развита своеобразная группа эндотирид - квазиэндотиры. Позднее развитие квазиэндотир пошло по двум основным направлениям: одна ветвь - в сторону усиления признака инволютности (род. Endothyra), другая - эволютности, уплощения формы и быстрого уменьшения роли дополнительных отложений (непостоянные у Loeblichia или только боковые уплотнения у Planoendothyra).
Начиная с турнейского века параллельно развиваются два крупных ствола: подсем. - Chernyshinellinae и Plectogyrinae. Первые характеризуются примитивным типом септации, вторые - своеобразным плектогироидным завиванием и часто дополнительными отложениями экранного типа. Plectogyrinae в визейский век развиваются в направлении увеличения углов отклонения оси навивания, увеличения общих размеров раковин, заменой дополнительных отложений типа тонких гребней массивными валиками, постепенно в процессе эволюции сливающимися и исчезающими. Строение стенки дифференцируется, приобретая в некоторых случаях четкую пористость, в других - многослойность. Появляются формы с развернутой спиралью и с ситовидным устьем (Michailovella). Начиная со средне каменноугольной эпохи развитие плектогирин значительно сокращается.
В визейский век обособляется подсем. Endothyranopsinae, развитие которого идет в сторону закрепления признаков спирально-плоскостного навивания, инволютности, увеличения размеров, усиления способности к агглютинированию.
В нижней части визейского яруса появляются первые представители сем. Bradyinidae, по всей вероятности, генетически связанные с подсем. Endothyranopsinae. Брэдииниды представляют резкое отклонение от общего ствола эндотиридей по признакам альвеолярного строения стенки и сложных добавочных устьев. В ср. карбоне развивается новый род Glyphostomella с тонкой стенкой, с септальными пластинами только в последнем обороте и небольшим числом камер.
Географическое распространение эндотиридей широкое. В карбоне одни и те же или близкие роды (иногда виды) найдены в Европе, Азии и Америке.
Как организмы, быстро реагирующие на изменение внешних условий, эндотиридеи имеют большое значение при дробных стратиграфических расчленениях, главным образом в течение времени своего наибольшего расцвета - в эпоху н. карбона.
[nom. transl. Rhumbler, 1895 (ex Endothyrinae H. В. Brady, 1884)]
Раковина от уплощенно-дисковидной до шарообразной, спирально-плоскостная или плектогироидная, иногда развернут. Число оборотов обычно 3-5; число камер в последнем обороте чаще не более 12. Устье простое, обычно полулунное, базальное, реже сложное. Стенки разнообразной зернистости, нередко с агглютинированными частицами, иногда из нескольких слоев и пористые. Обычны дополнительные отложения: хоматы, псевдохоматы, экраны и отложения, выстилающие основание и заполняющие боковые части камер. Девон - пермь; триас (?). Подсемейства: Endothyrinae, Chernyshinellinae, Plectogyrinae, Endothyranopsinae.
Раковина от уплощенно-дисковидной до наутилоидной, в ранней стадии часто плектогироидная, инволютная, во взрослой нередко спирально-плоскостная, эволютная. Устье простое, базальное, редко в срединной части устьевой поверхности или конечное ситовидное и с добавочными устьями. Стенка иногда пористая, иногда с стекловато-лучистым слоем. Дополнительные образования - хоматы, псевдохоматы и боковые заполнения. Семь родов. Девон - пермь.
Endothyra Phillips, 1846. Тип рода - Endothyra bowmani Phillips, 1846; н. карбон, Англия. Раковина наутилоидная, инволютная, спирально-плоскостная, иногда в ранней стадии слабо плектогироидная. Число оборотов 4-6, камер в последнем обороте 10-12, септы плоские, прямые. Устье базальное, обычно полулунной формы. Стенка однородно тонко- или мелкозернистая. Хоматы и псевдохоматы (табл. V, фиг. 1). Несколько видов. Н. карбон Русской платформы, Урала, Англии.
Quasiendothyra Rauser, 1948. Тип рода - Endothyra (?) kobeitusana Rauser, 1948; верхи девона (?), Куйбышевская обл. Раковина дисковидная или наутилоидная, инволютная и плектогироидная в ранней стадии, эволютная и спирально-плоскостная в поздней. Число оборотов обычно 4-5; камер от 10 до 17 в последнем обороте; септы выпуклые. Устье овальное или полулунное, базальное, реже ситовидное, из мелких отверстий. Стенка однородно тонкозернистая, нередко с внутренним стекловато-лучистым слоем. Дополнительные отложения в виде хомат и утолщений края септ над устьем (табл. V, фиг. 2,3). Около пяти видов. Верхняя часть девона - низы турне. Восточная часть Русской платформы, Урал, Новая Земля, Кавказ.
Planoendothyra Reitlinger gen. n. Тип рода - Endothyra aljutovica Reitlinger, 1950; cp. карбон, Рязанская обл. Отличается от Quasiendothyra иным характером дополнительных отложений: последние обычно заполняют боковые части камер, выстилают основание камер и образуют нередко псевдохоматы (табл. V, фиг. 4, 5). Несколько видов. Карбон Русской платформы, Урала, Кузбасса.
Loeblichia Cummings, 1955. Тип рода - Endothyra ammonoides Brady, 1873; н. карбон, Англия. Раковина маленькая, дисковидная, уплощенная, в ранней стадии плектогироидная и инволютная, в поздней (или, реже, вся раковина) спирально-плоскостная и эволютная. Число оборотов обычно 5-7, редко до 10; камер в последнем обороте чаще 13-20, иногда менее 13 или доходит до 27; септы выпуклые. Устье базальное, полулунной формы. Стенка тонкая, неоднородно тонкозернистая (рис. 159, табл. V, фиг. 6, 7). Псевдохоматы обычно развиты слабо и непостоянны. Около пяти видов. Визе юго-зап. части Русской платформы, Казахстана, Англии и Шотландии.
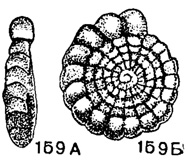
Рис. 159. А-Б. Сем. Endothyridae. Loeblichia translucens Dain; A - вид с периферии; Б - вид сбоку, × 72, карбон, визе, Донбасс (колл. Л. Г. Даин)
Paraendothyra N. Tchernysheva, 1940. Тип рода - Р. nalivkini N. Tchernysheva, 1940; турне, р. Сиказа на Ю. Урале. Раковина наутилоидная, не вполне инволютная, с глубокими пупками, почти спирально-плоскостная, на ранних стадиях иногда с небольшим отклонением оси навивания. Число оборотов 2-2,5, камер в последнем обороте 8-9. Устье полулунное, по середине септальной поверхности (рис. 160-162). Один вид. Турне Ю. Урала.
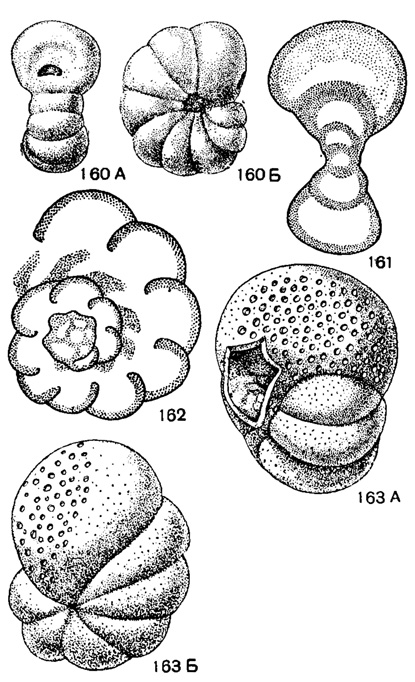
Рис. 160-163. Сем. Endothyridae. 160-162. Faraendolhyra nalivkini N. Tchernysheva; 160. А-Б - внешний вид, А - вид с периферии, Б - вид сбоку, × 30; 161 - продольное сечение; 162 - поперечное сечение, × 40, карбон, турне, Башкирская АССР (Н. Чернышева, 1940). 163. А-Б. Cribrospira panderi Moeller; A - вид со стороны устья; Б - вид сбоку, × 20, карбон, визейский ярус, Тульская обл. (Мёллер, 1880)
Cribrospira Moeller, 1878. Тип рода - С. panderi Moeller, 1878; визе, Тульская обл. Раковина почти шарообразная, инволютная, обычно почти спирально-плоскостная, с быстро возрастающими оборотами. Число оборотов не более двух с половиной, камер 5-7 в последнем обороте. Устье во взрослой стадии ситовидное. Стенка обычно тонкозернистая, тонкопористая (рис. 163, табл. V, фиг. 8). Три вида. Визе Европейской части СССР и Урала.
Janischewskina Mikhailov, 1935 (Samarina Rauser et Reitlinger, 1937). Тип рода - Janischewskina typica Mikhailov, 1935; визе, Ленинградская обл. Раковина почти шарообразная, состоящая из 2-3 оборотов, с 5-6 камерами в последнем обороте. Интерсептальные пространства в боковых частях раковины сообщаются с особой пупочной областью, в образовании ограничения которой участвуют септы и септальные пластинки (рис. 158). Устье - мелкие отверстия в основании выпуклых крышечек, прикрывающих многочисленные крупные округлые отверстия, расположенные на устьевой поверхности и вдоль швов (рис. 164-166). Мало видов. Визе Европейской части СССР.
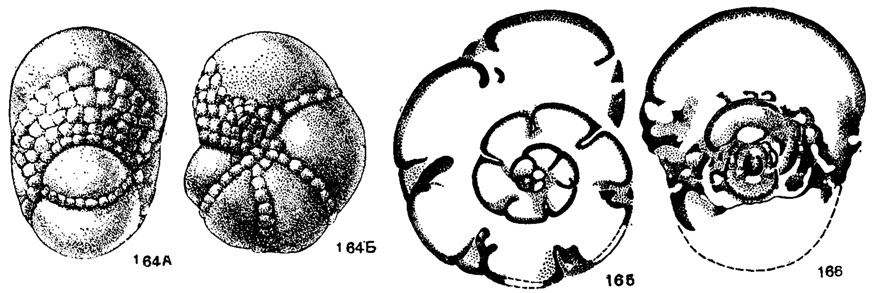
Рис. 164-166. Сем. Endothyridae. Janischewskina typica Mikhailov: 164. A - вид со стороны устья; 164. Б - вид сбоку, × 14; 165 - поперечное сечение; 166 - продольное сечение, × 25, карбон, визе, Куйбышевская обл. (Раузер-Черноусова и Рейтлингер, 1940)
Раковина обычно почти шарообразная, инволютная или частично эволютная, иногда выпрямляющаяся, в спиральной части плектогироидная. Подразделение на камеры происходит путем вздутия задней части камер и причленения последующей камеры почти у края предыдущей, без загиба стенки в месте сочленения; в ранней стадии иногда только пережимы. Устье простое. Стенка неоднородно мелкозернистая. Редко дополнительные отложения в виде валиков позади устья. Два рода. Турне.
Chernyshinella Lipina, 1955. Тип рода - Endothyra glomiformis Lipina, 1948; турне, Калужская обл. Раковина состоит из трех-четырех, реже пяти, оборотов; камер от трех до семи в последнем обороте. Устье образовано суженной передней частью камер. Редко дополнительные отложения в виде валиков (рис. 167, табл. V, фиг. 9). Около пяти видов. Турне Русской платформы, Урала и Кузбасса; возможно турне (свита киндергук) С. Америки.
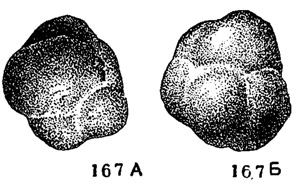
Рис. 167. А-Б. Сем. Endothyridae. Cherny shinella glomiformis (Lipina); A - вид со стороны устья; Б - вид сбоку, × 72, карбон, турнейский ярус, Донбасс (колл. Л. Г. Даин)
Chernyshinellina Reitlinger, gen. nov. Тип рода - Ammobaculites. pygmeus Malachova, 1954; турне, р. Чусовая (Урал). Отличается от Chernyshinella наличием выпрямленной цилиндрической части с однорядным расположением камер. Устье простое, базальное в спиральной части и конечное в развернутой (табл. V, фиг. 10). Один вид. Турне Ср. Урала.
Раковина дисковидная или наутилоидная, иногда выпрямляющаяся, инволютная или полуэволютная, завитая по плектогироидной спирали. Стенка зернистая, нередко состоит из нескольких слоев, иногда ясно пористая. Устье простое, редко ситовидное. Обычны дополнительные отложения экранного типа или сплошь выстилающие основание камер. Пять родов. Карбон и пермь.
Plectogyra Е. Zeller, 1950. Тип рода - Р. plectogyra Е. Zeller, 1950; миссисипская система, (визе), С. Америка. Раковина дисковидная или наутилоидная, инволютная, частично эволютная. Число оборотов около трех, камер 6-10 в последнем обороте. Устье базальное, полулунное. Стенка обычно однородно зернистая, иногда с наружным тонким микрозернистым тектумом; нередко видимы тонкие прямые поры. У большинства форм имеются дополнительные отложения в виде гребней, валиков, расположенных позади устья, или выстилающие основание камер (табл. V, фиг. 11). Около 25 видов. Карбон-пермь. Русская платформа, Урал, Кузбасс, Англия и С. Америка.
Pledogyrina Reitlinger, gen. n. Тип рода - Endothyra (?) fomichaensis Lebedeva, 1954; верхи турне - низы визе, Кузбасс. Отличается от Pledogyra характером навивания раковинки - внутренние обороты почти спирально-плоскостные, эволютные, наружный оборот расположен под углом, близким к 90° по отношению к предыдущим, и инволютен (табл. V, фиг. 12). Около пяти видов. Верхи турне - низы визе Кузбасса; возможно в. турне вост. части Русской платформы; свита киндергук С. Америки.
Globoendothyra, Reitlinger, gen. n. Тип рода - G. pseudoglobulus Reitlinger nom. nov. (= Endothyra globulus Moeller, 1878, non Eichwald); визе, Тульская обл. Раковина наутилоидная, инволютная, нередко во взрослой стадии почти спирально-плоскостная. Число оборотов 3-5; в последнем обороте обычно 9-10 камер. Септы наклонные. Устье базальное, полулунное. Стенка неоднородно зернистая, обычно многослойная, часто с внутренним стекловато-лучистым слоем. Дополнительные отложения обычно выстилают основание камер, редко в последней камере гребень позади устья (рис. 168, табл. V, фиг. 13). Около десяти видов. Визе, редко турне. Европейская часть СССР и Кузбасс.
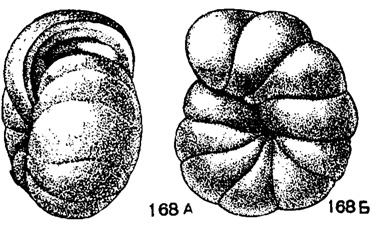
Рис. 168. А-Б. Сем. Endothyridae. Globoendothyra pseudoglobulus Reitlinger; A - вид со стороны устья; В - вид сбоку, × 22, карбон, визе, Тульская обл. (Мёллер, 1878)
Mikhailovella Ganelina, 1956. Тип рода - Endothyrina (?) gradlis Rauser, 1948; визе, Тульская обл. Раковина на ранней стадии плектогироидная, инволютная, позднее однорядная, субцилиндрическая. Устье простое на ранней стадии роста, в последних камерах спиральной части и во всей однорядной ситовидное. Стенка обычно тонкозернистая (табл. V, фиг. 14). Мало видов. Визе Европейской части СССР.
Endothyranella Gallowayet Harlton, 1930. Тип рода - Ammobaculitespowersi Harlton; 1927; ср. карбон, Оклахома. Раковина на ранней стадии инволютная или полуэволютная, в поздней выпрямляется. Число оборотов около трех, камер 5-10 в обороте. Устье базальное, полулунное в спиральной части и конечное, округлое или овальное в прямой. Стенка обычно тонкозернистая (табл. V, фиг. 15). Около семи видов. Ср. и в. карбон Европейской части СССР; верхняя часть миссисипской и пенсильванской систем С. Америки.
Раковина от дисковидной до шарообразной, почти спирально-плоскостная, обычно инволютная. Устье простое базальное. Стенка с явно агглютинированным материалом, иногда со стекловато-лучистым слоем. Псевдохоматы и заполнение боковых частей камер. Два рода, один из них Rhenothyra - условно. Девон (?), н. карбон.
Endothyranopsis Cummings, 1955. Тип рода - Involutina crassa Brady, 1869; н. карбон, Англия. Раковина состоит из 3-4 оборотов; камер 10-13 в последнем обороте, септы плоские, прямые. Устье полулунное. Стенка толстая, неоднородно зернистая с агглютинированными частицами, часто с внутренним стекловатолучистым слоем. Псевдохоматы и заполнение боковых частей камер (рис. 169, табл. V, фиг. 16-17), около пяти видов. Визе (возможно, турне). Русская платформа, Урал, Кузбасс, З. Европа, С. Америка (?).
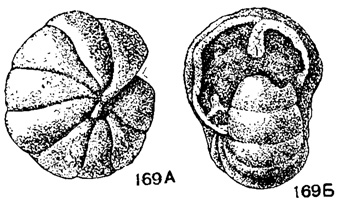
Рис. 169 А-Б. Сем. Endothyridae. Endothyranopsis crassus (Brady); A - вид сбоку; Б - вид со стороны устья. × 30, карбон, визе, Англия (Brady, 1876)
Rhenothyra Beckman, 1950. Тип рода - Rh. refrathensis Beckmann, 1950; ср. девон, окрестности Кёльна (Германия). Раковина дисковидная, эволютная, спирально-плоскостная. Число оборотов около трех; камер в последнем обороте до пятнадцати. Перегородки в ранней стадии перпендикулярны к стенке, в поздней слегка наклонены вперед. Устье округлое. Стенка толстая (по всем оборотам равной толщины), тонкозернистая, со включением очень мелких агглютинированных частиц, трехслойная (рис. 170-172). Один вид. Ср. девон Германии.
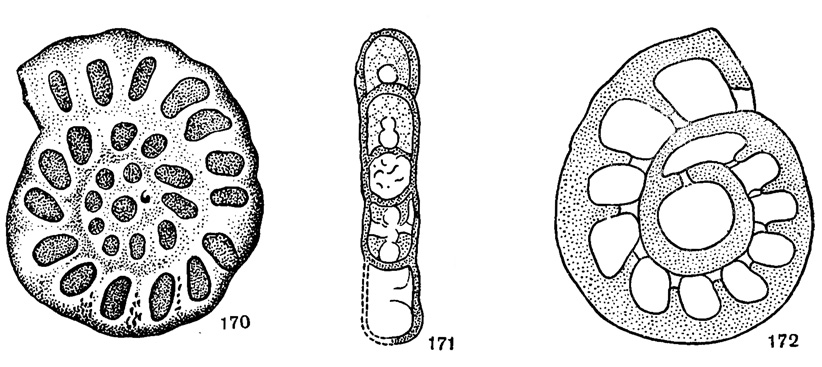
Рис. 170-172. Сем. Endothyridae; Rhenothyra refrathensis Beckmann; 170 - типичный представитель; 171 - сагиттальное сечение; 172 - поперечное сечение (Beckmann, 1950)
Раковина от уплощенно-наутилоидной до шарообразной, почти спирально-плоскостная, инволютная. Камеры часто разделены интерсептальными пространствами. Последние образованы септой и двумя тонкими пластинами, постсептальной и пресептальной, являющимися выростами стенки непосредственно позади и спереди септы. Устье обычно сложное, по крайней мере во взрослой стадии; отверстия на септальной поверхности и добавочные устья вдоль швов, открывающиеся в интерсептальные пространства. Стенка пористая, обычно с агглютинированными частицами (преимущественно зернышками кальцита, реже мелкие раковины фораминифер и детрит скелетов других организмов); у внешней поверхности обособляется очень тонкий более темный слой типа тектума. Три рода. Карбон, пермь.
Bradyina Moeller, 1878. Тип рода - Nonionina rotula Eichwald, 1860; визе, Тульская обл. Раковина наутилоидная, оборотов не более трех с половиной, камер в последнем обороте 5-9. Обе септальные пластины развиты с самой ранней стадии роста; пластины каждой камеры в боковой части раковины срастаются с задней частью камер и образуют боковые камерки сообщающиеся с интерсептальными пространствами (рис. 157). Устья - круглые или щелевидные отверстия, расположенные по всей септальной поверхности и вдоль швов. Стенка толстая, часто с разветвленными порами (рис. 173-175, табл. V фиг. 18). Около 20 видов. Карбон (с визе), реже - пермь. Русская платформа, Урал, Англия, Монголия, Китай.
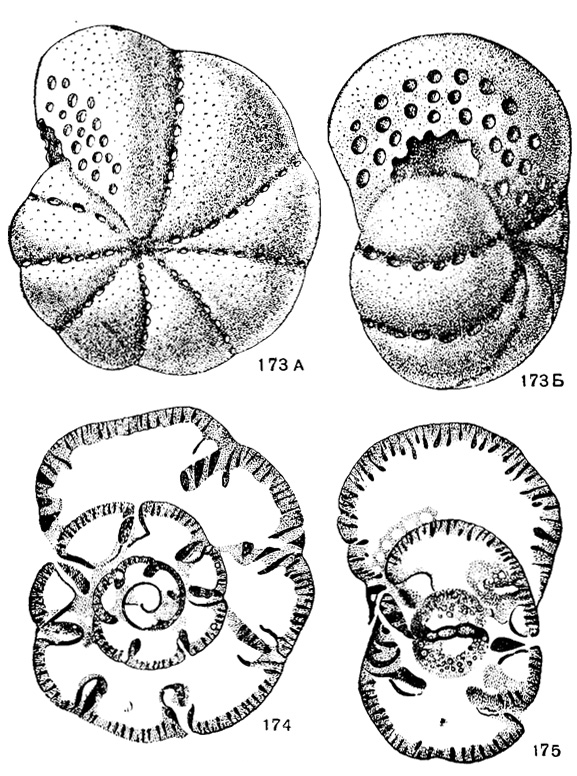
Рис. 173-175. Сем. Bradyinidae. 173-175. Bradyina rotula (Eichwald); 173. А - вид сбоку; Б - вид со стороны устья, × 10; 174. Поперечное сечение; 175. Продольное сечение. × 20, карбон, визе, Куйбышевская обл. (Раузер-Черноусова и Рейтлингер, 1910)
Glyphostomella Cushman et Waters, 1928. Тип рода - Ammochilistoma (?) triloculina Ciishman et Waters, 1927; cp. карбон, Техас. Отличается от Bradyina малым числом камер по оборотам (в последнем обороте обычно три-четыре), обычно более сжатой или более шарообразной раковиной, относительно тонкой стенкой (особенно во внутренних оборотах) с более тонкими, обычно простыми порами, присутствием септальных пластин только в камерах последнего оборота, характером устья; у типа рода С. Америки устье на ранней стадии представлено щелевидными отверстиями, расположенными параллельно шву, а во взрослой стадии - под прямым углом к последнему (табл. V, фиг. 19). Около семи видов. Ср. и в. карбон, пермь. Русская платформа, Урал, Ср. Азия, С. Америка.
Pseudobradyina Reitlinger, 1950. Тип рода - Р. pulchra Reitlinger, 1950; ср. карбон, Архангельская обл. Раковина маленькая, число оборотов около двух; обычно три камеры в последнем обороте. Септальные пластины часто не развиты или имеется только постсептальная пластина в последней или предпоследней камере. Устье - щелевидные отверстия в последней камере перпендикулярно основанию септальной поверхности. Стенка тонкая, мелкозернистая и тонкопористая (рис. 176, табл. V, фиг. 20). Мало видов. Ср. карбон (мячковский горизонт) и в. карбон Русской платформы.
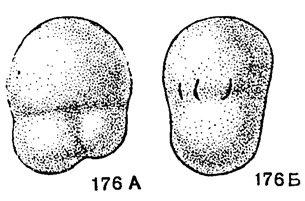
Рис. 176. Сем. Bradyinidae. А-Б Pseudobradyina pulchra Reitlinger; А - вид сбоку; Б - вид со стороны устья, × 40, карбон, касимовский ярус, Московская область (Рейтлингер, 1950)
Раковина на ранних стадиях большей частью спирально-плоскостная, инволютная, редко плектогироидная, на поздних иногда развертывается с образованием однорядного отдела. Устье или щелевидное - внутри - краевое, или септальное, или сентальное ситовидное. Стенка известковая, иногда с агглютинированным материалом во внешнем слое, однослойная непрободенная или двуслойная с наружным непрободенным и внутренним грубопрободенным ячеистым слоем; септы однослойные непрободенные или прободенные. Около пяти родов. Юра, н. мел.
Mesoendothyra Dain gen. n. Тип рода - М. izjumiana Dain, 1956; в. юра, Украина. Раковина инволютная, на самых ранних стадиях плектогироидная. Устье щелевидное, внутри краевое. Стенка, за исключением септ, двуслойная; септы однослойные, непрободенные (рис. 177-178). Один вид. Кимеридже Украины.

Рис. 177-189. Сем. Mesoendothyridae. 177-178. Mesoendothyra isjumiana Dain; 177 - поперечное сечение, 178 - продольное (аксиальное) сечение, × 50, н. кимеридж, Украина (колл. Л. Г. Дайн). 179-181. Pseudocyclammina lituus (Yokoyama); 179 - вид сбоку, распрямленный отдел реконструирован, 180 - устьевая сторона, 181 - поперечное сечение, × 40, юра - мел (переходные слои), Япония (по Yokoyama Galloway, 1933). 182-183. Pseudocyclammina ukrainica Dain; 182 - A-B внешний вид: А, Б - с боков, В - с периферического края, со стороны септальной поверхности, × 30, 183 - медианное сечение, × 50, кимеридж, Украина (колл. Л. Г. Дайн). 184-185. Choffaiella decipiens Schlumberger; 184 А-Б - внешний вид; А - сбоку, Б - с периферического края, × 12, альб, Португалия (Schlumberger, 1904), 185 - поперечное сечение, × 14, баррем, Швейцария (Tobler, 1928). 186-187. Stomatostoecha plummerae Applin, Loeblich et Tappan; 186 А-Б - внешний вид: A - сбоку, Б - с периферического края, × 26, 187 - медианное сечение, × 30, н. мел, Техас (Applin, Loeblich and Tappan, 1950. 188-189. Phenacophragma assurgens Applin, Loeblich et Tappan; 188 A-Б; внешний вид: A - сбоку, Б - с периферического края, × 50, 189 - медианное сечение, × 64, н. мел, Техас (Applin, Loeblich and Tappan, 1950)
Pseudocyclammina Yabe et Hanzawa, 1926. Тип рода - Cyclammina lituus Yokoyama, 1890; юра - мел (переходные слои), Япония. Раковина на ранних стадиях спирально-плоскостная, инволютная, с 5-15 камерами в последнем обороте, на поздних или слегка развертывается, или образует отчетливый однорядный отдел. Устье ситовидное. Стенка, за исключением септ, двухслойная; септы однослойные, грубопористые (рис. 179-183). Несколько видов. В. юра Европы (в том числе Европейской части СССР); переходные слои от юры к мелу Японии.
Choffatella Schlumberger, 1904. Тип рода - Ch. decipiens Schlumberger, 1904; гольт (альб), Португалия. Раковина на ранних стадиях инволютная, на поздних иногда (редко) с тенденцией к развертыванию и образованию однорядного отдела; сильно уплощенная, с 15-25 короткими изогнутыми, быстро увеличивающимися в ширину камерами в последнем обороте. Устье - вертикальный ряд отверстий на устьевой поверхности. Стенка иногда с большим или меньшим количеством агглютинированного материала, по структуре двухслойная; поры внутреннего слоя иногда раздваиваются и анастомозируют у наружного слоя; септы однослойные, прободенные (рис. 184-185). 5-6 видов. Юра (оксфорд) - н. мел (альб.). Ю. Европа, ю.-з. Азия (Ближний Восток), С. Африка, Япония, Центр. и Ю. Америка.
Вне СССР, встречаются: Stomatostoecha Applin, Loeblich et Tappan, 1949 (рис. 186-187); Phenacophragma Applin, Loeblich et Tappan, 1949 (рис. 188-189).
Раковина, в типичном случае, проходит три стадии развития: начальную - спирально-плоскостную, промежуточную - с почкообразно изогнутыми камерами, и конечную - циклическую; камеры очень многочисленны, большей частью подразделены перегородками на множество вторичных камерок. Устье или в виде многочисленных отверстий на периферическом крае, или ситовидное на устьевой поверхности. Стенка непрободенная, иногда ячеистого строения. Пять родов. Юра (лейас) - эоцен.
Orbitopsella Munier-Chalmas, 1902. Тип рода - Orbitolites praecursor Giimbel, 1872; лейас, Италия. Первая и вторая стадии короткие, быстро сменяются циклической; периферические камеры по сравнению с центральными значительно утолщены; подразделены на правильные прямоугольные камерки. Устье на периферическом крае (рис. 190-191). 2-3 вида. Юра (лейас), Европа.

Рис. 190-202. Сем. Spirocyclinidae. 190-191. Orbitopsella praecttrsor (Giimbel); 190 - внешний вид; 191 - аксиальное сечение; × 8; юра, Италия (Cumbel из Cushman, 1948). 192-194. Orbitammina elliptica (Arehiac); 192 - внешний вид, в нат. велич.; 193 - схема расположения основных камер (без начальной части), × 2,5; 194 - фрагмент поперечного (экваториального) сечения, показывающий деление основных камер на камерки, × 15, батский ярус, Франция, Арденны (Bonte, 1942). 195. Spirocyclina choffati Munier-Chalmas; внешний вид, × 3, в. юра (портланд), Португалия (Schlumberger, 1904). 196-197. Spirocyclina sp. 196 - схема спирально-плоскостного расположения начальных камер; 197 - схема расположения почкообразно изогнутых камер и перехода к циклическим (Charles, 1932). 198. Spirocyclina choffati var. euxina Charles; поперечное (аксиальное) сечение перегородки камер; видно своеобразное альвеолярное строение стенок с внутренними столбиками, × 30 (Charles, 1932). 199. Diciyopsella chalmasi Schlumberger; внешний вид - сбоку, × 10, мел, Испания (Vaughan из Gacloway, 1933), 200. Diciyopsella kiliani Munier-Chalmas; поперечное (экваториальное) сечение, × 15, сантон, Испания (Schlumberger, 1900). 201-202. Yaberinella jamaicensis Vaughan; 201 - внешний вид, × 4, эоцен, Ямайка (Vaughan из Galloway, 1933); 202 - фрагмент сечения, показывающий внутренние перегородки камер, × 15 (Vaugnan из Galloway, 1933)
Orbitammina Berthelin, 1893. Тип рода - Orbicula elliptica Archiac, 1843; бат, Франция. Начальная стадия неизвестна, вероятно, спирально-плоскостная; поздняя характеризуется почкообразно изогнутыми камерами; основные камеры подразделены перегородками на вторичные камерки. сообщающиеся между собой посредством канальцев. Устье - на периферическом крае (рис. 192-194). Один вид. Ср. юра (бат и байос) Европы.
Spirocyclina Munier-Chalmas, 1887. Тип рода - S. choffati Munier-Chalmas, 1887; н. мел, Португалия. Спирально-плоскостные камеры сменяются почкообразно изогнутыми, которые, в свою очередь, в некоторых случаях (не всегда) сменяются циклическими; камеры очень неравномерны подразделены перегородками на вторичные камерки. Устье на периферическом крае. Стенка своеобразного ячеистого строения с короткими внутренними столбиками (рис. 195-198). Два-три вида. В. юра Крыма; в. юра и н. мел Европы.
Dictyopsella Munier-Chalmas, 1899. Тип рода - D. kiliani Munier-Chalmas, 1899; сенон. Португалия. Раковина спирально-плоскостная, сильно уплощенная; циклическая стадия слабо выражена. Камеры ясные, отчетливо подразделены радиальными и вертикальными перегородками, образующими подобие сетки, на мелкие камерки. Устье ситовидное (рис. 199-200). Один вид. В. мел Европы.
Вне СССР: Yaberinelta Vaughan, 1928 (рис. 201-202).
|
ПОИСК:
|

При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paleontologylib.ru/ 'Палеонтология - книги и статьи'